
Abstract
The combination of Trichoderma virens Gl006 and B. velezensis Bs006 as a consortium has high potential to control Fusarium wilt (FW) of cape gooseberry (Physalis peruviana) caused by Fusarium oxysporum f. sp. physali (Foph). However, the interactions between these two microorganisms that influence the biocontrol activity as a consortium have not been studied. Here, we studied the interactions between Gl006 and Bs006 that keep their compatibility under in vitro and greenhouse conditions. Antagonism tests between Gl006 and Bs006 inoculated both individually and in consortium against Foph strain Map5 was carried out on several solid media. The effect of supernatant of each selected microorganism on growth, conidia germination, biofilm formation and antagonistic activity on its partner was also studied. Biocontrol activity by different combinations of cells and supernatants from both microorganisms against Fusarium wilt was evaluated under greenhouse conditions. In vitro antagonism of the consortium against Foph showed a differential response among culture media and showed compatibility among BCA under nutritional conditions close to those of the rhizosphere. The supernatant of Bs006 did not affect the antagonistic activity of Gl006 and vice versa. However, the supernatant of Bs006 promoted the biocontrol activity of Gl006 in a synergistic way under greenhouse, reducing the disease severity by 71%. These results prove the compatibility between T. virens Gl006 and B. velezensis Bs006 as a potential tool to control Fusarium wilt of cape gooseberry.
Similar content being viewed by others

Introduction
Fusarium oxysporum is the causal agent of Fusarium wilt disease in several species of cultivated plants worldwide. This phytopathogen is in the fifth place within the top ten of the most important plant pathogens due to its effects in crops of economic importance causing severe losses1. Fusarium wilt is the main limitation of cape gooseberry (Physalis peruviana) crop in Colombia2. Wilt symptoms in field include wilting of top leaves, stunting of plants, lateral yellowing of branches and leaves, some plants present both dry and alive branches. Plants finally become yellow and die, resulting in plant losses and reduced fruit yield. This phytosanitary problem has also caused the migration of cropped areas within the country, but the problem persists in old and new cropped areas2. Currently, there are no registered phytosanitary products for the control of this disease in cape gooseberry.
In this context, biological control has emerged as an environmentally sustainable alternative. However, classical biological control in which just one biological control agent (BCA) is used has shown high variability among tests. Considering that biocontrol agents are living organisms and they may not be active in all agroecosystems3, many of the recent studies have focused on using combinations of BCA to increase the efficacy against phytopathogens. Those combinations are known as consortia. A consortium is a microbial association of two or more microorganisms, which could be archaea, fungi, bacteria, virus or algae4,5. However, there can be either direct or indirect interactions among the different populations of BCA in consortium, which could result in negative or positive effects on the biocontrol efficacy6. Therefore, in a combination of two BCA, the biocontrol activity can be increased, reduced or similar compared to individual activity.
Several studies have evaluated the efficacy of microorganisms in consortium, but few have evaluated how microorganisms interact each other and nor have identified the mechanisms by which the consortium has higher biocontrol activity than the individual components. Knowing these interactions will allow to design more effective consortia and take advantage of its potential. In the earliest studies of microbial consortia, the selection of pairs or mixtures was done according to the individual potential but did not mention whether there was an antagonistic or suppressive effect on its partner. The recommendation for current studies is to evaluate how the components of a consortium interact with each other, including how they interact at functional levels and to determine the predominant component of the consortium7.
There are studies about the synergistic or additive effects of application of microbial consortia containing two, three and even four microorganisms, the most frequent in these studies are Trichoderma spp., Pseudomonas spp., Bacillus spp., and Gliocladium spp. Mixtures of fungus - fungus, fungus - bacteria and bacteria – bacteria, combination of microorganisms belonging to the same genus or strains of the same species have been evaluated7. Some examples of these studies are the reduction of diseases caused by Fusarium solani and Rhizoctonia solani using Bacillus subtilis MBI600 and Rhizobium tropici UMR 1899 in bean8; the use of Trichoderma harzianum DB11 and Gliocladium catenulatum to control Phytophthora cactorum in strawberry9; the decrease of the mortality of pea plants due to Sclerotinia sclerotiorum using a consortium of Pseudomonas sp., Bacillus sp. and Trichoderma sp10; the combination of B. subtilis and Pseudomonas fluorescens against F. solani in bell pepper11; the reduction of the incidence of the disease caused by Rhizoctonia bataticola in cotton using a consortium of Azospirillum sp. AZ204 and P. fluorescens Pf112; and the combination of T. harzianum Tr16 and Pseudomonas sp. Ps14 against F. oxysporum in cucumber13.
Trichoderma spp. and Bacillus spp. are among the most commonly used microorganisms in biological control against F. oxysporum14 but few studies have evaluated consortia based on Trichoderma spp. and Bacillus spp. There have been evaluated different isolates of Trichoderma spp. and Bacillus spp. against R. solani and as plant growth promoters on cucumber and beans15. However, none of the combinations improved the efficacy showed by the individual activity of three isolates of Trichoderma spp. and only plant growth promotion by the consortium was observed. Some studies about the potential combination of Trichoderma spp. and Bacillus spp. have been conducted mainly in vitro. For instance, Wu et al.16 evaluated the antimicrobial effect of co-culturing Trichoderma asperellum GDFS1009 and Bacillus amyloliquefaciens ACCC11060 in BP broth (beef extract 0.3% and peptone 0.5%) finding the greatest effects against Botrytis cinerea by the supernatant from the co-culture than individual treatments. Other study demonstrated that in vitro co-culture of T. asperellum GDFS1009 and B. amyloliquefaciens 1841 induced the production of compounds not detected under pure cultures, attributed to the competition between them17. According to these findings, studies between Trichoderma spp. and Bacillus spp. are related to production of antimicrobial compounds in co-culture but the in vivo interactions between cells-supernatants or cells-cells of biocontrol agents have not been addressed. To the best of our knowledge, the work of Silva18 is the only one reporting the evaluation of in vitro compatibility between strains of Trichoderma spp. and strains of B. subtilis and Bacillus cereus as a method to screening potential consortia against FW in cape gooseberry. However, there are no published works on in vivo evaluations.
In previous studies the rhizobacteria Bacillus velezensis Bs006 was selected for its potential to control Fusarium wilt, but it has shown variable efficacy among tests2. Also, the combination of Trichoderma virens Gl006 and B. velezensis Bs006 increased the biocontrol activity and maintained consistently the effectiveness, against Fusarium wilt of cape gooseberry (Izquierdo-García et al. Unpublished). However, the interactions between these BCA remain unknown which would help to establish guidelines to make the consortium more effective. In this context, the aim of this study was to evaluate the compatibility among the biocontrol agents Gl006 and Bs006 and to know their interactions.
