
Abstract
Preclinical studies suggested that IgG2c isotype may specifically impair skeletal muscle insulin sensitivity in mice. In this study we investigated the association between serum levels of the four IgG subclasses and insulin sensitivity in non-diabetic individuals. Total IgG, IgG1, IgG2, IgG3 and IgG4 levels were measured in 262 subjects. Whole-body insulin sensitivity was assessed by euglycemic hyperinsulinemic clamp. IgG2 levels were positively correlated with BMI, waist circumference, 2-h post-load glucose levels and complement C3. Serum IgG2, but not IgG1, IgG3 and IgG4 levels were negatively correlated with whole-body insulin sensitivity (r = −0.17; P = 0.003) and muscle insulin sensitivity index (r = −0.16; P = 0.03) after adjustment for age and gender. No significant correlation was found between IgG2 levels and hepatic insulin resistance assessed by HOMA-IR and liver IR index. In a multivariable regression analysis including variables known to affect insulin sensitivity such as age, gender, BMI, smoking, lipids, inflammatory markers, fasting and 2-h post-load glucose levels, IgG2 levels were independently associated with insulin-stimulated glucose disposal (β = −0.115, 95% CI: −0.541 to −0.024; P = 0.03). These data demonstrate the independent association between higher levels of IgG2 and decreased whole-body insulin sensitivity, thus confirming in humans the animal-based evidence indicating the pathogenic role of IgG2 in insulin resistance.
Similar content being viewed by others


Introduction
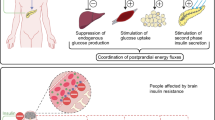
Observational studies have solidified the concept that low-grade inflammation plays a pathophysiological role in insulin resistance associated with type 2 diabetes mellitus1,2,3,4,5,6,7. Markers of inflammation have been extensively studied in these metabolic conditions, with convincing evidence showing that elevated concentrations of C reactive protein, fibrinogen, complement C3, and white blood cells count are predictors of type 2 diabetes. In addition to these reliable markers of inflammation, widely used in clinical practice, also immunoglobulin G (IgG) level, an indicator of adaptive immune system activation, has been associated with obesity and type 2 diabetes in cross-sectional and prospective studies8,9,10. Experimental studies in mice have suggested a role of B cells in the pathogenesis of insulin resistance associated with diet-induced obesity11,12. In particular, it has been shown that transfer of proinflammatory IgG2c isotype purified from sera of diet-induced obese mice caused insulin resistance and glucose intolerance in the recipient rodents11. More recently, it has been reported that IgG2c isotype, but not IgG1 or IgG2b, selectively caused skeletal muscle insulin resistance in diet-induced obese mice by activating the IgG receptor FcγRIIB in endothelium12. The activation of FcγRIIB by IgG2c isotype from diet-induced obese mice, in turn, impaired endothelial cell insulin transcytosis, resulting in reduced insulin delivery to skeletal muscle and impairment in muscle glucose disposal12. Furthermore, IgG isolated from individuals with type 2 diabetes caused glucose intolerance and insulin resistance in IgG-deficient mice via FcγRIIB, indicating that similar processes may be effective in humans12. Prompted by the need to better understand the pathophysiological role of human IgG isotypes in glucose homeostasis disorders, we sought to determine their potential role in insulin resistance. To address this issue, we assessed human serum IgG isotypes (IgG1, IgG2, IgG3, and IgG4) in non-diabetic offspring of type 2 diabetic individuals and correlated their concentration with insulin sensitivity assessed using the euglycemic hyperinsulinemic clamp.
Results
Clinical features and biochemical findings of the study group stratified by gender are shown in Table 1. Men were more likely to be current smokers and exhibited a worse cardio-metabolic risk profile including higher waist circumference values, blood pressure, triglycerides and fasting plasma glucose levels, and lower HDL cholesterol, muscle insulin sensitivity index and insulin-stimulated glucose disposal, assessed by euglycemic-hyperinsulinemic clamp. No gender-specific differences were observed in the other indexes of insulin sensitivity either based on fasting measurements such as QUICKI, and homeostasis model assessment insulin resistance (HOMA-IR) or derived by OGTT including liver insulin resistance (liver IR) index, glucose0–30 (AUC) × insulin0–30 (AUC) index, Stumvoll Insulin Sensitivity Index (ISI), Gutt’s ISI0,120, and Matsuda index. Additionally, no gender-specific differences were observed in total IgG or IgG isotypes levels except for IgG4, which were higher in men (Table 1).
