
Abstract
This study integrates bone collagen stable isotope data (carbon, nitrogen and sulphur) from 33 human adult tibiae (15 females; 18 males) and 13 faunal remains from Tomar, while it was under the Military Orders domain (eleventh–seventeenth centuries). Historical literature indicates that the amount of meat consumption amongst Templars was lower than in individuals with similar social status. In Medieval times, these Military Orders had total control of towns and angling and fishing rights, but their influence on the general population diet remains unknown. While no statistically significant differences (p > 0.05) were found between sexes, social status, or for bone collagen δ13C and δ34Sbetween age groups, δ15N did differ significantly with age, which may be related to tooth loss in old individuals. Additionally, the human samples have higher stable isotope differences, in comparison with faunal samples, than would be expected within the food web, particularly for δ13C. This human bone collagen δ13C enrichment may reflect a diet rich in aquatic protein intake, which is also supported by δ34S archived in human and faunal samples, and the presence of oysters and cockles shells at the excavation. The religious diet restrictions might have led to a higher intake of aquatic protein when meat consumption was not allowed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
This study investigates diet in a town ruled by religious military orders and how the general population diet may have adapted to religious dietary restrictions. This study is the first of its kind to analyse carbon, nitrogen and sulphur stable isotope data from skeletons of the eleventh–seventeenth centuries in Portugal.
Stable isotopic analysis
Analysis of stable isotope ratios from mineralised tissue has been widely used for dietary reconstruction. This technique is based on the assumption that “you are what you eat (plus a few ‰)” (DeNiro and Epstein 1976), as a consumer’s tissues reflect the isotopic array of the ingested foods. Food webs have an impact on carbon isotope values due to the correlation between animal tissue carbon values (δ13C) and their diet (DeNiro and Epstein 1978; Teeri and Schoeller 1979). There is enrichment in δ13C in an animal’s body tissues relative to its diet due to the fractionation that occurs during the formation of tissues (van der Merwe and Vogel 1978). Primary consumers have a fractionation factor (enrichment in δ13C) of approximately 5‰ in their bone collagen relative to their diet (van der Merwe and Vogel 1978; Ambrose and Norr 1993) and an enrichment of 1‰ between trophic levels (DeNiro and Epstein 1978; Tieszen et al. 1983). In marine plants, the main carbon source is dissolved carbonate (0‰), instead of atmospheric CO2 (− 7‰), therefore, this difference is reflected in the δ13C in tissues of mammals feeding from these two different ecosystems (Tauber 1981; Chisholm et al. 1982, 1983). δ13C in bone collagen can also help in identifying freshwater resources. Katzenberg and Weber (1999) observed a range of − 14.2 to − 24.6‰ in fish bones in Siberia with higher δ13C in species inhabiting shallow waters and lower δ13C for fish inhabiting deeper open waters on the lake. Freshwater fish exhibit variation in δ13C depending on the ecosystem as freshwater plants have numerous sources of carbon, unlike terrestrial plants (Zohary et al. 1994; Dufour et al. 1999).
In terrestrial ecosystems, there is an increment of 3 to 5‰ in δ15N between trophic levels when compared with consumer diet (Schoeninger et al. 1983; Minagawa and Wada 1984; Schoeninger and DeNiro 1984; Bocherens and Drucker 2003). This fractionation enables the use of stable nitrogen isotopes (δ15N) to infer trophic level, and high δ15N recorded in bone collagen usually indicates high-protein diets (Sponheimer et al. 2003). Nitrogen isotope values can also be used to differentiate between terrestrial and marine food sources (DeNiro and Epstein 1981; Schoeninger et al. 1983; Walker and DeNiro 1986; Richards and Hedges 1999), especially when combined with carbon isotope data. Bone collagen δ15N can also be used to analyse access to freshwater resources, as organisms in these ecosystems exhibit higher δ15N than those in terrestrial ecosystems (van Klinken et al. 2002).
Advances in mass spectrometry and methodology development, following the work of Leach et al. (1996) allow an easier and more frequent analysis of sulphur isotope data (δ34S). Sulphur isotope analysis can shed some light on the use of freshwater or marine resources (Nehlich and Richards 2009; Nehlich et al. 2010; Nehlich 2015), especially when combined with the analysis of carbon and nitrogen-stable isotopes. A freshwater ecosystem, which is highly dependent on the geological conditions and source of water sulphates (Nehlich 2015), has an impact on terrestrial δ34S, especially if the fauna fed on the floodplains of the river (Fry 2002; Nehlich et al. 2011). δ34S at riverine ecosystems fall between − 5 and + 15‰, but the values can be outside this range in a relatively small geographical scale due to specific environmental conditions (Nehlich 2015).
Historic background
The city of Tomar had a very important military role consolidating the Kingdom of Portugal by resisting the advances of the last Moroccan king of Hispania, Iacub ben Iuçuf Almançor (França 1994). The construction of the Convent of Christ, a Templar stronghold, began in 1160 and was also likely around that time that the Church of Santa Maria do Olival was constructed (Conde 1996). In 1317, Pope Clement issued the Papal Bull Pastoralis Praeeminentiae, which instructed all Christian monarchs in Europe to arrest all Templars and seize their assets (Barber 2012). Portugal successfully lobbied the papacy, and the Templars did not face a trial, instead the Order’s assets and personnel were transferred to the newly established Order of Christ, a continuation of the Templars in Portugal (Valente 1998). Tomar then became a centre of Portuguese overseas expansion under Henry the Navigator, the Grand Master of the Order of Christ (Conde 1996).
Trade in Europe began to increase in the eleventh century (Malgosa 2011), since Tomar was located at the main Portuguese road connecting the North of the country to the limits of the Reconquista (Conde 1996). Given Tomar’s location, it would have frequent movement of goods but also people and one of its functions was to receive and protect refugees in case of invasion (Conde 1996).
According to historical data, the staple Medieval diet in Portugal was bread accompanied by wine, olives and olive oil (Vicente 2013). A significant part of agriculture was focused on cereals, but a large percentage of the harvest was inaccessible to peasants after paying tributes to lords and the church (Vicente 2013). Chestnuts and sweet acorns could sometimes substitute the bread (Vicente 2013), and some legumes could be reduced to flour when there was a lack of cereals (Gonçalves 2004). The acorns were frequently used to feed the livestock, especially swine that also fed from various roots and mushrooms (Vicente 2013).