Results
Interactions between biological control agents under in vitro conditions
Effect of supernatants on the growth of biological control agents
Soaking the conidia of T. virens Gl006 during 48 h in Bs006-sterile supernatant solution at different concentrations (1 to 30%) did not affect its growth (Supplementary material - Figure S1a). It was observed that the concentrations of 1 and 20% stimulated the growth of Gl006 at 24 hours of exposition, but this effect was transient since at 48 hours there were no significant differences among the treatments (Fig. 1a). On the other hand, all concentrations of GI006 supernatant delayed Bs006 growth in a concentration dependent manner, starting at eight hours of fermentation (Fig. 1b, Supplementary material - Figure S1b).

(a) Growth of T. virens Gl006 in PDB medium supplemented with different concentrations (1, 5, 10, 20 and 30%) of B. velezensis Bs006 cell-free supernatant. (b) Growth of B. velezensis Bs006 in LB medium supplemented with 1, 5, 10, 20 and 30% of T. virens Gl006 cell-free supernatant. Growth was expressed as optical density (OD630 nm) measured in a microplate reader at 24 and 48 hours. Columns with the same letter are not significantly different according to Tukey test (α = 0.05) at each time of evaluation. Bars on the columns represent standard deviation (n = 3).
Germination of Gl006 conidia and viability of Bs006 treated with supernatants
Germination of the exposed conidia of Gl006 to Bs006-supernatant showed values close to 90% on water-agar medium after 24 hours of incubation. Thus, none of the Bs006-supernatant concentrations caused negative effects on Gl006 conidia after six hours of exposure. Same results were observed regarding the viability of Bs006 cells exposed to the Gl006-supernatant and none of the concentrations affected to Bs006 (data not shown).
Effect of supernatants on the antagonistic activity
Antagonistic activity of Gl006 was not affected after six hours of exposure of conidia to Bs006-supernatant in concentrations of 1 to 30%. Inhibition of Foph growth by Gl006-treated conidia was in a range of 76 to 78% (Fig. 2). Similar results were observed when Bs006 cells were exposed to the Gl006-supernatant, since exposed cells reduced the growth of Foph in a range of 52 to 54% (Fig. 3).

Antagonistic activity of T. virens Gl006 after being exposed to Bs006-supernatant in dual confrontation test against F. oxysporum f. sp. physali strain Map5 (Foph). Gl006 (G) conidia were exposed to Bs006 cell-free supernatant (Sup) from 1 to 30%. Growth inhibition of Foph was measured according to diametral growth in the positive control - F in PDA medium, after seven days of incubation at 25 °C.

Antagonistic activity of Bs006 after being exposed to Gl006-supernatant in dual confrontation test against F. oxysporum f. sp. physali strain Map5 (Foph). Bs006 (B) cells exposed for six hours to Gl006 cell-free supernatant (Sup) from 1 to 30%. Growth inhibition of Foph was measured according to diametral growth in the positive control - F in PDA medium after seven days of incubation at 25 °C.
Biofilm formation by B. velezensis Bs006 treated with Gl006-supernatant and under co-culture
It was observed that continuous exposition of Bs006 to Gl006-supernatant for 24 hours affected the ability of the bacteria to form biofilms in a concentration dependent manner. The exposure of Bs006 to 1% of Gl006-supernatant did not show negative effects on biofilm formation, but higher concentrations of the supernatant affected significantly this trait (Fig. 4a). Similar results were observed regarding the evaluation of co-cultures of Gl006 with Bs006, where the lowest concentration of Gl006 (1 × 105 conidia.mL−1) did not affect biofilm formation by Bs006, but this performance decreased when the bacteria grew with Gl006 in the same medium at 1 × 106 and 1 × 107 conidia.mL−1 (Fig. 4b).

Biofilm formation of B. velezensis Bs006 in presence of T. virens Gl006 supernatant (a) and conidia (b). B. velezensis Bs006 was grown for 24 h in LB medium supplemented with Gl006-supernatant from 1 to 30% (SupGl1 to SupGl30) or viable conidia of T. virens Gl006 at 105 to 107 conidia.mL−1 (CoGlE5 – CoGlE7). Control: Growth of Bs006 in LB free of Gl006-supernatant and conidia. Columns with the same letter are not significantly different according to Tukey test (α = 0.05). Bars on the columns represent standard deviation (n = 9).
Effect of co-cultures of Bs006 and Gl006 on the antagonistic activity on different solid media
The highest percentage of growth inhibition of F. oxysporum Foph on solid media was caused by Gl006 alone (>70%) in all culture media. Differential effect among culture media was observed in dual confrontation tests of Gl006 and Bs006 co-culture against Foph. Thereby, the diameter of the Foph colony was similarly reduced by the treatments Gl006, Bs006 and the combination of Gl006 with Bs006 on synthetic media (LB, PDA and the combination of LB with PDA) (Fig. 5), suggesting that under these conditions there was a neutral interaction between both biocontrol agents. In contrast, Bs006 did not grow and did not reduce Foph growth on soil solution-agar (SS), cape gooseberry root exudates prepared on SS-agar (SCRE) and cape gooseberry root exudates-agar (CRE) media, whereas Gl006 colonized the surface of all these media and inhibited Fusarium in both treatments, Gl006 individual and in consortium with Bs006 (Fig. 5), suggesting that Foph inhibition in such conditions was caused by Gl006. But on the other side, it was observed that these three treatments reduced the growth of Foph in media containing only artificial root exudates (ARE) or artificial root exudates prepared in soil solution (SARE), with Gl006 showing the highest growth inhibition of Foph (96 and 75% respectively) (Fig. 5). Bs006 grew and inhibited the growth of Fusarium between 45 to 50% on ARE and SARE solid media. In this bioassay there was no additive or synergistic effect by the consortium.

Growth inhibition of Foph colony in solid media by Gl006, Bs006 and the consortium. (ARE), artificial root exudates. soil solution + synthetic root exudates (SARE), soil solution (SS). Luria Bertani-Agar (LBA), Potato-Dextrose-Agar (PDA), PDA (50%) + LBA (50%) (LBA+PDA), Cape gooseberry root exudates (CRE), soil solution + Cape gooseberry root exudates (SCRE). Bars on the columns represent standard deviation between experimental units of three replicates in the time (n = 12). Columns with the same letter are not significantly different according to Tukey test (α = 0.05).
On the other hand, the co-inoculation of Foph, Gl006 and Bs006 on solid media (i.e. all together in the same suspension) showed up to 100% inhibition of Foph, also depending on the culture medium. Thereby Bs006 grew only on synthetic media, while growth of the two biocontrol agents was observed on the media containing soil solution and root exudates (ARE and SARE) (Table 1). Moreover, it was found that Bs006 inhibited the growth of Gl006 by 60% on synthetic media PDB and LB in the dual confrontation test of both BCA. Whilst the inhibition was 50% on SARE and ARE media. In contrast, Bs006 did not inhibit the growth of Gl006 on SS and cape gooseberry root exudates media CRE and SCRE (Supplementary material - Figure S2).