Univariate correlations between circulating IgG isotypes levels and anthropometric and metabolic variables are shown in Table 2. After adjusting for age and gender, only IgG2 levels exhibited a significant correlation with anthropometric and metabolic traits with the exception of IgG1, which displayed an inverse relationship with percentage of fat free mass. Particularly, IgG2 levels were positively correlated with BMI, waist circumference, 2-h post-load glucose levels and complement C3. Serum IgG2, but not IgG1, IgG3, and IgG4 levels were negatively correlated with insulin-stimulated glucose disposal, and with the OGTT-derived insulin sensitivity indexes Stumvoll ISI, Matsuda, Gutt’s ISI0,120, and muscle insulin sensitivity index (Table 2). No significant association between IgG2 concentrations and hepatic insulin sensitivity estimated by HOMA-IR, liver IR and glucose0–30 (AUC) × insulin0–30 (AUC) index was observed. The correlation between IgG2 levels and insulin-stimulated glucose disposal remained significant after further adjusting for BMI in addition to gender and age (P = 0.005). By contrast, the correlations between IgG2 levels and 2-h post-load glucose or complement C3 levels were no longer statistically significant after adjustment for BMI. Furthermore, the negative relationship between IgG2 levels and insulin-stimulated glucose disposal was statistically significant in the subgroup of subjects with normal glucose tolerance (n = 200) as well as in the small subgroup of individuals with impaired glucose tolerance and/or impaired fasting glucose (n = 62) (Suppl. Tables 1 and 2).
In order to confirm the specific association between insulin-stimulated glucose disposal and IgG2 levels we subdivided the whole study population in tertiles of insulin-stimulated glucose disposal. After adjusting for age and gender serum IgG2 levels, but not IgG1, IgG3, and IgG4 levels progressively decreased in the intermediate and highest tertiles of insulin-stimulated glucose disposal in comparison to the lowest tertile (Table 3).
Next, we built three models of multivariable regression analysis including variables known to affect insulin sensitivity to evaluate the independent contribution of IgG2 levels to insulin-stimulated glucose disposal (Table 4). Comparison of standardized coefficients allowed the determination of the relative strength of each trait association with insulin-stimulated glucose disposal. We found that IgG2 levels were associated with insulin-stimulated glucose disposal independently of age, gender, and BMI (Table 4, model 1). The association between IgG2 levels and insulin-stimulated glucose disposal remained significant when smoking status, blood pressure, total and HDL cholesterol, triglycerides, fasting and 2-h post-load glucose concentrations were included in the model in addition to age, gender, and BMI (Table 4, model 2). Similar results were obtained when waist circumference replaced BMI in the regression model (Table 4, model 3). In a full adjusted model including hsCRP, white blood cells count, and complement C3 in addition to variables analyzed in the model 2, IgG2 levels were independently associated with insulin-stimulated glucose disposal (β = −0.115, P = 0.03), with the model explaining 32.9% of variation (R2 = 0.324) of insulin-stimulated glucose disposal.