Cattle were not abundant, compared with sheep and goats, and only the pigs were purposely raised for meat production (Gonçalves 2004). Other sources of meat were chicken, duck and goose as well as a variety of game (Gonçalves 2004). For the peasants, hunting could represent the only access to meat; however, in Tomar, angling and warren rights were reserved for the military orders. Amongst Medieval Iberian faunal assemblages the domestic animals predominate (Grau-Sologestoa 2017), which can be a result of hunt restrictions. Fish was an expensive food, with the exception of sardines which were more abundant and easy to preserve salted or smoked (Gonçalves 2004). Fish was indispensable during the numerous fast days that the Medieval religious calendar imposed (Vicente 2013) but it was consumed more in the littoral despite the availability of Portuguese rivers (Gonçalves 2004). Molluscs and crustaceans were also part of the diet of all social status but were considered a “food of the poor” due to their abundance (Gonçalves 2004).
Various studies suggest dietary differences between sex, age groups and social status in Medieval times (e.g. Adamson 2004, Kjellström et al. 2009, Linderholm et al. 2008, Polet and Katzenberg 2003, Schutkowski et al. 1999, Reitsema et al. 2010, Reitsema and Vercellotti 2012). Since fish was expensive but necessary for religious fasts and the military orders had angling and warren rights, diet in Tomar may also reflect social status. The historical literature (Barber and Bate 2002) implies that the amount of meat consumption amongst Templars was lower than in individuals with similar social status, and the intake of vegetables was higher. In Tomar, merchants, crafters and farmers participated actively at the local army alongside with knights, raising their status (Conde 1996) and probably having access to similar food resources to the Templars.
Materials and sampling
This study analyses bone collagen stable isotope data from 33 human adult tibiae (15 females; 18 males) and 13 fauna remains (2 wild Sus; 2 domestic Sus; 1 juvenile Sus; 1 Canidae; 3 Bos; 1 Equus; 3 Ovicapridae) from Santa Maria do Olival graveyard (eleventh–seventeenth centuries), in Tomar, Portugal. Only individuals from areas 13 to 20 (2nd phase of the excavation; Annex, Fig. A1) were analysed. Areas 13, 15, 18 and 19 were considered to be a place of burial for individuals with higher social status not only due to the proximity to the church (Binski 1996; Daniell 1998; Graves 1989; Ottaway 1992; Platt 1981; Swanson 1989) but also because of the higher frequency of structured graves. Faunal remains were collected from areas 14,17 and 20 (Annex, Fig. A1). The faunal remains from area 20 were mixed with human remains in an ossuary with at least 14 human adults. The faunal remains recovered from areas 14 and 17 were in grave fill material.
The skeletons (all ages and both sexes) distribution within the necropolis suggest that Santa Maria do Olival collection represents the general population of Tomar and not, or at least not only, the individuals from the military orders. The uniform spatial distribution between sexes within the graveyard and the use of structured graves for both males and females suggest that, at least at death, social status was not dependent on sex. However, social status seems to increase with age as older individuals were more frequently buried in structured graves.
Only individuals without signs of physiological stress were sampled in an attempt to estimate the diet of the general population and avoid isotopic data that may represent differing metabolism during disease and/or malnutrition (Steele and Daniel 1978; Hobson and Clark 1992; Hobson et al. 1993; Gaye-Siessegger et al. 2004; Fuller et al. 2005; D'Ortenzio et al. 2015). To avoid sampling individuals with physiological stress, only individuals without skeletal markers of stress, such as cribra orbitalia or obvious enamel hypoplasias, were selected. Since low stature can also be associated with physiological stress (e.g. Haviland 1967; Morris and McAlpin 1979; Allen and Uauy 1994; Roberts and Manchester 2007; Moore and Ross 2013), only individuals with maximum length of the skeleton (measurement was taken during the excavation while the skeleton was still articulated, in situ, in extended supine position and used as a proxy for stature) equal or above the mean for this population (151.8 ± 6.1 cm for the females, n = 256; 163.4 ± 7.5 cm for the males, n = 287) were sampled.
Methods
Sex was estimated based on pelvic (Phenice 1969; Buikstra and Ubelaker 1994) and cranial features (Buikstra and Ubelaker 1994). Adult age at death estimates employed a combination of skeleton maturation (Scheuer and Black 2000), pubic symphysis degeneration (Brooks and Suchey 1990; Buikstra and Ubelaker 1994) and auricular surface degeneration (Lovejoy et al. 1985). The skeletons analysed were classified as young (18 to 29 years), mature (30 to 60 years) and old (more than 60 years) adults.
Collagen extraction was done following Longin (1971), Brown et al. (1988) and Richards and Hedges (1999). The collagen samples were weighed into tin capsules and combusted into CO2 and N2 using an elemental analyser (Flash/EA) coupled to a Thermo Finnigan DeltaPlus XL isotope ratio mass spectrometer via a ConFlo III interface at NERC Isotope Geosciences Facility (Nottingham, UK). Sulphur stable isotopes were analysed at the Faculdade de Ciências da Universidade de Lisboa (Lisbon, Portugal). δ13C and δ15N were calibrated using an in-house reference material M1360p (powdered gelatine from British Drug Houses) with expected δ values of − 20.32‰ (calibrated against CH7, IAEA) and + 8.12‰ (calibrated against N-1 and N-2, IAEA) for carbon and nitrogen, respectively. Samples were run in duplicate and the 1σ reproducibility for mass spectrometry controls for these analyses were δ15N = ± 0.08‰ and δ13C = ± 0.07‰. The sulphur isotope analysis was done at SIIAF (University of Lisbon), using an IsoPrime mass spectrometer. The collagen was combusted with additional V2O5 and a pulse of oxygen. δ34S was calibrated using the inorganic international standards NBS127 (+ 20.3‰), IAEA S1 (− 0.3‰) and casein protein (+ 4.0‰). Mass spectrometry control for these analyses was δ34S = ± 0.08‰.
Mann-Whitney U non-parametric tests were used for pair-wise comparisons, and Kruskal-Wallis non-parametric tests were used to compare more than two groups. All statistics were computed in SPSS 24 for Windows and p ≤ 0.05 were considered statistically significant.
Results
The bones from all individuals in the present study had acceptable C:N ratios (2.9 to 3.6; DeNiro 1985) and S% (0.15 to 0.35%, Nehlich and Richards 2009). Herbivores, with the exception of Equus, have similar values for bone collagen δ13C (− 21.2 to − 20.9‰), while bone collagen δ15N (4.8 to 7.8‰) and δ34S (13.1 to 18.5‰) values are more variable (Figs. 1 and 2). The domestic Sus and the only carnivore analysed (Canidae) have similar δ13C and δ15N values to the herbivores (Fig. 1). The faunal remains have higher bone collagen δ34S than the human remains (Fig. 2), only the Equus displays bone collagen δ34S expected for an exclusive terrestrial diet.