Microscopic observations of the Gl006 and Bs006 interactions
Interactions on solid media
Inoculation of Gl006 and Bs006 on jellified cape gooseberry root exudates allowed to observe that Gl006 conidia germinated and their hyphae extended to the interaction zone of both microorganisms (Supplementary material - Figure S3). Bs006 cells were adhered both to the non-germinated and germinated conidia. Bs006 cells also were adhered to the Gl006 hyphae forming aggregates like a biofilm (Fig. 6). No damages on fungal structures were observed.

Microscopy observations of the interaction between T. virens Gl006 and B. velezensis Bs006 in jellified cape gooseberry root exudates under fluorescent microscope. (a) Germinated conidia of T. virens Gl006 in absence of B. velezensis. (b–d) Interaction between Gl006 conidia and Bs006 cells. Yellow arrows indicate Bs006 spores at the beginning of biofilm formation on Gl006 surface after 72 hours contact.
In planta interactions
It was observed that Gl006 colonized the seed-surface forming a net (Fig. 7). Mycelium of Gl006 was observed around the root hairs. However, some conidia did not germinate at 24 h after inoculation. On the other hand, Bs006 made a biofilm on the surface of both the seed and the root (Fig. 7). When Gl006 and Bs006 were inoculated together on the host surface, it was noticed that both microorganisms colonized the surface of the plant tissue, Gl006 formed a net and Bs006 a biofilm (Fig. 7).

Environmental scanning microscopy images of non-germinated and germinated seeds of cape gooseberry inoculated with T. virens Gl006 and B. velezensis Bs006. (a) Surface of seed coat without inoculations, (b and c) co-inoculated seed with Gl006 and Bs006, (d) co-inoculated seedling with Bs006, Gl006 and Foph. Bs006: B. velezensis Bs006 cells, Gl006: T. virens Gl006 hypha, Foph: F. oxysporum f. sp. physali-Map5. Samples were observed at 24 hours after inoculation and incubation at 25 °C.
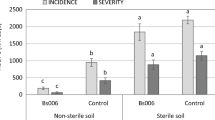
Biocontrol activity of Gl006 and Bs006 consortium against Fusarium wilt disease under greenhouse conditions
Application of Bs006 cells suspension prepared in Gl006-supernatant solution and vice versa and the combination of cells and supernatants of both microorganisms to the soil, showed that Bs006 and Gl006 cells, supernatants, and Gl006 conidia resuspended in Bs006-supernatant significantly reduced the progress of both Fusarium wilt incidence (Supplementary material - Figure S4) and severity (Fig. 8), as compared to the negative control. Interestingly, Bs006 cells suspended in its own supernatant showed reduced effectivity against FW, compared to the application of Bs006-supernatant alone and the Bs006 cells suspension in water (Fig. 8, Table 2). Whereas the suspension of Bs006 in Gl006-supernatant decreased the efficacy against Fusarium wilt. The application of Gl006 conidia suspended in water or its own supernatant, as well as the application of Gl006-supernatant, significantly reduced the progress of the disease, in a similar way to the combination of components of the two biocontrol agents (Fig. 8, Table 2). The combination of Bs006-supernatant with Gl006-conidia showed synergy in the reduction of the disease severity progress at 50 days after the application of treatments (Table 2).

Effect of the combination of cells and supernatants of Gl006 and Bs006 on the progress of Fusarium wilt severity. Columns with the same letter are not significantly different according to Duncan's multiple range test (α = 0.05). Con = Conidia, Cel = cells, Bs = B. velezensis Bs006, Gl = T. virens Gl006. Sup = cell-free supernatant.
Discussion
In most of the published studies, initial screenings of mixtures of biological control agents were carried out under laboratory conditions, on synthetic media used regularly for the growth of bacteria and fungi, such as nutritive medium, LB or PDA. According to Weller (1988)19, in several cases the best biocontrol agents in planta are not those showing the highest inhibition of pathogens in vitro and that is why new bioassays with conditions close to natural ones have been designed, in which fragments or the entire host are used.
In this work we found that the culture medium influenced the response of the compatibility between T. virens Gl006 and B. velezensis Bs006, since the synthetic media favored the growth of the bacteria, which completely inhibited the fungus growth. Contrary to what was observed in the medium containing soil solution and root exudates from cape gooseberry, in which only Gl006 grew. In this respect, it is known that Trichoderma spp. colonizes nutrient-poor environments, and even nutritional stress may be a prerequisite for the expression of important traits in their biocontrol activity, like some cell-wall degrading enzymes20. Unlike to what was found in soil solution medium, Gl006 and Bs006 grew well in the medium containing synthetic root exudates, which contain greater availability of nutrients than the soil solution medium.
In this study, it was found that Bs006-supernatant added to culture medium at 1 and 20%, tend to stimulate the growth of Gl006 during the first 24 h of contact under in vitro conditions (Fig. 1, Supplementary material - Figure S1a). In the same way, exposure of Gl006 conidia to Bs006 supernatant for up to 6 hours, did not affect its antagonistic activity or its viability, suggesting high compatibility among Gl006 with the compounds produced by Bs006 and excreted to the medium, indicating the high potential to use these two biocontrol agents in consortium. In fact, the exchange of metabolites and communication signals among members in a consortium could increase or improve the performance of an artificial consortium21. It was observed that exposing Bs006 to Gl006-supernatant negatively affected the growth of the bacteria (Fig. 2, Supplementary material - Figure S1b), its ability to form biofilms (Fig. 5) and its biocontrol activity (Fig. 8, Table 2), in a concentration dependent manner, although it did not affect its viability and neither its antagonistic ability. This also suggests that Bs006 can achieve the functions of biocontrol in the rhizosphere in Gl006 presence, considering that the concentration of antimicrobial compounds of Gl006 or Trichoderma spp. are much lower in natural conditions than those evaluated in vitro in the present work.
The success of biopesticides based on microorganisms depends largely on the ability to actively colonize the target site by the introduced microorganisms22. Conidia germination is among the critical features in the process of colonization and survival of a fungus in the rhizosphere, where the production of antibiotics and enzymes, like the cell-wall degrading enzymes could favor its competitive ability23,24. It is known also that the formation of biofilms by plant growth promoting rhizobacteria (PGPR) and specifically by B. velezensis, is associated with root colonization. PGPR form micro-colonies or biofilms in sites where there is high concentration of root exudates. In these biofilms, bacteria communicate to each other to perform in a coordinated way and has been related to the induction of resistance25. In this work we found that the contact of Gl006 conidia with Bs006 did not affect the conidial germination, so this direct interaction would not affect the establishment of the fungus in soil. Likewise, high concentrations of Gl006 supernatant were required to affect to Bs006, which is considered a positive result, since concentration of secreted compounds by Trichoderma spp. to the environment in natural conditions, probably is much lower than in the concentration in the supernatant used here which is a dilution (0.25% or 1 × 106 conidia mL−1) of the stock inoculum harvested from solid state fermentation (4 × 108 conidia mL−1).