Discussion
Prior cross-sectional and prospective studies have shown that total serum IgG levels, a nonspecific marker of the adaptive immune system activation, are associated with obesity and type 2 diabetes8,9,10. Additionally, animal-based studies have suggested that IgG2c isotype, but not IgG1 or IgG2b, may have a role in causing a selective state of muscle insulin resistance in diet-induced obese mice11,12. These observations provided the rationale to investigate the relationship between serum IgG1, IgG2, IgG3, and IgG4 isotypes and insulin sensitivity in humans. In this cross-sectional study including nondiabetic offspring of type 2 diabetic individuals with a wide range of BMI, we found that in univariate analysis serum concentrations of IgG2, but not IgG1, IgG3, or IgG4, were inversely associated with whole-body insulin-stimulated glucose disposal. This association remained statistically significant in multivariate analysis after adjustments for potential confounders such as indexes of adiposity, lipids levels, measures of glucose tolerance status, and markers of inflammation. Keeping in mind that under the hyperinsulinemic conditions reached during euglycemic hyperinsulinemic clamp studies (with insulin infusion rate fixed 40 mU/m2 per min) hepatic glucose production is suppressed and 80–90% of the infused glucose is taken up by skeletal muscle, insulin-stimulated glucose disposal may be assumed to primarily reflect skeletal muscle insulin sensitivity13. Notably we found that IgG2 levels were negatively correlated with both insulin-stimulated glucose disposal and muscle insulin sensitivity index. Accordingly, IgG2 levels were negatively correlated with surrogate OGTT-derived indexes including Stumvoll ISI, Gutt’s ISI0,120, Matsuda which are strongly correlated with skeletal muscle insulin sensitivity14. By contrast, no significant correlations were found between IgG2 levels and HOMA-IR index, a surrogate index of liver insulin resistance, primarily reflecting hepatic glucose production14, and the liver IR index and glucose0–30 (AUC) × insulin0–30 (AUC) index, which were specifically developed to estimate hepatic insulin resistance14,15. These results are in line with animal-based evidence showing that IgG2c affects muscle rather than hepatic insulin sensitivity12. To the best of our knowledge, this is the first study in humans assessing the relationship between serum IgG isotypes levels and whole-body insulin sensitivity evaluated using the euglycemic hyperinsulinemic clamp. Preclinical studies have shown that visceral adipose tissue lysates from mice fed with high-fat diet displayed higher concentrations of IgG2c, which are responsible for selective muscle insulin resistance11,12, and therefore, it is conceivable that increased adipose tissue mass may explain the relationship between IgG2 and insulin sensitivity. We found that the association between IgG2 levels and whole-body insulin sensitivity remained statistically significant after adjustment for measures of adiposity such as BMI, and waist circumference, it is thus, unlikely that an expansion of fat mass may fully explain this association.
Alternatively, the association between IgG2 and insulin sensitivity may reflect a generalized subclinical inflammatory state rather than a specific effect of IgG2 isotype on insulin sensitivity. However, we found that the association between IgG2 levels and insulin-stimulated glucose disposal remained statistically significant after adjustment for subclinical inflammatory markers such as hsCRP, complement C3 and white blood cell count; this result arguing against the possibility that a dysregulation in the production of inflammatory molecules may explain the relationship between IgG2 and insulin sensitivity. The view that IgG2 is a bona fide modulator of insulin sensitivity is also supported by the finding that IgG1, IgG3, and IgG4 are not associated with insulin-stimulated glucose disposal. Moreover previous studies have reported that insulin resistance in obese subjects is associated with a distinct profile of IgG11, and that IgG2c and their sialylation levels play a causal role in obesity-induced insulin resistance in mice by modulating insulin endothelial transcytosis via FcγRIIB receptor12.
Our study has some strengths including the demographically homogeneous cohort of offspring of patients with type 2 diabetes, equally comprising both male and female subjects, the wealth of exhaustive anthropometric and metabolic variables collected by a trained staff according to an international standardized protocol, the use of the gold standard hyperinsulinemic euglycemic clamp for insulin sensitivity assessment, and the exclusion of confounding conditions potentially affecting both insulin sensitivity and immune response.