Stable isotope values of fauna and human (from different social status) bone collagen. Lines indicate the mean without the outlier (δ13C = − 18.6‰; δ15N = 10.8‰) and two standard deviations (μ ± 2σ). Grey area indicates the expected values for the trophic level increase from the analysed fauna

Stable isotope values of fauna and human bone collagen. Lines indicate the mean without the outlier (δ13C = − 18.6‰; δ34S = 13.1‰) and two standard deviations (μ ± 2σ)
Amongst the humans sampled there is an outlier, a male young adult. While his bone collagen δ15N (12.3‰) are amongst the highest, his bone collagen δ13C (− 15.4‰) is highly enriched (Fig. 1) and he displays bone collagen δ34S depleted (9.3%) compared with the other individuals (Fig. 2). Overall, bone collagen δ15N and δ34S recorded in humans are more variable than their bone collagen δ13C (Table 1). The females show higher variance in their bone collagen δ15N, while the males display a higher variance in their bone collagen δ34S (Table 1). There are no statistically significant differences (p > 0.05) in stable isotope values between sexes, social status, or for δ13C and δ34S between age groups. However, bone collagen δ15N recorded in the human skeletons display significant differences (p = 0.05) with age groups (Table 2).
The individuals for which it was possible to estimate sex and age are represented in Fig. 3 illustrating δ13C and δ15N differences in bone collagen between age groups. While the young adults are above or close to the mean values for δ13C (− 18.6‰) and δ15N (10.8‰), the old adults are all under the mean values for both δ13C and δ15N (with the exception of one), but most fall within two standard deviations from the mean. There are no differences in bone collagen δ34S between the age groups (Fig. 4).

Stable isotope values of individuals with estimated sex and age. Lines indicate the mean and two standard deviations (μ ± 2σ) for all the samples except the outlier (δ13C = − 18.6 ± 1.0‰; δ15N = 10.8 ± 1.7‰), the young (δ13C = −18.4 ± 0.9‰; δ15N = 11.4 ± 1.4‰) and the old (δ13C = −18.7 ± 0.5‰; δ15N = 10.6 ± 1.8‰) adults

Stable isotope values (δ34S, δ15N and δ13C) of individuals from different social status and with estimated age (young, mature and old adults)
Discussion
General diet at Tomar
There might be dietary differences between different chronologies, especially after the sixteenth century with the introduction of new food sources, like C4 plants from America, but unfortunately, it was not possible to decrease the chronological interval estimated for Tomar (eleventh–seventeenth centuries). The high density of burials and the fact that Christian burials usually do not have associated artefacts, did not allow reliable dating.
Herbivores’ δ13C (− 21.3 to − 20.1‰, Fig. 1, Annex, Table A1) suggest a diet based on C3 plants (Vogel 1978; Schoeninger and DeNiro 1984; Chisholm 1989). Despite the wide range of the estimated chronology for Tomar’s necropolis (eleventh to seventeenth centuries) and the possibility that the analysed fauna represents different times (areas 14, 17 and 20), the herbivores’ δ13C are similar, arguing against the introduction of new food sources like maize (C4 plants). In contrast, bone collagen δ15N recorded in herbivores are more variable (4.8 to 7.9‰, Fig. 1) and with some enrichment, particularly observed for the Ovicapridae. Enrichment in faunal bone collagen δ15N may be related to variable animal husbandry practices and land management. Manured soils raise δ15N in soil and plants (van Klinken et al. 2000; Bogaard et al. 2007), having an impact on the local food web. δ15N enrichment is particularly evident between the Ovicapridae, which may be related to different food sources for sheep (grass; hay) and goats (bushes; tree leaves/bark). The Ovicapridae have higher δ15N than Bos. Higher δ15N in Ovicapridae compared with Bos are also observed in faunal remains (Annex, Fig. A2) from Koksijde (Polet and Katzenberg 2003) but not from Benipeixcar (Alexander et al. 2015). The domestic and wild Sus have similar δ13C and δ15N to the herbivores, as well as the only carnivore analysed (Canidae), suggesting a diet poor in animal protein. While pigs are frequently kept as herbivores (e.g. Quirós Castillo 2013), dog isotopic ratios usually cluster with the humans (e.g. Halffman and Velemínský 2015; Quirós Castillo 2013; Lubritto et al. 2013). As dogs frequently eat food scraps, their isotope values can be indicative of a human diet poor in animal protein.
The mean increase from faunal (except the Equus and Canidae) to human remains is 2.3‰ (in some individuals, more than 3‰, Fig. 1) for δ13C and 4.9‰ for the δ15N. Some individuals have higher enrichment than would be expected within the food web: up to 1‰ for δ13C (Schoeninger et al. 1983; Minagawa and Wada 1984; Schoeninger and DeNiro 1984) and 3 to 5.7‰ for δ15N (van der Merwe and Vogel 1978; Ambrose and Norr 1993). The trophic level increase expected based on the faunal isotope values would be between 7.8 and 13.6‰ for δ15N and from − 21.3 to − 19.9‰ for δ13C (grey area at Fig. 1). While human δ15N at Tomar range between 9.0 and 12.5‰ and can be explained by the trophic increment, their δ13C vary between − 19.4 and − 17.3‰, being clearly enriched compared with the faunal remains recovered in Tomar. This δ13C enrichment observed in the human remains can reflect a diet complemented by some aquatic (Tauber 1981; Chisholm et al. 1982, 1983) or C4 plants (Vogel 1978; Chisholm 1989) intake. Due to the presence of oysters and cockles shells at the excavation (areas 14 and 17) and that the analysed fauna were feeding on C3 plants it is more probable that the δ13C enrichment in the human bone collagen is related to aquatic protein than C4 plants intake.
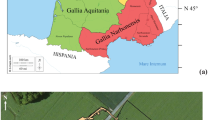
To better understand diet at Tomar, δ34S were also analysed for some faunal and human remains. Surprisingly, the fauna δ34S are higher than would be expected for terrestrial animals (δ34S > 12‰; Nehlich 2015) and correspond to values expected from coastal fauna influenced by marine sea spray (Nehlich 2015). However, Tomar is located at approximately 70 km from the coast and sea spray sulphates only reach up to 30 km inland (Wakshal and Nielsen 1982). Tide floods from the Atlantic Ocean increase the Tagus River flow and its salinity, reaching the floodplains near Santarém (Fig. 5) and increasing the sea spray reach but not enough to justify the δ34S registered for the fauna on its own.