Triveni, Prasanna, & Saxena26 showed that the mycelium of Trichoderma viride can behave like a matrix used by bacteria P. fluorescens, Azotobacter chrococcum and B. subtilis, to adhere and form biofilms on the fungus surface. The microscopic observations made in the present work allowed to observe this same phenomenon, in which the spores of Bs006 were adhered to conidia of Gl006 after three days contact. It has been described how fungi can contribute to the migration of bacteria as helpers, stimulating their adherence and movement in the soil, as in the case of Burkholderia terrae forming biofilms on the hyphae of Lyophyllum sp.27. This relationship is beneficial for bacteria, considering that movement of bacteria in the soil can be restricted and thus they can reach new microhabitats due to the presence of air among the soil particles28. Moreover, it has been found that during biofilm formation by bacteria on fungi structures, bacteria obtain nutrients from exudates secreted by the fungus or released content after inducing lysis of fungus cells or both microorganisms could have a synergistic action in which they need to break complex substrates in the soil26. In the present study we found that the interaction between Gl006 and Bs006 was not negative for the fungus, considering that the adherence of the bacteria did not affect Gl006 conidia germination and growth, and physical damage was not observed.
Considering that the concentration of antimicrobial metabolites of B. velezensis strains in the rhizosphere is low, its direct activity in the suppression of soil-borne phytopathogens is doubtful29. However, in the present work neither the application of Bs006-supernatant nor the suspension of Bs006-cells on its supernatant were effective against Fusarium wilt, despite the application of higher concentrations of antimicrobial metabolites than those that probably exist in the rhizosphere. Suggesting there are other factors in the rhizosphere or in the plant that influence the antagonistic function of these metabolites synthesized by Bs006 or that these metabolites influence in some manner the host response or affect the native microflora in favor of Fusarium colonization and infection2.
Based on our results, the synergy between Gl006 and Bs006 to control Fusarium wilt could be attributed to the direct action of metabolites contained in Bs006-supernatant on Foph, which could act as a complementary mechanism of action by Gl006. In supernatants and cells interaction experiment it was observed that by eliminating Bs006 cells from the consortium, that is, combining Bs006-supernatant with Gl006-conidia, a high efficiency to control the disease severity was obtained (72%). It has been found that the supernatant of fermentation of Bs006 in LB broth for 48 hours, contained homologous compounds of the cyclic lipopeptides fengycins, iturins and surfactins2, and high production of proteases and cellulases, Izquierdo-García and Moreno-Velandia (unpublished). However, these compounds present in the broth are not enough to completely control Fusarium wilt in cape gooseberry considering that in the present work the application of only supernatant treatment reduced the disease incidence by 34% and the severity by 2%. Furthermore, Gl006-conidia reduced Fusarium wilt incidence by 60% and severity by 55%, activity that could be associated mainly with competition for nutrients and space, in addition to the direct action of cell wall degrading enzymes (CWDE) against Foph. On the other hand, Bs006-cells reduced the disease incidence by 55% and the severity by 44%, which allowed to see that, Gl006 cells and Bs006 cells showed high efficacy both individually and as a mixture (66% on both incidence and severity). However, the synergistic effect to control the disease was not attributed to the presence of Bs006-cells in consortium, it could be attributed to direct interaction between Bs006-supernatant with Gl006-conidia, probably due to direct mechanisms of action in a complementary way against Foph.
Trichoderma spp. and Bacillus subtilis/amyloliquefaciens complex are within the most studied microorganisms in biological control of phytopathogens. The ecological functions of these microorganisms besides biocontrol, such as biodegradation, mineralization, plant growth promotion, stablish the balance in soil for conventional and organic agriculture by increasing the productivity in an agroecosystem30. Both microorganisms are highly resistant to adverse environmental conditions, Trichoderma spp. have long been known to be a soil-borne fungus, and they are certainly common in the soil31, can colonize the surface and inside the plant as endophytes31,32, is ubiquitous and highly diverse, Trichoderma spp. are the culturable fungi most prevalent in soil studies in vitro32. Moreover, Trichoderma spp. can act as a biofertilizer solubilizing mineral nutrients33, has a relatively faster metabolic rates, and secrete anti-microbial secondary metabolites34. Trichoderma spp. and Bacillus spp. also have been studied for their capability in remediation and amelioration of pollutants in the environment35,36. Bacillus velezensis/amyloliquefaciens complex are highly known as good producers of cyclic lipopeptides (CLPs) that display multiple versatile functions in the interactions with co-existing organisms, including bacteria, fungi, oomycetes, protozoan predators and plants, and contribute greatly to its biological control activity in the colonization process and in the direct control against plant pathogens37,38. These characteristics make them ideal candidates to be employed as a consortium in an agroecosystem to control Fusarium wilt disease, but they can also offer other services that can be used in the agroecosystems.
About mechanisms of action of Gl006 and Bs006, we have observed that Gl006 synthesizes CWDE such as chitinases (N-acetyl-glucosaminidase and chitobiosidase), proteases, and β,1-3,glucanase, while Bs006 synthesizes cyclic lipopeptides, chitobiosidase, proteases, and β,1-3,glucanase in presence of Foph and individually as well as in consortium. When Gl006 and Bs006 were evaluated in culture media with natural and artificial root exudates, the activity of each one was maintained but we don’t have found an additive or synergistic effect. On the other hand, we evaluated the ability for induce resistance in cape gooseberry under a split root model and found similar results in control of the disease, Izquierdo-García and Moreno-Velandia (unpublished).
The success of a microbial consortium depends on the individual ability of BCA and their mutual interaction21. However, it has been found that the biocontrol effect in a consortium can be due to a predominant microorganism in the mixture39. In this study, it was found that neither of the two microorganisms negatively affected the other under nutritional conditions similar to those in the rhizosphere (Fig. 7).
Adhesion of B. velezensis spores to the surface of T. virens conidia without causing damage to the fungus, suggests a compatible interaction between both biocontrol agents. Supernatants from liquid culture of B. velezensis could stimulate the biocontrol activity of T. virens, while supernatant of T. virens reduced B. velezensis growth. However, considering that concentrations of metabolites synthesized by these microorganisms in soil could be lower than those synthesized in culture medium, our results suggest that metabolites that are present in supernatant do not affect its biocontrol performance in vivo. We found that biocontrol agents T. virens Gl006 and B. velezensis Bs006 grew in a compatible way on artificial exudates, cape gooseberry root exudates and on the surface of cape gooseberry seeds and roots, suggesting that they could be used in consortium as a biocontrol treatment in cape gooseberry plants.
T. virens Gl006 and B. velezensis Bs006 are two native soil inhabitants microorganisms and their compatibility as a consortium has been demonstrated in this study, where it is important to maintain the concentrations of both cells and supernatants in favor of their compatibility. Additional studies are required to know molecular interactions of these microorganisms for a better understanding of their compatibility in the rhizosphere.
