
Abstract
In the ever-changing fungal environment, fungi have to cope with a wide array of very different stresses. These stresses frequently act in combination rather than independently, i.e., they quickly follow one another or occur concomitantly. Combinatorial stress response studies revealed that the response of fungi to a stressor is highly dependent on the simultaneous action of other stressors or even on earlier stresses to which the fungi adapted. Several important phenomena were discovered, such as stress pathway interference, acquired stress tolerance, stress response memory or stress cross-protection/sensitization, which cannot be interpreted when we study the consequences of a single stressor alone. Due to the interactions between stressors and stress responses, a stress response that develops under a combined stress is not the simple summation of stress responses observed during single stress treatments. Based on the knowledge collected from single stress treatment experiments, we cannot predict how fungi will respond to a certain combination of stresses or even whether this combination will be more harmful than single stress treatments. This uncertainty warns us that if we want to understand how fungi adapt to a certain habitat (e.g., to the human body) to find a point of weakness in this adaptation, we must understand how the fungi cope with combinations of stresses, rather than with single stressors.
Similar content being viewed by others

Introduction
The stress concept has become very widespread and popular in the biological sciences since H. Selye first published his theory in 1936 (Selye 1936). Since that time, the definition of stress has changed considerably, and by now, the term “stress” does not necessarily mean the same thing for every scientist. This is also true for mycologists. There is no clear consensus in using the terms “stressed” and “unstressed” cultures (Hohmann and Mager 2003a, b; Thammavongs et al. 2008; Hallsworth 2018; Rangel et al. 2018; Antal et al. 2020), which can lead to misunderstanding of the results of others or to misinterpretation of our own results. Here, we use a stress definition that is close to the original definition of Selye (1976). Accordingly, the stress-free state is a kind of balance between the fungus and its environment. Any external factors that disturb this balance are regarded as stressors, the resulting imbalance is regarded as stress, and the sum of adaptation processes aiming to reach a new balance is regarded as a stress response. When we speak about the stress tolerance of a fungus, we characterize the success of the stress response from the perspective of a specific feature of the fungus (e.g., growth, conidia formation or viability of cells). Importantly, this stress definition means that, for example, the addition of a toxic heavy metal ion (e.g., Cd2+) to the culture media or limiting the availability of an important metal ion (e.g., Fe2+/Fe3+) can cause stress, but thereafter, as the fungus adapts to the new conditions, living in the presence of Cd2+ or at a low level of Fe2+/Fe3+ can be suboptimal for growth or conidia formation but not stress.
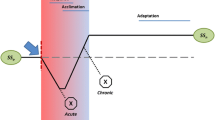
Studying the stress responses of fungi means studying the life of fungi not under their optimal culture conditions but under well-controlled disruptions of culture conditions to observe how fungi behave “in action.” This approach has great theoretical significance. It helps us to elucidate how fungi sense changes in their environment, how they can adapt to these changes and how their regulatory network meets these challenges (Hohmann and Mager 2003a, b; Atay and Skotheim 2017; Brown et al. 2017). A deeper understanding of the stress biology of fungi will help answer questions of practical importance as well. These questions include how fungi survive in the body in immunocompromised patients, how they behave in stored food and feed, on art treasures or in fermenters, how stress alters their mycotoxin production or how they can adapt to fungicides. In recent decades, a large number of studies have examined fungal stress responses initiated by a specific stressor (Abad et al. 2010; Morano et al. 2012; Hagiwara et al. 2016a, b; Pais et al. 2019). These studies identified several receptors sensing stress, genes, proteins, and metabolites that are important in adaptation to stress and signaling pathways connecting receptors to regulated genes or proteins. In their real habitats, however, fungi have to cope with several stressors sequentially or even concomitantly (Brown et al. 2017). As an example, in the human body, fungi have to face, among others, oxidative and nitrosative stress, carbon, iron, zinc and oxygen limitation, and they also have to adapt to the local pH and temperature or the presence of antimycotics (Cooney and Klein 2008; Abad et al. 2010). The terms “combinatorial stress” and “combinatorial stress response” were applied systematically in mycology by Kaloriti et al. (2012) for the first time. These authors studied the effects of osmotic, oxidative and nitrosative stressors as well as their various combinations on the growth of human pathogenic Candida albicans and Candida glabrata (Kaloriti et al. 2012; Brown et al. 2017). Interactions among stressors and/or stress responses can substantially influence the nature and success of adaptation (Brown et al. 2017). The results of combinatorial stress response experiments, therefore, allow us to get one step closer to understanding the real nature of how regulatory networks of fungi work, how they can adapt to their ever-changing environment, or how they can occupy new habitats. Studying combinatorial stresses revealed new aspects of stress responses, such as “stress response memory,” “stress cross-protection” or “stress pathway interference.” Here, we summarize our current knowledge on these features of fungal stress responses. We use “combinatorial stress” or “combinatorial stress responses” as terms in all experiments where the effects of either concomitantly or sequentially acting stressors are investigated (Fig. 1).