Geological map of Tomar’s region. Tomar’s and Santarém surroundings represent the evaporites, gypsum and marl. 1, Nabão River; 2, Zêzere River; 3, Tagus River
Riverine sulphates can also be found on the riverbanks and floodplains, influencing the isotopic composition of the surrounding landscape (Fry 2002; Nehlich et al. 2011), and the values observed for Tomar’s fauna may be related to the livestock feeding on the floodplains. However, floodplains tend to have lower δ34S than areas further away from freshwater ecosystems (Nehlich et al. 2011). Therefore, it is possible that the use of algae as a fertiliser may have increased the δ34S in the food web as fresh seaweed can also be used to feed livestock, mostly ruminants and pigs (Chapman and Chapman 1980). In Portugal, algae has been used in agriculture previously to the fourteenth century (Veiga de Oliveira et al. 1975; Vieira and Santos 1995), but it would likely be restricted to coastal areas as algae are heavy and usually not carried very far inland (McHugh 2003). Even though the seaweed could be sundried and stored to be used as winter feedstuff for sheep and cattle (Evans and Critchley 2014), it would probably not be taken so far inland.
δ34S vary not only by dietary behaviour (Richards et al. 2001) but also by location (Hobson 1999), ranging from − 40 to + 40‰ in terrestrial rocks (Nielsen et al. 1991) and between − 20 and + 20‰ in terrestrial organic matter (Peterson and Fry 1987). The oxidation of sulphides and organic sulphur by microorganisms in the soils can also result in high δ34S and therefore influence the food web (Böttcher et al. 1998; Nehlich et al. 2011). Therefore, it is possible that the higher δ34S observed in the terrestrial fauna from Tomar may be related to the geochemistry of that area and not with agricultural or husbandry practices. Tomar is located at an area with evaporites, gypsum and marl (yellow area at Fig. 5) that would increase the δ34S in the food webs of this region. The Equus, with the lowest δ34S (10.2‰, Fig. 2), supports this hypothesis, as it was more mobile than the other domestic animals. The wild Sus also have lower δ34S (14.6 and 14.8‰), which can also be related with a higher mobility. Interestingly, the human collagen from Tomar has lower δ34S (9.3 to 15.6‰) than the faunal remains (13.1 to 18.5‰), suggesting that those terrestrial animals were not frequently consumed by the local population, who could have relied on other food sources from another geographical area with lower δ34S in its geo-ecosystem.
δ34S at riverine ecosystems usually fall between − 5 and + 15‰ (Nehlich 2015), but values can fall outside this range, depending on the geological surroundings of the river basin, ultimately influencing the δ34S of the river fauna (Nehlich et al. 2010; Nehlich et al. 2011). Unfortunately, fish bones were not recovered from Tomar’s excavation to confirm the values of the fish consumed by this population. If fresh water protein intake was important and with low δ34S, it could decrease the high δ34S within the surroundings of Tomar, related to its particular geological context. However, Nabão River, that crosses Tomar, is also located within the same geological substrate and thus, δ34S within its food webs are probably high. Zêzere River is located at approximately 10 km from Tomar rising at Serra da Estrela (a granitic and metamorphic mountain range) and meeting the Tagus River (an international river) at about 15 km from Tomar (Fig. 5). Since Zêzere and Tagus rivers do not pass through an area with evaporites and gypsum, δ34S of their food webs are probably lower than those at Nabão River. Human bone collagen δ34S suggest that if they were eating fresh water protein it was probably coming from Zêzere and/or Tagus. Besides, their larger dimensions could offer more food sources than Nabão River. The presence of shells at the excavation suggests also some marine protein intake. Nazaré is the closest coastal town where today fish and octopus are still sundried at the beach, this way of preserving the fish might have allowed its consumption further inland, in towns like Tomar, alongside with fish from the surrounding rivers.
The lower δ34S registered in human bone collagen can also be related with terrestrial intake from a geographical location with lower δ34S in its food webs. Since the staple Medieval diet in Portugal was bread (Vicente 2013), it is possible that it was being made with flour from cereals grown in a location different from Tomar’s surroundings. If bread, made with cereals with low δ34S, was consumed in high quantities, it could also have lowered the δ34S of individuals, independently of the geological substrate in the surroundings of Tomar. The possibility of C4 plants being consumed only by humans cannot be excluded. It could have been entering their diet in the form of maize flour, for example, and if the maize was not cultivated in Tomar, it could explain both the lower δ34S and the higher δ13C recorded in human bone collagen. However, the negative relation between δ34S and δ15N (Fig. 4) likely indicates that the higher δ15N are related with protein with lower δ34S and therefore the high δ15N represent protein from fresh water rather than from terrestrial fauna.
As Tomar was ruled by the Order of the Temple and later the Order of Christ (Vicente 2013), it is possible that religious dietary restrictions would be reflected in Tomar’s population. Also, in Tomar, merchants, crafters and farmers participated actively at local army levels alongside with knights, raising their status (Conde 1996) and probably giving them access to similar food resources. Müldner et al. (2009) found isotopically distinct diets between bishops and the general population in Scotland, the latter having higher fish intake, related to religious fasting. These dietary restrictions may have led to a higher intake of aquatic protein when meat consumption was not allowed with towns controlled by military orders likely being under increased pressure to follow religious dietary restrictions. More isotopic data from different places with similar chronologies is necessary to understand if the high intake of aquatic protein is due to the presence of the military orders at Tomar or if it was a generalised religious phenomenon.
Human diet at Medieval Tomar was complex and likely included food sources from outside Tomar. The general diet was poor in terrestrial protein and rich in fresh water protein with possible terrestrial protein from other geographical locations.
Dietary differences within Tomar
Even though some historical (e.g. Adamson 2004) and anthropological (e.g. Kjellström et al. 2009; Linderholm et al. 2008; Polet and Katzenberg 2003; Schutkowski et al. 1999; Reitsema et al. 2010; Reitsema and Vercellotti 2012) sources suggest different food access based on age, sex and status in Medieval times, that was not observed in Tomar. When the skeletons sampled were grouped by sex, age or inferred social status, only δ15N in the age groups was statistically different (p < 0.05; Table 2).