Materials and methods
Microorganisms and culture conditions
F. oxysporum f. sp. physali strain Map5 (hereafter referred as Foph) and biological control agents (BCA) Trichoderma virens (Gl006) and Bacillus velezensis (Bs006) were stored at −80 °C in glycerol (30%) and peptone (0,1%) sterile solution at the Microorganisms Collection of Corporación Colombiana de Investigación Agropecuaria - AGROSAVIA. Inoculum of Foph was produced following the procedure described by Moreno2. Briefly, this fungus was grown for 7 days on sterile potato-dextrose broth (PDB, Difco®) with an initial concentration of 1 × 106 microconidia.mL−1, under continuous agitation (125 rpm) at 25 °C. After this, microconidia produced were harvested. The concentration of microconidia suspension required for the experiments was adjusted by Neubauer chamber to 1 × 106 microconidia.mL−1, with sterile distilled water (SDW) for in vitro tests and with tap water for greenhouse tests. Gl006 was reactivated on potato-dextrose-agar culture medium (PDA) and conidia from the second seven-day-old subculture were harvested by scraping with SDW to obtain the inoculum. Bs006 was reactivated in Luria Bertani Agar culture medium (LBA: Tryptone 10 g, NaCl 10 g, yeast extract 5 g and agar 18 g/L). Pre-inoculum was prepared from the second 24 h-old subculture to start the fermentation in LB broth in an initial concentration of 5 × 106 cfu.mL−1. Liquid culture of Bs006 was maintained in continuous agitation in orbital shaker at 125 rpm and 30 °C for 48 h. The required concentration of the bacterial suspensions was adjusted by measuring the optical density in spectrophotometer (BIOTEK®) (OD600 nm = 0.5 ~ 2.49 × 108 cfu.mL−1).
Plant material
Commercial cape gooseberry seeds (Semicol®)) were used. Seeds were surface disinfected before sowing by dipping in NaOCl solution (3%) during 20 min and then three washes with sterile distilled water. For greenhouse trials 60-day-old seedlings were used, grown in germination trays of 72 alveoli with autoclaved peat (two continuous cycles of 20 min, 15 PSI and 121 °C). Each seedling was transplanted to 600 g of soil substrate: rice husk (3:1 v/v) in black polypropylene bags of 1000 cm3.
In vitro evaluations of the effect of supernatants from liquid cultures of Bs006 and Gl006
The potential effects of secondary metabolites secreted in liquid culture on growth, antagonistic activity, conidia germination for Gl006 and biofilms formation for Bs006 were determined under in vitro conditions, after exposure of each microorganism to different concentrations of its partner’s supernatant.
Production of supernatants from biocontrol agents
To obtain the supernatant of Gl006, the fungus was grown in Erlenmeyer (300 mL) with 150 mL of sterile PDB (Difco®) for five days (125 rpm, 25 °C) with an initial concentration of 1 × 105 conidia.mL−1. After incubation, the fermentation broth was harvested filtering by Whatman No. 1 paper and subsequently by sterile 0.22 μm filters (Millipore®) and stored at −20 °C until its use. To obtain the supernatant of Bs006, bacteria was grown in sterile LB broth for 48 hours and then was centrifuged. Supernatant was harvested and filtered with sterile 0.22 μm filters (Millipore®). Biomass obtained from Bs006 culture was washed twice to eliminate waste of supernatant and resuspended in SDW, homogenizing by shaking in vortex at maximum speed for two minutes, to be used as inoculum in these experiments. On the other hand, conidia of Gl006 were harvested from a seven-day-old culture on PDA, flooding the medium with sterile distilled water and scraping with a sterile metal rake. The suspension obtained was shaken in a vortex at maximum speed for 30 seconds and then was filtered with sterile muslin (pore size 0,5 mm) to eliminate pieces of mycelium.
Effect of supernatants on the growth of biological control agents
To evaluate the effect of Gl006 supernatant on Bs006 and vice versa (Bs006 supernatant on Gl006) the microorganisms were grown in LB or PDB broth, respectively, supplemented with 1, 5, 10, 20 and 30% of supernatant of the other microorganism in 96-well microplates (Greiner®). The initial concentrations of Bs006 and Gl006 were adjusted to 5 × 106 cfu.mL−1 and 1 × 105 conidia.mL−1, respectively. Liquid media supplemented with the supernatants without microbial inoculum were used as blanks, and liquid media supplemented with sterile PDB or LB medium and inoculated with the biocontrol agents were used as controls. Inoculated microplates were incubated for 48 h at 28 °C for Gl006 and 30°C for Bs006 in continuous agitation. The optical density (OD) at wavelength of 630 nm was measured every hour using a microplate reader (BIOTEK®) managed with Gen5® software. Microscope observations were made to evaluate possible effects on the morphology of microorganisms.
Effect of B. velezensis Bs006 supernatant on conidia germination of T. virens Gl006
Conidia of Gl006 [5 × 106 conidia.mL−1] were exposed during one or six hours to solution of Bs006 supernatant at concentrations mentioned above prepared in sterile solution of Tween 80 (0.1%). After exposition, Gl006 conidia were separated from the solution by centrifugation (14000 rpm, 5 min), then washed twice and resuspended in sterile distilled water. Nine microdrops-10 μL each were inoculated on sterile water-agar in Petri dishes and incubated for 24 h. The proportion of germinated conidia was recorded under 40X magnification in an optical microscope (Olympus® CX41).
Effect of Gl006 conidia and supernatant on Bs006 biofilm formation
To determine the effect of T. virens conidia and supernatant on Bs006 biofilm formation, we followed the procedure described by Peeters, Nelis, & Coenye (2008)40 and O’Toole (2011)41. Briefly, the minimum standard medium M63 for biofilm formation supplemented with magnesium sulfate (Merck®), glucose (Sigma®) and casamino acids (Difco®) was used (O’Toole, 2011). This medium was supplemented with Gl006-supernatant at concentrations of 1, 5, 10, 20 and 30%. Moreover, the effect of co-culture of Gl006 in concentrations of 105, 106 and 107 conidia.mL−1 with Bs006 cells (24 h-old in LBA) at 1 × 108 cfu.mL−1 was evaluated. In both cases, 100 μL of suspension were added per well in a 96-well microplate (Greiner®) and incubated for 24 hours at 30 °C. Nine wells were served for each treatment. The fermented broth and the cells in suspension were eliminated after the incubation, and a rinsing was done by adding 100 μL of NaCl solution (0.85%) per well. Then, 100 μL of methanol (99%) were added incubating for 15 minutes, then was eliminated, and the plate was left to dry in laminar flow chamber. Subsequently 200 μL of violet crystal (for gram staining) were added and incubated for 20 minutes, then the colorant was removed and washed with tap water. 150 μL of acetic acid solution (33%) were added to dilute the adhered dye. Absorbance at 590 nm was recorded using a microplate reader (BIOTEK®) and the Gen5® software. The experiment was repeated once.
Effect of supernatants on the antagonistic activity of BCA on solid media
Conidia of T. virens Gl006 and vegetative cells of B. velezensis Bs006 exposed for one and six hours to the reciprocal supernatant solutions prepared in Tween 80 (0.1%) sterile solution as described above, were used to carry out a dual confrontation test with Foph in Petri dishes (90 mm) with PDA medium. The inoculated dishes were incubated for seven days at 28 °C in darkness and then the diameter of Foph colony was measured with a caliper (Fischer®). The experiment had five dishes per treatment and was performed twice.