Notwithstanding, the current study has a number of potential limitations that should be recognized. First, the present study has a cross-sectional design, and thus it does not allow us to definitely establish the causal relationship between IgG2 levels and muscle insulin sensitivity or to draw any conclusion on the role of IgG2 in the occurrence of insulin resistance characterizing type 2 diabetes, even though prior preclinical evidence supports the biological plausibility of the present findings. Second, the present results might have been influenced by the presence of a family history of type 2 diabetes. Indeed, it has been reported that parental diabetes is associated with a significant increase in total IgG in the offspring10. Relatives share not only genetic determinants influencing the immune response but also environmental factors such as exposure to infections. Therefore, the extension of the present results to the general population must be made with caution. Third, the present results are only based on White individuals, and generalizing them to other ethnic groups may be inappropriate, because differences between ethnic groups in concentrations of IgG, IgA, and IgM have been reported16. Moreover, we did not have access to direct, detailed measures of hepatic insulin resistance by clamp combined with tracer technique since it is complex and expensive, and therefore not feasible for large epidemiological studies. However, we have employed validated proxy measures of hepatic insulin resistance including the liver IR index and glucose0–30 (AUC) × insulin0–30 (AUC), which contain more information compared with fasting based measures utilized for the HOMA-IR index14,15. Finally, we cannot determine which potential underlying mechanism might induce an elevation in serum IgG2 in insulin resistant individuals. In this regard it should be noted that IgG2 isotype is strongly involved in the responses to bacterial capsular polysaccharide antigens17. Accordingly, previous studies have described increased circulating levels of IgG2 in individuals with periodontitis, predominantly reactive against the major oral pathogens, A. actinomycetemcomitans and P. gingivalis18,19,20. On the other hand several evidences have demonstrated the association between periodontal disease and the risk to develop type 2 diabetes supporting the idea that microbial-induced chronic activation of immune response may exert deleterious effects on glucose homeostasis21,22,23. Numerous studies have consistently pointed to a link between stimulation of toll-like receptors (TLRs), by binding of bacterial antigens, activation of pro-inflammatory signaling pathways and insulin resistance24,25,26. Notably TLRs not only play a critical role in innate immunity but are also essential in the modulation of humoral responses27,28. In particular, TLR-4 activated by bacterial lipopolysaccharide (LPS) elicits IgG2 response by promoting Th1 associated cytokines production such as interleukin-12 (IL-12) and interferon gamma (IFN-γ)29,30. Importantly, alteration of gut microbiota or intestinal permeability, as observed in insulin resistance-related conditions, has been shown to increase gut-derived LPS circulating levels, thus resulting in systemic activation of TLR4-driven immune response25,31,32.
Considering these data we can hypothesize that all these mechanisms may be responsible for increased production of IgG2, which, in turn, leads to insulin resistance, however we cannot exclude that the increased levels of IgG2 found in subjects with a higher degree of insulin resistance represent an epiphenomenon of other alterations, directly involved in the pathogenesis of insulin resistance.
In conclusion the present findings indicate that serum concentrations of IgG2, but not IgG1, IgG3 or IgG4, are associated with insulin-stimulated glucose disposal. The current study is novel because it suggests that the relationship between IgG2 isotype and insulin sensitivity is independent of important determinants of glucose metabolism including gender, age, adiposity, lipids, and subclinical inflammation. Future studies are needed to examine whether IgG2 plays a specific role in the development of type 2 diabetes independently of confounders factors.
Materials and Methods
The study sample comprised 262 non-diabetic offspring of patients with type 2 diabetes participating in the European Network on Functional Genomics of Type 2 Diabetes (EUGENE2) project33. Exclusion criteria included: history of type 1 and type 2 diabetes, malignant diseases, inflammatory bowel diseases and gastrointestinal disorders associated with malabsorption, chronic pancreatitis, use of drugs able to interfere with glucose homeostasis and levels of inflammatory markers such as steroids and oral contraceptive agents, history of alcohol abuse, immunological or rheumatic diseases, acute and chronic infections or positivity for antibodies to hepatitis C virus (HCV) or hepatitis B surface antigen (HBsAg). Individuals were consecutively recruited at the Department of Medical and Surgical Sciences of the University ‘Magna Graecia’ of Catanzaro as previously described33,34. All subjects were of European ancestry and underwent anthropometrical evaluation including measurements of body mass index (BMI), waist circumference, and body composition assessed by bioelectrical impedance. A 75 g oral glucose tolerance test (OGTT) was performed with 0, 30, 60, 90 and 120 min sampling for plasma glucose and insulin assays. Insulin sensitivity was determined by euglycemic hyperinsulinemic clamp study, as previously described34. Briefly, a priming dose of insulin (Humulin, Eli Lilly & Co., Indianapolis, IN) was administered during the initial 10 min to acutely raise plasma insulin followed by continuous insulin infusion fixed at 40 mU/m2 × min. The blood glucose level was maintained constant during the 2-h clamp study by infusing 20% glucose at varying rates according to blood glucose measurements assessed by a glucose analyzer at 5 minute intervals (mean coefficient of variation of blood glucose was <5%).
The study was approved by the local ethics committee (Comitato Etico Azienda Ospedaliera “Mater Domini”). Written informed consent was obtained from each participant before commencing the study, in accordance with the principles of the Declaration of Helsinki.