Combinatorial stress response experiments Part A: Overview of a typical combinatorial stress response experiment where concomitantly acting stressors are investigated. Part B: Overview of a typical combinatorial stress response experiment where sequentially acting stressors are investigated. The effect of pretreatment can be either permanent (cells are treated with stressor 2 in the presence of stressor 1) or temporal (cells are treated with stressor 2 after removal of stressor 1)
Adaptation to concomitantly acting stressors–stress pathway interference.
In the case of concomitantly acting stressors, two parameters/components (at least) of the culture conditions are varied, and fungi have to adapt to both of them concomitantly (Figs. 1A, 2A and 2B). Most of the studies published in this field study the effect of either combined treatment with two antifungal or concomitantly acting physiologically relevant stresses (e.g., oxidative, nitrosative, cationic stresses) drugs on human pathogenic fungi.

Possible outcome of combinatorial stress response experiments Part A and B: Candida albicans Ca372 cultures were treated either with 1 M NaCl (red), 5 mM H2O2 (blue) or 1 M NaCl + 5 mM H2O2 concomitantly (purple) or were kept untreated (yellow). Figures were prepared according to data presented by Kaloriti et al. (2014). Part A: Distribution of upregulated genes among the stress-treated cultures. Upregulated genes were defined as genes showing a consistent ≥ 2.5-fold increase in their normalized microarray signal intensity in three independent experiments. Untreated cultures were used as references. Part B: Antifungal activity of the stress treatments. Mean ± SD calculated from three independent experiments are presented. Values marked with the same letter do not differ from one another significantly according to Dunnett’s t test (p < 0.05). Part C and D: Aspergillus fumigatus Af293 was cultured at sufficient iron concentrations (yellow and blue) and under iron-limited conditions (red and purple). After adaptation to iron availability, some of the cultures were treated with 5 mM H2O2 (yellow and purple) (sequentially treated cultures). Figures were prepared according to data presented by Kurucz et al. (2018). Part C: 2′,7′-Dichlorofluorescein (DCF) production of Aspergillus fumigatus Af293 cultures. DCF production is a characteristic of redox imbalance. Mean ± SD calculated from three independent experiments are presented. Values marked with the same letter do not differ from one and other significantly according to Student’s t test (p < 0.05). Part D: Principal component analysis of transcriptome data (RNA sequencing)
Synergistic interactions are frequently published between stresses induced by different antifungal drugs. From a therapeutic point of view, a very promising interaction is the synergistically increased antifungal effect found in the combined echinocandin (e.g., caspofungin, micafungin, anidulafungin) plus nikkomycin Z treatment (Lima et al. 2019). This treatment was efficient against various human pathogenic fungi, their biofilms and even some echinocandin-resistant strains (Cheung and Hui 2017; Kovács et al. 2019; Lima et al. 2019). Echinocandins inhibit the synthesis of the fungal cell wall polysaccharide β-1,3-glucan, and fungi frequently respond to this stress with enhanced chitin synthesis (“compensatory chitin synthesis”) (Walker et al 2010; Emri et al. 2013). Nikkomycin Z inhibits chitin synthesis, which explains the observed synergistic interactions (Walker et al 2010; Lima et al. 2019). Unfortunately, the results of intensive studies with combination antifungal therapies show that the effects of concomitantly acting stressors are only additive in many cases, and in addition to synergistic interactions, antagonistic interactions also occur (Tóth et al. 2013; Campitelli et al. 2017; Wambaugh et al. 2020). The well-known antagonism between amphotericin B and azole antifungals is explained by the fact that amphotericin B has to bind to ergosterol to destroy the cell membrane, but azoles inhibit ergosterol biosynthesis, leading to decreased availability of the target of amphotericin B (Sugar and Liu 1998; Campitelli et al. 2017).