The bone collagen of young adults display higher δ15N than the old adults (Fig. 4), suggesting a higher animal protein intake for the young individuals. Since only skeletons without signs of physiological stress were sampled, the higher δ15N for the young adults is not related to chronic stress (Steele and Daniel 1978; Hobson et al. 1993; Gaye-Siessegger et al. 2004; Fuller et al. 2005; Deschner et al. 2012; D'Ortenzio et al. 2015) that might have resulted in premature death, due to ill health (Wood et al. 1992). These isotopic differences between young and old adults may be related to severe tooth loss that was observed in old individuals who therefore may have had increased difficulty ingesting some foods along with changes associated with metabolism in the ageing, such as reductions in taste, smell and hunger, and delayed rate of absorption (Roberts and Rosenberg 2006). The amount of freshwater fish is variable and not related to sex, social status or age (Fig. 4). Overall, the skeletons analysed had similar diets with smaller δ34S differences compared with other European samples (e.g. Nehlich et al. 2011), despite the wide chronology estimated for Tomar’s necropolis (eleventh–seventeenth centuries).There are no δ34S differences between age, sex or social status but there could be dietary differences between chronologies. Unfortunately, it was not possible do date the faunal or human remains.
The absence of statistically significant isotopic differences between sexes (Table 2) suggests that males and females had similar protein intakes at Tomar, however, sample sizes may be too small and dispersed (sex, age, social status and chronology) to detect significant differences. The uniform spatial distribution for males and females within the graveyard and the use of structured graves for both sexes also suggest that, at least at death, social status was not dependent on sex though Medieval society was male dominant.
The only outlier analysed, a young adult male, has higher values of both δ15N (12.3‰) and δ13C (− 15.4‰) and low δ34S (9.3‰). The low δ34S suggests that this individual might be an outsider, coming from a place with lower δ34S in its ecosystem, but the possibility of these isotope values being the result of a high freshwater protein intake from low δ34S cannot be excluded. The high δ13C and δ15N can also represent a terrestrial diet rich in C4 plants, directly or fed to the livestock, particularly if this individual was from a different geographical location.
Food can reflect social status and define social, cultural and religious boundaries (e.g. Thomas 2007; Curet and Pestle 2010); however, this was not observed within the samples analysed. The δ13C, δ15N and δ34S distribution is uniform when comparing individuals with higher and lower social status. Individuals from lower social status may be more susceptible to physiological stress, particularly due to malnutrition (e.g. Weston 2012). Since only individuals without skeletal signs of physiological stress (low stature, criba orbitalia, porotic hyperostosis) were sampled, the ones with lower social status could have been avoided. Adult stature is determined by genetics but also has an environmental determinant (e.g. Haviland 1967; Larsen 1997; Bogin 1999; Cardoso and Gomes 2009). The areas further away from the church (1st phase of the excavation; Fig. 1) better represent the individuals from the lower social status and the ones buried inside the church would be a better example of the people from higher social status. Therefore, the individuals analysed probably represent the average population and neither of the social extremes. More isotopic data from different social and health status would help understand if the diet at Tomar was uniform or if our results were biased by selecting only apparently healthy individuals.
Other European studies
Comparing the data with other Late Medieval European samples (Fig. 6), those with similar δ13C and δ15N averages are Koksijde (from a coastal Belgian monastery; Polet and Katzenberg 2003) and Rome (from an Italian mass grave; Salamon et al. 2008). Contrary with what would be expected, the stable isotope values from Tomar are closer to the Belgian sample (Polet and Katzenberg 2003) than to the other Iberian samples (Lubritto et al. 2013; Alexander et al. 2015), which may be related to religious dietary requirements, particularly low meat consumption, as the Belgian sample represents a monastic community (Polet and Katzenberg 2003). The similar faunal values for Tomar and Koksijde (Annex, Fig. A2) allow a comparison between the two locations, despite their geographical and social differences. The impact of religious directives of the Catholic Church on the diet has been registered before (Salamon et al. 2008). This was facilitated by industrial-scale fishing in the Atlantic (Barret et al. 2004) and improvement of food preservation methods (Heinrich 1986). Müldner and Richards (2007) also associated the increased intake of aquatic protein (mostly marine fish with some freshwater fish or molluscs) at St. Andrew (Fig. 6) with religious dietary habits in Later Middle Ages. Agricultural and husbandry practices used during the Middle Ages may also explain the different isotopic values between the Medieval skeletons buried in Tomar and the Prehistoric ones, alongside with a higher aquatic protein intake (Fig. 6).

Carbon and nitrogen stable isotope comparison between Prehistoric and Late medieval Tomar and other Late medieval European samples. Portugal: Tomar (this study) and Tomar Prehistoric (n = 2, Abrigo do Morgado Superior, unpublished data). Spain: Zaballa (n = 14, tenth–fifteenth centuries, Lubritto et al. 2013), Treviño (n = 15, twelfth–fourteenth centuries, Quirós Castillo 2013), Zornoztegi (n = 7, twelfth–fourteenth centuries, Quirós Castillo 2013), Colegiata St. Maria (n = 24, thirteenth–sixteenth centuries, Alexander et al. 2015) and Benipeixcar (n = 20, fifteenth–sixteenth centuries, Alexander et al. 2015). Italy: Rome (n = 29, fifteenth century, Salamon et al. 2008) and Trino Vercellese (n = 30, eighth–thirteenth centuries, Reitsema and Vercellotti 2012). Poland: Giecz (n = 24, eleventh–twelfth centuries, Reitsema et al. 2010). Belgium: Koksijde (n = 19, twelfth–fifteenth centuries, Polet and Katzenberg 2003). England: St. Andrew (n = 155, thirteenth–sixteenth centuries, Müldner and Richards 2007)
Out of the Iberian samples compared, Tomar has the highest δ15N mean, particularly when compared with Zaballa (Lubritto et al. 2013), Treviño (Quirós Castillo 2013) and Zornoztegi (Quirós Castillo 2013). The high δ15N mean can represent high animal protein intake, however, δ34S suggests a high aquatic protein intake which can also be related with higher δ15N. The faunal remains recovered from Zaballa (Lubritto et al. 2013), Treviño (Quirós Castillo 2013) and Zornoztegi (Quirós Castillo 2013) also have lower δ15N when compared with the ones from Tomar. Colegiata St. Maria (Alexander et al. 2015) and Benipeixcar (Alexander et al. 2015) have similar δ15N mean to Tomar’s but higher δ13C, which the authors relate to C4 plants consumption (directly or fed to domestic animals) or marine fish intake. It is also important to note the different locations of the Spanish collections. While Zaballa, Treviño and Zornoztegi are located at the Basque Country, Northeast of Spain, at approximately 90 km to the North Atlantic Ocean, the collections from Colegiata St. Maria and Benipeixcar are from Catalonia, South East of Spain, and at approximately 5 km to the Mediterranean Sea. The different locations of these collections may explain why Colegiata St. Maria and Benipeixcar may have higher aquatic protein intake than Zaballa, Treviño and Zornoztegi. Tomar’s population has closer mean δ15N to the ones closer to the Mediterranean Sea, suggesting also a higher intake of aquatic protein, while the different δ13C may be related to C4 plant consumption at Colegiata St. Maria and Benipeixcar (Salamon et al. 2008).