In vitro evaluations of co-cultures of biocontrol agents and the influence of the culture medium on the antagonistic activity
The effect of the culture medium and the combination of Bs006 with Gl006 on its antagonistic activity against Foph were measured by dual confrontation tests. The culture media used were LBA, PDA, PD-LBA (50% each), artificial root exudates (ARE)37, root exudates from cape gooseberry (CRE), soil solution (SS), ARE and CRE media prepared on SS (SARE and SCRE, respectively).
Root exudates were obtained from 30 days-old cape gooseberry plants. Plants were grown on peat in rooting trays and 30 days after sowing were uprooted from the substrate and washed with tap water. Subsequently, the aerial part of the plant (stem and leaves) and roots were immersed in NaOCl solution (2%) during 0.2 and 5 min, respectively and washed four consecutive times with sterile distilled water. Disinfected plants were placed in sterile plastic tubes 50 mL (Falcon®) with roots immersed in 40 mL of sterile soil solution (filtered by 0.2 μm) and were kept at room temperature (18 °C on average) during five days with photoperiod (12 h artificial light − 12 h dark). Rooting solution was harvested and filtered by 0.2 μm filters and stored at −20 °C until use.
Soil solution was obtained by mixing 500 g of substrate (soil:rice husk 3:1 v/v) with 1000 mL of tap water. This solution was agitated on a rotary shaker at 125 rpm during one hour at room temperature and then was filtered with muslin cloth (0.5 mm pore size), then it was passed through 0.8 μm and 0.22 μm sterile filters.
Suspensions of Bs006 vegetative cells (24 h-old on LBA) and Gl006 conidia (7 days-old on PDA) were prepared at 1 × 108 cfu.mL−1 and 1 × 106 conidia.mL−1, respectively. Two 10 μL-drops were inoculated at opposite points, 1.5 cm from the edge of the Petri dish. Simultaneously, Foph (10 μL, 1 × 105 microconidia.mL−1) was inoculated in the center of the plate. Another set of Petri dishes with culture media were inoculated only with one 10 μL-drop of a joint suspension of Bs006, Gl006 and Foph adjusted to concentrations described above. The inoculated plates were incubated for seven days at 28 °C in darkness and then the diameter of the Foph colony was measured with a caliper (Fischer®).
Microscopy observations
Micro-culture of BCA
For microscopic observations of the interacting cells of T. virens Gl006 and B. velezensis Bs006, a solid support consisting of root exudates of cape gooseberry and sterile agar (18 g.L−1) mixture (CREA) was inoculated with 2 μL-drops of cell suspension from Bs006 48 h-old (spores mainly) and Gl006 7 day-old (conidia) on opposite sides on jellified squares-1,5 cm2. The jellified support was incubated in a moistened chamber for 72 hours. After this time, observations were made on an optical microscope (Olympus® CX41) and fluorescence microscope (Olympus® IX71).
Combined inoculations of Bs006 and Gl006 in planta
Disinfected cape gooseberry seeds were germinated on jellified nutrient medium (GNM)37 incubated in the dark for 15 days. Germinated seeds were inoculated at the crown level with 2 μL of mixture or individual inoculum of T. virens Gl006 and B. velezensis Bs006 [3 × 105 conidia.mL−1 and 3 × 107 cfu.mL−1, respectively]. Additional inoculations were also carried out on disinfected seeds. Inoculated seeds and seedlings were maintained on GNM at 25 °C for 48 hours. Samples of 1 cm in length were taken from germinated seeds, including the point of inoculation, non-germinated seeds were also observed by scanning electron microscope (FEI®, QUANTA FEG 650) with Field Emission technology. The samples were processed by environmental microscopy of plant tissue at 2–4 °C, 400 Pa of pressure and 49.2 to 56% RH.
Evaluation of the biocontrol activity of Bs006 and Gl006 under greenhouse conditions
The effect of T. virens Gl006 with B. velezensis Bs006 consortium, including combinations of biomass-biomass, supernatant-biomass, and supernatant-supernatant (Table 2) against Fusarium wilt of cape gooseberry was evaluated under greenhouse conditions. The wet soil was artificially inoculated with microconidia suspension of Foph to obtain a concentration of 1 × 104 cfu.g−1 of soil. Then, cape gooseberry seedlings (60 day-old) were transplanted. Gl006 was adjusted to 1 × 106 conidia.mL−1 and Bs006 to 1 × 108 cfu.mL−1. Biocontrol treatments were applied to the soil immediately after transplant (30 mL.plant−1). Plants were kept for 6 weeks under greenhouse with an average temperature of 25 °C.
For this, Bs006 was grown in LB broth for 48 hours at 28 °C and 125 rpm while Gl006 was grown on PDA for seven days at 28 °C. The fermented broth of Bs006 was centrifuged (15000 rpm, 15 min, 4 °C) to separate biomass from supernatant. Supernatant was harvested and filtered by 0.22 μm filters (Sartorius®) and biomass of Bs006 was washed twice with sterile distilled water to remove supernatant residues. Gl006 conidia were harvested by scraping the Petri dish and to obtain the supernatant of this fungus, PDA medium in which the fungus grew was liquefied in ULTRA TURRAX® at maximum speed for two minutes, adding 15 mL of sterile distilled water per Petri dish. The suspension obtained was centrifuged under the conditions described before and the obtained supernatant was filtered by 0.22 μm. For the preparation of the treatments, biomass of each biological control agent was resuspended in a supernatant solution of the reciprocal microorganism (biomass of Bs006 in Gl006 supernatant solution and vice versa). Bs006-supernatant and Gl006-supernatant solutions prepared in tap water at 8.3 and 0.25%, respectively were used to dilute the microorganisms according to the volume of the original suspension required to adjust the biomass concentration for each case to 108 cfu.mL−1 for Bs006 and 106 conidia.mL−1 for Gl006. Combinations of supernatant-supernatant and conidia-bacteria free of supernatant were included as additional treatments. The application of the treatments was performed immediately after transplant.
An experimental design of randomized complete blocks with four replicates was used. The size of the experimental unit was ten plants. Incidence, expressed as the percentage of diseased plants of the total number of plants in the experimental unit and severity of the disease were measured using the scale described by Moreno2 (Supplementary material - Table S1, Figure S5) and presented as the disease intensity index, calculated with the equation \({DII}=(\sum {Si}\ast {Ni})\div(5\ast N)\), where Si is the level of severity of the symptoms, Ni is the number of plants in each severity level Si, and N is the total number of plants in the experimental unit42. Disease recording was made every four days from the expression of the first symptoms and the area under the disease progress curve (AUDPC) was calculated. The efficacy of disease reduction was determined based on the AUDPC values with the equation E=((a − b) ÷ a) * 100, where a represents the value of the disease in the negative control and b represents the disease in a treatment43.