Analytical determinations
Glucose, triglycerides, total and high density lipoprotein (HDL) cholesterol concentrations were determined by enzymatic methods (Roche, Basel, Switzerland). Serum insulin concentrations were determined with a chemiluminescence-based assay (Immulite®, Siemens, Italy). White blood cell count was determined using an automated particle counter (Siemens Healthcare Diagnostics ADVIA® 120/2120 Hematology System, Italy). Serum levels of high sensitivity C reactive protein (hsCRP), complement C3, total IgG, IgG1, IgG2, IgG3 and IgG4 subclasses were assayed by an automated nephelometric technology using the BN™II System analyzer with high levels of sensibility and specificity (Siemens Healthcare, Italy). Glycated hemoglobin (HbA1c) was assessed by high performance liquid chromatography employing a National Glycohemoglobin Standardization Program certified automated analyzer (Adams HA-8160 HbA1C analyzer, Menarini, Italy).
Calculations
Glucose disposal was calculated as the mean rate of glucose infusion measured during the last 60 min of the clamp examination (steady-state) and it is expressed as milligrams per minute per kilogram fat-free mass measured with the use of electrical bioimpedance. The HOMA-IR index was calculated as fasting insulin × fasting glucose/22.535. The liver IR index was calculated using the formula: −0.091 + (log insulin area under the curve [AUC] 0–120 min × 0.400) + (log fat mass % × 0.346) − (log HDL Cholesterol × 0.408) + (log BMI × 0.435)15. The glucose0–30 (AUC) × insulin0–30 (AUC) index of hepatic resistance to insulin was calculated as reported14. The muscle insulin sensitivity index was calculated as the rate of decline in plasma glucose concentration divided by mean plasma insulin concentration during OGTT as described by Abdul-Ghani MA et al.14. The QUICKI index was calculated using the formula: 1/(log insulin0 + log glucose0) as reported36. The Stumvoll ISIOGTT was calculated as 0.226 − 0.0032 × BMI − 0.0000645 × Insulin120 − 0.00375 × Glucose9037. The Matsuda index was calculated as 10,000/square root of [fasting glucose (mmol/L) × fasting insulin (mU/L)] × [mean glucose × mean insulin during OGTT]38. The Gutt’s ISI0,120 index insulin sensitivity was calculated using the formula: (m/[(G0 + G120)/2])/log [(I0 + I120)/2] where m = (75,000 mg + (fasting glucose − 2-h glucose) × 0.19 × body weight)/120 min (glucose in mg/dl; insulin in µIU/ml) as previously described39.
Statistical analysis
The results for continuous variables are given as means ± SD. Variables with skewed distribution including triglycerides, fasting insulin, HOMA-IR index, glucose0–30 (AUC) × insulin0–30 (AUC) index, Gutt’s ISI0,120, Stumvoll ISIOGTT, Matsuda, muscle insulin sensitivity index and hsCRP were natural log transformed for statistical analyses. Unpaired Student’s t was used to compare differences of continuous variables between two groups. Relationships between variables were determined by Pearson’s correlation coefficient (r). Partial correlation coefficients adjusted for age and gender were computed between variables. A multivariable linear regression analysis was performed in order to evaluate the independent contribution of IgG isotypes and other inflammatory and metabolic variables to insulin sensitivity. The variance inflection factor (VIF) was less than 2 in all the analyses indicating that multicollinearity among variables was not a problem in the multiple regression models. The normality of the standardized residuals was tested using the Kolmorogov-Smirnov test that was >0.05 indicating a normal distribution of the residuals in the multiple regression model.
All tests were two-sided, and a P value < 0.05 was considered statistically significant. All analyses were performed using SPSS (Chicago, IL, USA) software programme Version 22.0 for Windows.
Ethical approval and informed consent statement
The study was approved by the local ethics committee (Comitato Etico Azienda Ospedaliera “Mater Domini”). Written informed consent was obtained from each study participant. All procedures were performed in accordance with the principles of the Declaration of Helsinki.
References
Pickup, J. C. & Crook, M. A. Is type II diabetes mellitus a disease of the innate immune system? Diabetologia 41, 1241–1248 (1998).
Schmidt, M. I. et al. Markers of inflammation and prediction of diabetes mellitus in adults (Atherosclerosis Risk in Communities study): a cohort study. Lancet 353, 1649–1652 (1999).