Conclusion
This study is part of a larger project comparing stable isotopic data from individuals without skeletal lesions compatible with diseases and/or physiological stress (presented here) and those with signs of infectious diseases. Since skeletons with lesions were not analysed, this study might better represent the diet at Tomar, instead of metabolic changes during physiological stress. The bone collagen stable isotope values (δ13C, δ15N and δ34S) suggest that individuals in Tomar had a complex diet, low in terrestrial animal protein and high in aquatic protein intake, despite its inland location, which could be related to the presence of the military orders in the town and more strict religious dietary restrictions. Dietary differences between sex or social status were not observed for the population of Tomar, but the quantity of aquatic protein intake is variable, with δ34S ranging from 11.4 to 15.6‰ (excluding the outlier). Diet appears to be very diverse in Medieval Iberia. Isotopic data from more archaeological sites are necessary to better understand how diet represents social, religious and economic factors, as well as increase our knowledge of trade, agricultural and husbandry practices in Medieval times. Data from archaeological sites near Tomar would also help understanding the impact of the presence of religious orders on a town’s general population.
References
Adamson MW (2004) Food in medieval times. Greenwood Publishing Group
Alexander MM, Gerrard CM, Gutiérrez A, Millard AR (2015) Diet, society, and economy in late Medieval Spain: stable isotope evidence from Muslims and Christians from Gandía, Valencia. Am J Phys Anthropol 156(2):263–273
Allen LH, Uauy R (1994) Guidelines for the study of mechanisms involved in the prevention or reversal of linear growth retardation in developing countries. Eur J Clin Nutr 48(Suppl 1):S212–S216
Ambrose SH, Norr L (1993) Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate. Prehistoric human bone. Springer, Berlin, pp 1–37
Barber M (2012) The new knighthood: a history of the order of the temple. Cambridge University Press, Cambridge
Barber M, Bate K (2002) The Templars: selected sources. Manchester University Press, Manchester
Barrett JH, Richards MP (2004) Identity, gender, religion and economy: new isotope and radiocarbon evidence for marine resource intensification in early historic Orkney, Scotland, UK. European Journal of Archaeology 7: 249–271
Binski P (1996) In: O’Connor M (ed) Medieval death: ritual and representation. British Museum Press, London
Bocherens H, Drucker D (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13(1–2):46–53
Bogaard A, Heaton TH, Poulton P, Merbach I (2007) The impact of manuring on nitrogen isotope ratios in cereals: archaeological implications for reconstruction of diet and crop management practices. J Archaeol Sci 34(3):335–343
Bogin B (1999) Patterns of human growth. Cambridge University Press, Cambridge
Böttcher ME, Oelschläger B, Höpner T, Brumsack HJ, Rullkötter J (1998) Sulfate reduction related to the early diagenetic degradation of organic matter and “black spot” formation in tidal sandflats of the German Wadden Sea (southern North Sea): stable isotope (13C, 34S, 18O) and other geochemical results. Org Geochem 29(5):1517–1530
Brooks S, Suchey JM (1990) Skeletal age determination based on the os pubis: a comparison of the Acsádi-Nemeskéri and Suchey-Brooks methods. Hum Evol 5(3):227–238
Brown TA, Nelson DE, Vogel JS, Southon JR (1988) Improved collagen extraction by modified Longin method. Radiocarbon 30(2):171–177
Buikstra JE, Ubelaker DH (1994) Standards for data collection from human skeletal remains
Cardoso H, Gomes J (2009) Trends in adult stature of peoples who inhabited the modern Portuguese territory from the mesolithic to the late 20th century. Int J Osteoarchaeol 19(6):711–725
Chapman VJ, Chapman DJ (1980) Seaweeds and their uses. Chapman and Hall, London
Chisholm BS (1989) Variation in diet reconstructions based on stable carbon isotopic evidence. In: Price TD (ed) The chemistry of Prehistoric human bone. Cambridge University Press, Cambridge, pp 10–37
Chisholm BS, Nelson DE, Hobson KA, Schwarcz HP, Knyf M (1983) Carbon isotope measurement techniques for bone collagen: notes for the archaeologist. J Archaeol Sci 10(4):355–360
Chisholm BS, Nelson DE, Schwarcz HP (1982) Stable-carbon isotope ratios as a measure of marine versus terrestrial protein in ancient diets. Science (New York, NY) 216(4550):1131–1132
Conde MAS (1996) Tomar Medieval. O espaço e oshomens, Cascais
Curet LA, Pestle WJ (2010) Identifying high-status foods in the archeological record. J Anthropol Archaeol 29(4):413–431
Daniell C (1998) Death and burial in Medieval England, 1066–1550. Routledge, London
DeNiro MJ (1985) Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317(6040):806–809
DeNiro MJ, Epstein S (1976) You are what you eat (plus a few‰): the carbon isotope cycle in food chains. Geol Soc Am 6:834
DeNiro MJ, Epstein S (1978) Influence of diet on the distribution of carbon isotopes in animals. Geochimica EtCosmochimicaActa 42(5):495–506
DeNiro MJ, Epstein S (1981) Influence of diet on the distribution of nitrogen isotopes in animals. Geochimica EtCosmochimicaActa 45(3):341–351
Deschner T, Fuller BT, Oelze VM, Boesch C, Hublin J, Mundry R, Hohmann G (2012) Identification of energy consumption and nutritional stress by isotopic and elemental analysis of urine in bonobos (Pan paniscus). Rapid Commun Mass Spectrom 26(1):69–77
D'Ortenzio L, Brickley M, Schwarcz H, Prowse T (2015) You are not what you eat during physiological stress: isotopic evaluation of human hair. Am J Phys Anthropol 157(3):374–388
Dufour E, Bocherens H, Mariotti A (1999) Palaeodietary implications of isotopic variability in Eurasian lacustrine fish. J Archaeol Sci 26(6):617–627
Evans FD, Critchley AT (2014) Seaweeds for animal production use. J Appl Phycol 26(2):891–899
França JA (1994) Cidades e Vilas de Portugal: Tomar, vol 18. Lisboa, Editorial Presença
Fry B (2002) Conservative mixing of stable isotopes across estuarine salinity gradients: a conceptual framework for monitoring watershed influences on downstream fisheries production. Estuar Coasts 25(2):264–271