To determine whether the efficacy on the disease incidence and/or severity showed by the BCA mixtures was the result of a synergistic, antagonistic or additive effect, the expected efficacy (Eesp) in consortium was calculated using the equations of Abbott43 Eesp = (a + b) − ((a × b)/100) where, a represents the efficacy by one component of the consortium and b represents the efficacy by the other component. Then the synergy factor (SF) was calculated using the equation \({SF}={Eobs}/{Eesp}\) where, Eobs corresponds to the efficacy showed by the consortium44.
Statistical analysis
Data were submitted to normality tests according to Shapiro Wilk (P > 0.05) and variance homogeneity with Levene test. Significance was determined with ANOVA analysis in a general linear model (GLM). Duncan multiple range test and Tukey test were used to compare media between treatments. Data were analyzed using the statistical software S.A.S. (9.4: SAS Institute, Cary, NC).
References
Dean, R. et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 13, 414–430, https://doi.org/10.1111/j.1364-3703.2011.00783.x (2012).
Moreno-Velandia, C. M., Izquierdo-García., L. F., Ongena, M., Kloepper, J. W. & Cotes, A. M. Soil sterilization, pathogen and antagonist concentration affect biological control of Fusarium wilt of cape gooseberry by Bacillus velezensis Bs006. Plant Soil 435, 39–55, https://doi.org/10.1007/s11104-018-3866-4 (2019).
Raupach, G. S. & Kloepper, J. W. Mixtures of plant growth-promoting rhizobacteria enhance biological control of multiple cucumber pathogens. Phytopathology 88, 1158–1164, https://doi.org/10.1094/PHYTO.1998.88.11.1158 (1998).
Paerl, H. W. & Pinckney, J. L. A mini-review of microbial consortia: Their roles in aquatic production and biogeochemical cycling. Microb. Ecol. 31, 225–247, https://doi.org/10.1007/BF00171569 (1996).
Zhang, S., Merino, N., Okamoto, A. & Gedalanga, P. Interkingdom microbial consortia mechanisms to guide biotechnological applications. Microb. Biotechnol. 11, 833–847, https://doi.org/10.1111/1751-7915.13300 (2018).
Xu, X.-M., Jeffries, P., Pautasso, M. & Jeger, M. J. Combined use of biocontrol agents to manage plant diseases in theory and practice. Phytopathology 101, 1024–1031, https://doi.org/10.1094/PHYTO-08-10-0216 (2011).
Sarma, B. K., Yadav, S. K., Singh, S. & Singh, H. B. Microbial consortium-mediated plant defense against phytopathogens: Readdressing for enhancing efficacy. Soil Biol. Biochem. 87, 25–33, https://doi.org/10.1016/j.soilbio.2015.04.001 (2015).
Estevez de Jensen, C., Percich, J. A. & Graham, P. H. Integrated management strategies of bean root rot with Bacillus subtilis and Rhizobium in Minnesota. F. Crop. Res. 74, 107–115, https://doi.org/10.1016/S0378-4290(01)00200-3 (2002).
Estberg, M. et al. Microbial inoculation for improving the growth and health of micropropagated strawberry. App. Soil Ecol. 27, 243–258, https://doi.org/10.1016/j.apsoil.2004.05.006 (2004).
Jain, A., Singh, A., Singh, S. & Singh, H. B. Biological management of Sclerotinia sclerotiorum in pea using plant growth promoting microbial consortium. J. Basic Microbiol. 55, 961–972, https://doi.org/10.1002/jobm.201400628 (2015).
Sundaramoorthy, S., Raguchander, T., Ragupathi, N. & Samiyappan, R. Combinatorial effect of endophytic and plant growth promoting rhizobacteria against wilt disease of Capsicum annum L. caused by Fusarium solani. Biol. Control 60, 59–67, https://doi.org/10.1016/j.biocontrol.2011.10.002 (2011).
Marimuthu, S., Ramamoorthy, V., Samiyappan, R. & Subbian, P. Intercropping system with combined application of Azospirillum and Pseudomonas fluorescens reduces root rot incidence caused by Rhizoctonia bataticola and increases seed cotton yield. J. Phytopathol. 161, 405–411, https://doi.org/10.1111/jph.12084 (2013).
Alizadeh, H. et al. Induced systemic resistance in cucumber and Arabidopsis thaliana by the combination of Trichoderma harzianum Tr6 and Pseudomonas sp. Ps14. Biol. Control 65, 14–23, https://doi.org/10.1016/j.biocontrol.2013.01.009 (2013).
McGovern, R. J. Management of tomato diseases caused by Fusarium oxysporum. Crop Prot. 73, 78–92, https://doi.org/10.1016/j.cropro.2015.02.021 (2015).
Yobo, K. S. Biological control and plant gowth promotion by selected Trichoderma and Bacillus species. Doctoral thesis. KwaZulu-Natal University. Pietermaritzburg. 205 p. (2015).
Wu, Q. et al. Co - culture of Bacillus amyloliquefaciens ACCC11060 and Trichoderma asperellum GDFS1009 enhanced pathogen - inhibition and amino acid yield. Microbial Cell Factories. 17, 1–12, https://doi.org/10.1186/s12934-018-1004-x (2018).
Karuppiah, V., Sun, J., Li, T., Vallikkannu, M. & Chen, J. Co-cultivation of Trichoderma asperellum GDFS1009 and Bacillus amyloliquefaciens 1841 causes differential gene expression and improvement in the wheat growth and biocontrol activity. Frontiers in microbiology. 10, 1–16, https://doi.org/10.3389/fmicb.2019.01068 (2019).
Silva, A. D. Acción in vitro de Trichoderma spp. y Bacillus spp. como controladores biológicos conjuntos contra Fusarium oxysporum en uvilla (Physalis peruviana), ecotipo colombiano, en la sierra norte y centro del Ecuador. Microbiology degree thesis. Universidad Católica de Ecuador. Quito. 112 p (2018).
Weller, D. The rhizosphere with Bacteria. Ann. Rev. Phytopathol. 26, 379–407, https://doi.org/10.1038/nrmicro1129 (1988).
Hjeljord, L. G., Stensvand, A. & Tronsmo, A. Antagonism of nutrient-activated conidia of Trichoderma harzianum (atroviride) P1 against Botrytis cinerea. Phytopathology 91, 1172–1180, https://doi.org/10.1094/PHYTO.2001.91.12.1172 (2001).
Bhatia, S. K. et al. Biotechnological potential of microbial consortia and future perspectives. Crit. Rev. Biotechnol. 38, 1209–1229, https://doi.org/10.1080/07388551.2018.1471445 (2018).
Polyanskaya, L. M. et al. Autoregulation of conidium germination in micromycetes of the genus Trichoderma. Microbiology 73, 79–83, https://doi.org/10.1023/B:MICI.0000016373.96469.5a (2004).
Mitchell, C. P. & Dix, N. J. Growth and germination of Trichoderma spp. under the influence of soil fungistasis. Trans. Br. Mycol. Soc. 64, 235–241, https://doi.org/10.1016/S0007-1536(75)80108-2 (1975).