Ford, E. S. Leukocyte count, erythrocyte sedimentation rate, and diabetes incidence in a national sample of US adults. Am J Epidemiol 155, 57–64 (2002).
Duncan, B. B. et al. Atherosclerosis Risk in Communities Study. Low-grade systemic inflammation and the development of type 2 diabetes. The Arteriosclerosis risk in communities study. Diabetes 52, 1799–1805 (2003).
Engstrom, G., Hedblad, B., Eriksson, K. F., Janzon, L. & Lindgärde, F. Complement C3 is a risk factor for the development of diabetes: a population-based cohort study. Diabetes 54, 570–575 (2005).
Perticone, F. et al. Endothelial Dysfunction and CRP are risk factors for diabetes in essential hypertension. Diabetes 57, 167–171 (2008).
Twig, G. et al. White blood cells count and incidence of type 2 diabetes in young men. Diabetes Care 36, 276–82 (2013).
Rodríguez-Segade, S., Camiña, M. F., Paz, J. M. & Del Río, R. Abnormal serum immunoglobulin concentrations in patients with diabetes mellitus. Clin Chim Acta 203, 135–42 (1991).
Ardawi, M. S., Nasrat, H. A. & Bahnassy, A. A. Serum immunoglobulin concentrations in diabetic patients. Diabet Med 11, 384–7 (1994).
Lindsay, R. S., Krakoff, J., Hanson, R. L., Bennett, P. H. & Knowler, W. C. Gamma globulin levels predict type 2 diabetes in the Pima Indian population. Diabetes 50, 1598–603 (2001).
Winer, D. A. et al. B cells promote insulin resistance through modulation of T cells and production of pathogenic IgG antibodies. Nat Med 17, 610–617 (2011).
Tanigaki, K. et al. Hyposialylated IgG activates endothelial IgG receptor FcγRIIB to promote obesity-induced insulin resistance. J Clin Invest 128, 309–322 (2018).
DeFronzo, R. A. & Tripathy, D. Skeletal muscle insulin resistance is the primary defect in type 2 diabetes. Diabetes Care 2, S157–63 (2009).
Abdul-Ghani, M. A., Matsuda, M., Balas, B. & DeFronzo, R. A. Muscle and liver insulin resistance indexes derived from the oral glucose tolerance test. Diabetes Care 30, 89–94 (2007).
Vangipurapu, J. et al. A novel surrogate index for hepatic insulin resistance. Diabetologia 54, 540–543 (2011).
Mili, F., Flanders, W. D., Boring, J. R., Annest, J. L. & Destefano, F. The associations of race, cigarette smoking, and smoking cessation to measures of the immune system in middle-aged men. Clin Immunol Immunopathol 59, 187–200 (1991).
Siber, G. R., Schur, P. H., Aisenberg, A. C., Weitzman, S. A. & Schiffman, G. Correlation between serum IgG-2 concentrations and the antibody response to bacterial polysaccharide antigens. N Engl J Med 303, 178–82 (1980).
Graswinckel, J. E., van der Velden, U., van Winkelhoff, A. J., Hoek, F. J. & Loos, B. G. Plasma antibody levels in periodontitis patients and controls. J Clin Periodontol 31, 562–8 (2004).
Polak, B. et al. IgG antibody subclass response to Porphyromonas gingivalis outer membrane antigens in gingivitis and adult periodontitis. J Periodontol 66, 363–8 (1995).
Kikuchi, T. et al. Dendritic cells stimulated with Actinobacillus actinomycetemcomitans elicit rapid gamma interferon responses by natural killer cells. Infect Immun 72, 5089–96 (2004).
Demmer, R. T., Jacobs, D. R. Jr. & Desvarieux, M. Periodontal disease and incident type 2 diabetes: results from the First National Health and Nutrition Examination Survey and its epidemiologic follow-up study. Diabetes Care 31, 1373–9 (2008).
Miyawaki, A., Toyokawa, S., Inoue, K., Miyoshi, Y. & Kobayashi, Y. Self-Reported Periodontitis and Incident Type 2 Diabetes among Male Workers from a 5-Year Follow-Up to MY Health Up Study. Plos One 11, e0153464 (2016).