Fuller BT, Fuller JL, Sage NE, Harris DA, O’Connell TC, Hedges RE (2005) Nitrogen balance and δ15N: why you’re not what you eat during nutritional stress. Rapid Commun Mass Spectrom 19(18):2497–2506
Gaye-Siessegger J, Focken U, Abel H, Becker K (2004) Individual protein balance strongly influences δ15N and δ13C values in Nile Tilapia, oreochromisniloticus. Naturwissenschaften 91(2):90–93
Gonçalves I (2004) Entre aabundância e a miséria: as práticasalimentares da Idade Médiaportuguesa. Estudosmedievais. Quotidiano Medieval: imaginário, representação e práticas. Livros Horizonte, Lisboa
Grau-Sologestoa I (2017) Socio-economic status and religious identity in Medieval Iberia: the zooarchaeological evidence. Environ Archaeol 22(2):189–199
Graves C (1989) Social space in the English Medieval parish-church. Econ Soc 18(3):297–322
Halffman CM, Velemínský P (2015) Stable isotope evidence for diet in early medieval Great Moravia (Czech Republic). Journal of Archaeological Science: Reports, 2, pp.1–8
Haviland WA (1967) Stature at Tikal, Guatemala: implications for ancient Maya demography and social organization. Am Antiq 32:316–325
Heinrich D (1986) Fishing and consumption of cod (Gadus morhua Linnaeus, 1758) in the Middle Ages. In: Brinkhuizen, D.C., Clason, A.T. (Eds.), Fish and Archaeology. British Archaeological Reports, Oxford, pp. 42–52
Hobson KA (1999) Tracing origins and migration of wildlife using stable isotopes: a review. Oecologia 120(3):314–326
Hobson KA, Alisauskas RT, Clark RG (1993) Stable-nitrogen isotope enrichment in avian tissues due to fasting and nutritional stress: implications for isotopic analyses of diet. Condor 95:388–394
Hobson KA, Clark RG (1992) Assessing avian diets using stable isotopes I: turnover of 13C in tissues. Condor 94:181–188
Katzenberg MA, Weber A (1999) Stable isotope ecology and palaeodiet in the lake Baikal region of Siberia. J Archaeol Sci 26(6):651–659
Kjellström A, Storå J, Possnert G, Linderholm A (2009) Dietary patterns and social structures in Medieval Sigtuna, Sweden, as reflected in stable isotope values in human skeletal remains. J Archaeol Sci 36(12):2689–2699
Larsen C (1997) Bioarchaeology: interpreting behavior from the human skeleton. Cambridge University Press, New York
Leach BF, Quinn CJ, Lyon GL (1996) A stochastic approach to the reconstruction of Prehistoric human diet in the pacific region from bone isotope signatures. Tuhinga 1–54
Linderholm A, Jonson CH, Svensk O, Liden K (2008) Diet and status in Birka: stable isotopes and grave goods compared. Antiquity 82:446–461
Longin R (1971) New method of collagen extraction for radiocarbon dating. Nature 230(5291):241–242
Lovejoy CO, Meindl RS, Pryzbeck TR, Mensforth RP (1985) Chronological metamorphosis of the auricular surface of the ilium: a new method for the determination of adult skeletal age at death. Am J Phys Anthropol 68(1):15–28
Lubritto C, Sirignano C, Ricci P, Passariello I, Castillo JQ (2013) Radiocarbon chronology and paleodiet studies on the Medieval rural site of Zaballa (Spain): preliminary insights into the social archaeology of the site. Radiocarbon 55(3):1222–1232
Malgosa A (2011) The Middle Ages viewed through Physical Anthropology. Imago Temporis. Medium Aevum, 5
McHugh DJ (2003) A guide to the seaweed industry. Food and Agriculture Organization of the United Nations, Rome
Minagawa M, Wada E (1984) Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age. Geochimica EtCosmochimicaActa 48(5):1135–1140
Moore M, Ross A (2013) Stature estimation. In: DiGangi E (ed) Research methods in human skeletal biology. Elsevier, Waltham, pp 151–180
Morris D, McAlpin M (1979) Measuring the condition of the world’s poor. Pergamon Press, New York
Müldner G, Richards MP (2007) Diet and diversity at later Medieval Fishergate: the isotopic evidence. Am J Phys Anthropol 134(2):162–174
Müldner G, Montgomery J, Cook G, Ellam R, Gledhill A, Lowe C (2009) Isotopes and individuals: diet and mobility among the Medieval Bishops of Whithorn. Antiquity 83(322):1119–1133
Nehlich O (2015) The application of sulphur isotope analyses in archaeological research: a review. Earth Sci Rev 142:1–17
Nehlich O, Borić D, Stefanović S, Richards MP (2010) Sulphur isotope evidence for freshwater fish consumption: a case study from the Danube Gorges, SE Europe. J Archaeol Sci 37(5):1131–1139
Nehlich O, Fuller BT, Jay M, Mora A, Nicholson RA, Smith CI, Richards MP (2011) Application of sulphur isotope ratios to examine weaning patterns and freshwater fish consumption in roman Oxfordshire, UK. Geochimica EtCosmochimicaActa 75(17):4963–4977
Nehlich O, Richards MP (2009) Establishing collagen quality criteria for sulphur isotope analysis of archaeological bone collagen. Archaeol Anthropol Sci 1(1):59–75
Nielsen H., Pilot J, Grinenko LN, Grinenko VA, Lein AY, Smith JW, Pankina RG (1991) Lithospheric sources of sulphur. In: HR Krouse (ed) Stable isotopes: natural and anthropogenic sulphur in the environment
Ottaway P (1992) Archaeology in British towns: from the emperor Claudius to the Black Death. Routledge, London
Peterson BJ, Fry B (1987) Stable isotopes in ecosystem studies. Annu Rev Ecol Syst 18(1):293–320
Phenice TW (1969) A newly developed visual method of sexing the os pubis. Am J Phys Anthropol 30(2):297–301
Platt C (1981) The parish churches of Medieval England. Secker & Warburg, London
Polet C, Katzenberg MA (2003) Reconstruction of the diet in a mediaeval monastic community from the coast of Belgium. J Archaeol Sci 30(5):525–533
Quirós Castillo JA (2013) Los comportamientosalimentariosdelcampesinado medieval en el País Vasco y suentorno (siglos VIII–XIV). Historia Agraria 59:13–41
Reitsema LJ, Crews DE, Polcyn M (2010) Preliminary evidence for medieval Polish diet from carbon and nitrogen stable isotopes. J Archaeol Sci 37(7):1413–1423
Reitsema LJ, Vercellotti G (2012) Stable isotope evidence for sex-and status-based variations in diet and life history at medieval Trino Vercellese, Italy. Am J Phys Anthropol 148(4):589–600
Richards MP, Fuller BT, Hedges RE (2001) Sulphur isotopic variation in ancient bone collagen from Europe: implications for human palaeodiet, residence mobility, and modern pollutant studies. Earth Planet Sci Lett 191(3):185–190