Harman, G. E. Myths and dogmans of biocontrol: changes in perceptions derived from research on Trichoderma harzianum T-22. Plant Dis. 84, 377–393, https://doi.org/10.1094/PDIS.2000.84.4.377 (2000).
Choudhary, D. K. & Johri, B. N. Interactions of Bacillus spp. and plants - With special reference to induced systemic resistance (ISR). Microbiol. Res. 164, 493–513, https://doi.org/10.1016/j.micres.2008.08.007 (2009).
Triveni, S., Prasanna, R. & Saxena, A. K. Optimization of conditions for in vitro development of Trichoderma viride-based biofilms as potential inoculants. Folia Microbiol. 57, 431–437, https://doi.org/10.1007/s12223-012-0154-1 (2012).
Warmink, J. A., Nazir, R., Corten, B. & van Elsas., J. D. Hitchhikers on the fungal highway: The helper effect for bacterial migration via fungal hyphae. Soil Biol. Biochem. 43, 760–765, https://doi.org/10.1016/j.soilbio.2010.12.009 (2011).
Warmink, J. A. & Van Elsas, J. D. Migratory response of soil bacteria to Lyophyllum sp. strain Karsten in soil microcosms. Appl. Environ. Microbiol. 75, 2820–2830, https://doi.org/10.1128/AEM.02110-08 (2009).
Chowdhury, S. P., Hartmann, A., Gao, X. W. & Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42 - A review. Front. Microbiol. 6, 1–11, https://doi.org/10.3389/fmicb.2015.00780 (2015).
Kostov, O., Vasileva, V. & Kaloianova, N. Industrial, ecological and agricultural importance of biocontrol agents based on Trichoderma species. Ecol. Futur. 8, 28–37 (2009).
Samuels, G. J. Trichoderma: Systematics, the Sexual State, and Ecology. Pyhtopathology. 96, 195-206, https://doi.org/10.1094/PHYTO-96-0195 (2006).
Lazarovits, G., Turnbull, A. & Johnston-Monje, D. Plant Health Management: Biological Control of Plant Pathogens. Encyclopedia of Agriculture and Food Systems Elsevier Ltd., 4, https://doi.org/10.1016/B978-0-444-52512-3.00177-7 (2014).
Harman, G. E., Howell, C. R., Viterbo, A., Chet, I. & Lorito, M. Trichoderma species - Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2, 43–56, https://doi.org/10.1038/nrmicro797 (2004).
Keswani, C., Singh, S. P. & Singh, H. B. A superstar in biocontrol enterprise: Trichoderma spp. Biotech Today. 3, 27, https://doi.org/10.5958/2322-0996.2014.00005.2 (2013).
Kareem, S. O., Adegoke, O. O., Balogun, S. A., Afolabi, A. T. & Akinde, S. B. Degradation of premium spirit (PMS) by lipase from Bacillus thuringiensis and Lysinibacillus sphaericus. Nigerian Journal of Biotechnology. 33, 34–40, https://doi.org/10.4314/njb.v33i1.5 (2017).
Harman, G. E., Lorito, M. & Lynch, J. M. Uses of Trichoderma spp. to alleviate or remediate soil and water pollution. Adv. Appl. Microbiol. 56, 313–30, https://doi.org/10.1016/s0065-2164(04)56010-0 (2004).
Nihorimbere, V. et al. Impact of rhizosphere factors on cyclic lipopeptide signature from the plant beneficial strain Bacillus amyloliquefaciens S499. FEMS Microbiol. Ecol. 79, 176–191, https://doi.org/10.1111/j.1574-6941.2011.01208.x (2012).
Ongena, M. & Jacques, P. Bacillus lipopeptides: versatile weapons for plant disease biocontrol. Trends Microbiol. 16, 115–125, https://doi.org/10.1016/j.tim.2007.12.009 (2008).
Denoth, M., Frid, L. & Myers, J. H. Multiple agents in biological control: Improving the odds? Biol. Control 24, 20–30, https://doi.org/10.1016/S1049-9644(02)00002-6 (2002).
Peeters, E., Nelis, H. J. & Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 72, 157–165, https://doi.org/10.1016/j.mimet.2007.11.010 (2008).
O’Toole, G. A. Microtiter Dish Biofilm Formation Assay. J. Vis. Exp. 10–11, 10.3791/2437 (2011).
Hervás, A., Landa, B., Datnoff, L. E. & Jiménez-Díaz, R. M. Effects of commercial and indigenous microorganisms on Fusarium wilt development in chickpea. Biol. Control 13, 166–176, https://doi.org/10.1006/bcon.1998.0659 (1998).
Abbott, W. S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 18, 265–267, https://doi.org/10.1093/jee/18.2.265a (1925).
Levy, Y., Benderly, M., Cohen, Y., Gisi, U. & Bassand, D. The joint action of fungicides in mixtures: comparison of two methods for synergy calculation. EPPO Bull 16, 651–657, https://doi.org/10.1111/j.1365-2338.1986.tb00338.x (1986).
Acknowledgements
This work was carried out with the financial support from Ministerio de Agricultura y Desarrollo Rural – MADR through Tv15 and Tv16 transference resources to Corporación Colombiana de Investigación Agropecuaria - AGROSAVIA, Project 687. The authors acknowledge Carlos Chacón and Humberto Ibarra by technical support at microscopy laboratories from Universidad Industrial de Santander – UIS – and Universidad de los Andes, respectively. Also, thanks to Dr. Andrea Paola Zuluaga for the critical review of the manuscript.
Author information
Authors and Affiliations
Contributions
C.A.M.V. and L.F.I.G. conceived the research idea. L.F.I.G. carried out the experiments and drafted the manuscript under the advice of C.A.M.V. L.F.I.G., A.G.A., A.M.C. and C.A.M.V. contributed to analysis and interpretation of data, as well as critical revising and approval of the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Izquierdo-García, L.F., González-Almario, A., Cotes, A.M. et al. Trichoderma virens Gl006 and Bacillus velezensis Bs006: a compatible interaction controlling Fusarium wilt of cape gooseberry. Sci Rep 10, 6857 (2020). https://doi.org/10.1038/s41598-020-63689-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-63689-y
- Springer Nature Limited
This article is cited by
-
Bacterial consortium based on Pseudomonas fluorescens, Lysinibacillus xylanilyticus and Bacillus velezensis reduces clubroot disease in broccoli
European Journal of Plant Pathology (2024)
-
The fungal root endophyte Serendipita vermifera displays inter-kingdom synergistic beneficial effects with the microbiota in Arabidopsis thaliana and barley
The ISME Journal (2022)
-
Bacterial-fungal interactions under agricultural settings: from physical to chemical interactions
Stress Biology (2022)
-
Characterization of the belowground microbial community and co-occurrence networks of tobacco plants infected with bacterial wilt disease
World Journal of Microbiology and Biotechnology (2022)
-
Screening for effective microbial consortia against Fusarium wilt of cape gooseberry (Physalis peruviana)
BioControl (2021)