Watanabe, K., Petro, B. J., Shlimon, A. E. & Unterman, T. G. Effect of periodontitis on insulin resistance and the onset of type 2 diabetes mellitus in Zucker diabetic fatty rats. J Periodontol. 79, 1208–16 (2008).
Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 1, 135–145 (2001).
Shi, H. et al. TLR4 links innate immunity and fatty acid-induced insulin resistance. J Clin Invest 116, 3015–3025 (2006).
Kim, J. J. & Sears, D. D. TLR4 and Insulin Resistance. Gastroenterol Res Pract. 2010 (2010).
Pasare, C. & Medzhitov, R. Control of B-cell responses by Toll-like receptors. Nature 438, 364–8 (2005).
Browne, E. P. Regulation of B-cell responses by Toll-like receptors. Immunology 136, 370–9 (2012).
Kawano, Y., Noma, T. & Yata, J. Regulation of human IgG subclass production by cytokines. IFN-gamma and IL-6 act antagonistically in the induction of human IgG1 but additively in the induction of IgG2. J Immunol 153, 4948–58 (1994).
Re, F. & Strominger, J. L. Toll-like receptor 2 (TLR2) and TLR4 differentially activate human dendritic cells. J Biol Chem 276, 37692–9 (2001).
Spruss, A. et al. Toll-like receptor 4 is involved in the development of fructose-induced hepatic steatosis in mice. Hepatology 50, 1094–104 (2009).
Brun, P. et al. Increased intestinal permeability in obese mice: new evidence in the pathogenesis of nonalcoholic steatohepatitis. Am J Physiol Gastrointest Liver Physiol 292, G518–25 (2007).
Marini, M. A. et al. The Arg972variant in insulin receptor substrate-1 is associated with an atherogenic profile in offspring of type 2 diabetic patients. J Clin Endocrinol Metab 88, 3368–3371 (2003).
Stancáková, A. et al. EUGENE2 Consortium. SNP rs7754840 of CDKAL1 is associated with impaired insulin secretion in non-diabetic offspring of Type 2 diabetic subjects (the EUGENE2 study) and in a large sample of men with normal glucose tolerance. J Clin Endocrinol Metab 93, 1924–1930 (2008).
Matthews, D. R. et al. Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 28, 412–9 (1985).
Katz, A. et al. Quantitative insulin sensitivity check index: a simple, accurate method for assessing insulin sensitivity in humans. J Clin Endocrinol Metab 85, 2402–2410 (2000).
Stumvoll, M. et al. Use of the oral glucose tolerance test to assess insulin release and insulin sensitivity. Diabetes Care 23, 295–301 (2000).
Matsuda, M. & De Fronzo, R. A. Insulin sensitivity indices obtained from oral glucose tolerance testing: comparison with the euglycemic insulin clamp. Diabetes Care 22, 1462–1470 (1999).
Gutt, M. et al. Validation of the insulin sensitivity index (ISI (0, 120)): comparison with other measures. Diabetes Res Clin Pract 47, 177–84 (2000).
Acknowledgements
This work was supported by the Italian Ministry of Education, University and Research, grant n. 2015MPESJS_006 to Giorgio Sesti.
Author information
Authors and Affiliations
Contributions
T.V.F. researched the data, contributed to write the paper, and edited the manuscript; E.S., F.A., F.A. researched data and reviewed the manuscript, A.G., C.P., A.Q. performed measurement of serum IgG levels, F.S. contributed to analyze the data, M.L.H. researched the data, contributed to the discussion and reviewed the manuscript; F.P., A.F., contributed to the discussion and reviewed the manuscript; G.S. designed the study, analyzed the data and wrote the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fiorentino, T.V., Succurro, E., Arturi, F. et al. Serum IgG2 levels are specifically associated with whole-body insulin-mediated glucose disposal in non-diabetic offspring of type 2 diabetic individuals: a cross-sectional study. Sci Rep 8, 13616 (2018). https://doi.org/10.1038/s41598-018-32108-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-32108-8
- Springer Nature Limited
Keywords
This article is cited by
-
Achieving quantitative reproducibility in label-free multisite DIA experiments through multirun alignment
Communications Biology (2023)
-
Improved quantification of muscle insulin sensitivity using oral glucose tolerance test data: the MISI Calculator
Scientific Reports (2019)