Richards MP, Hedges RE (1999) Stable isotope evidence for similarities in the types of marine foods used by Late Mesolithic humans at sites along the Atlantic coast of Europe. J Archaeol Sci 26(6):717–722
Roberts C, Manchester K (2007) The archaeology of disease. Cornell University Press, Ithaca
Roberts SB, Rosenberg I (2006) Nutrition and aging: changes in the regulation of energy metabolism with aging. Physiol Rev 86(2):651–667
Salamon M, Coppa A, McCormick M, Rubini M, Vargiu R, Tuross N (2008) The consilience of historical and isotopic approaches in reconstructing the medieval Mediterranean diet. J Archaeol Sci 35(6):1667–1672
Scheuer L, Black S (2000) Development and ageing of the juvenile skeleton. Hum Osteol Archael Forensic Sci 9–21
Schoeninger MJ, DeNiro MJ (1984) Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochimica EtCosmochimicaActa 48(4):625–639
Schoeninger MJ, DeNiro MJ, Tauber H (1983) Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial components of Prehistoric human diet. Science (New York, NY) 220(4604):1381–1383
Schutkowski H, Herrmann B, Wiedemann F, Bocherens H, Grupe G (1999) Diet, status and decomposition at Weingarten: trace element and isotope analyses on early mediaeval skeletal material. J Archaeol Sci 26(6):675–685
Sponheimer M, Robinson T, Ayliffe L, Roeder B, Hammer J, Passey B, West A, Cerling T, Dearing D, Ehleringer J (2003) Nitrogen isotopes in mammalian herbivores: hair δ15N values from a controlled feeding study. Int J Osteoarchaeol 13(1–2):80–87
Steele K, Daniel RM (1978) Fractionation of nitrogen isotopes by animals: a further complication to the use of variations in the natural abundance of δ15N for tracer studies. J Agric Sci 90(01):7–9
Swanson RN (1989) Church and society in late medieval England. Blackwell, Oxford
Tauber H (1981) 13C evidence for dietary habits of Prehistoric man in Denmark. Nature 292(5821):332–333
Teeri J, Schoeller D (1979) δ13C values of an herbivore and the ratio of C3 to C4 plant carbon in its diet. Oecologia 39(2):197–200
Thomas RM (2007) Food and the maintenance of social boundaries in medieval England. The Archaeology of Food, and Identity. Occasional Papers 34:130–151
Tieszen LL, Boutton TW, Tesdahl K, Slade NA (1983) Fractionation and turnover of stable carbon isotopes in animal tissues: implications for δ13C analysis of diet. Oecologia 57(1–2):32–37
Valente J (1998) The new frontier: the role of the knightstemplar in the establishment of Portugal as an independent kingdom. Mediterr Stud 7:49–65
Van der Merwe NJ, Vogel JC (1978) 13C content of human collagen as a measure of Prehistoric diet in woodland North America. Nature 276(5690):815–816
van Klinken GJ, Richards MP, Hedges REM (2000) An overview of causes for stable isotopic variations in past European human populations: environmental, ecophysiological and cultural effects. In: Ambrose SH, Katzenberg MA (Eds.), Biogeochemical Approaches to Palaeodietary Analysis. Kluwer Academic/ Plenum Publishers, New York
Van Klinken GJ, Richards MP, Hedges BE (2002) An overview of causes for stable isotopic variations in past European human populations: environmental, ecophysiological, and cultural effects. Biogeochemical approaches to paleodietary analysis. Springer, Berlin, pp 39–63
Veiga de Oliveira E, Galhano F, Pereira B (1975) Actividades agro-marítimasem Portugal. Instituto de Alta Cultura, Lisboa
Vieira VV, Santos M (1995) Directório de aquacultura e biotecnologiamarinha. Escola Superior de Biotecnologia da Universidade Católica Portuguesa, Porto
Vicente M (2013) Entre Zêzere e Tejo, Propriedade e Povoamento Doutoramentoem Historia Medieval Universidade de Lisboa [Unpublished]
Vogel JC (1978) Isotopic assessment of the dietary habits of ungulates. S Afr J Sci 74(8):298–301
Wakshal E, Nielsen H (1982) Variations of δ34S (SO4), δ18O (H2O) and Cl/SO4 ratio in rainwater over northern Israel, from the Mediterranean Coast to Jordan Rift Valley and Golan Heights. Earth Planet Sci Lett 61(2):272–282
Walker PL, DeNiro MJ (1986) Stable nitrogen and carbon isotope ratios in bone collagen as indices of Prehistoric dietary dependence on marine and terrestrial resources in southern California. Am J Phys Anthropol 71(1):51–61
Weston DA (2012) Nonspecific infection in paleopathology: interpreting periosteal reactions. In: Grauer AL (ed) A companion to paleopathology. Wiley-Blackwell, pp 492–512
Wood J, Milner G, Harpending H, Weiss K (1992) The osteological paradox - problems of inferring Prehistoric health from skeletal samples. Curr Anthropol 33(4):343–370
Zohary T, Erez J, Gophen M, Berman-Frank I, Stiller M (1994) Seasonality of stable carbon isotopes within the pelagic food web of Lake Kinneret. Limnol Oceanogr 39(5):1030–1043
Acknowledgments
The authors would like to thank all the anthropologists and archaeologists involved at the excavation of Tomar’s, in particular to Sérgio Simões Pereira, Ricardo Ávila Ribeiro, Sónia Ferro, Cláudia Santos and Helena Santos for the maps and information about the excavation. We would like to thank Dr. Angela Lamb for her support at NERC facilities. We are also thankful to Rodrigo Maia for the sulphur isotope analysis, Prof José Mirão for his help understanding Tomar’s geochemistry and Dr. Ana Cruz for allowing us to compare our data with unpublished isotopic data from Prehistoric Tomar (Abrigo do Morgado Superior).
Funding
A Curto acknowledge the financial support of the University of Kent through the University of Kent 50th Anniversary Scholarship. A-F Maurrer and C. Barrocas-Dias acknowledge the financial support of FCT throughout project HERCULES Laboratory-Cultural Heritage Studies and Safeguard with reference UID/Multi/04449/2013.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
ESM 1
(DOCX 826 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Curto, A., Maurer, AF., Barrocas-Dias, C. et al. Did military orders influence the general population diet? Stable isotope analysis from Medieval Tomar, Portugal. Archaeol Anthropol Sci 11, 3797–3809 (2019). https://doi.org/10.1007/s12520-018-0637-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-018-0637-3