In other experiments, combinatorial osmotic/cationic (NaCl) plus oxidative (H2O2) stress as well as combinatorial oxidative (H2O2) plus nitrosative stress (dipropylenetriamine NONOate) resulted in a more extended lag phase (adaptation time) than the corresponding individual stresses in both C. albicans and C. glabrata cultures (Kaloriti et al. 2012). In contrast, combinatorial osmotic/cationic plus nitrosative stress had no similar (synergistic) effect in the same experiment, demonstrating again that synergism is specific to certain combinations of stresses and does not occur in the combination of any two stresses (Kaloriti et al. 2012). The observed synergisms have great medical significance since they contribute to the efficiency with which neutrophils kill fungal cells by combined attack of reactive oxygen species, reactive nitrogen species and cations (Nüsse 2011), as experimentally demonstrated by Kaloriti et al. (2014). In the case of combinatorial osmotic/cationic plus oxidative stress, the decreased stress tolerance, relative to the effect of single stress treatments, was also characterized by decreased viability of the cells (Kaloriti et al. 2014). More importantly, microarray experiments demonstrated that the stress response to the combined stress treatment is unique, and it is not a simple summation of stress responses recorded in single stress treatments (Kaloriti et al. 2014; Fig. 2A). The synergistic interaction can be explained by “stress pathway interference” (Kaloriti et al. 2014), which is a kind of antagonistic cross-talk between stress signaling pathways. In the case of combinatorial NaCl plus H2O2 stress, synergism is the consequence of the inhibitory effect of cations on catalase. It leads to hyperaccumulation of reactive oxygen species and inactivation of the Cap1 transcription factor, which is responsible for the initiation of the oxidative stress response (Kaloriti et al. 2014; Kos et al. 2016). Moreover, cations also stimulate the nuclear exclusion of Cap1 by the Crm1 nuclear export protein (Kos et al. 2016). These factors all lead to reduced oxidative stress tolerance under cationic stress and to a stress response that differs from both the cationic and oxidative stress responses. Interactions at the level of signaling pathways cannot be ruled out in the case of combined antifungal therapy. The existence of stress pathway interference demonstrates that despite their commonly high stress tolerance and their superb ability to adapt to continuously changing and/or new environments, the adaptation potential of fungi is limited. Understanding these limitations is crucial to develop new antifungal strategies.
In the abovementioned experiments, researchers applied treatments that induced stress quickly or at least with similar kinetics to observe their simultaneous effects. Unfortunately, there are time-consuming stress treatments. For example, when we transfer the fungus from glucose to hemicellulose, cells will recognize the absence of glucose first (carbon starvation stress), and they need some time to detect the alternative carbon source and to respond to its presence (carbon limitation stress) (van Munster et al. 2014). Alternatively, to induce iron starvation stress, it is not sufficient to use iron-free medium. We also have to wait until the fungus depletes all of the iron occurring both in the medium as a contaminant and in its intracellular iron stores (Tóth et al. 2009). In the case of such treatments, investigating the effects of concomitantly acting stressors can be problematic since the results highly depend on how efficiently we can synchronize the two treatments. Waiting until successful adaptation to the first stress treatment is achieved and applying the second stress treatment only thereafter (i.e., studying sequentially acting stressors) can be an alternative way to study combinatorial stress responses in these cases.
Adaptation to sequentially acting stressors-acquired stress tolerance.
When we use sequentially acting stressors (Figs. 1B, 2C and 2D), we examine how the adaptation to one stressor modifies the response to another stressor. In these experiments, we can investigate how stress tolerance depends on the composition of applied media (e.g., on the availability of iron or the type of carbon/energy source), how stress tolerance changes over time (e.g., during the shift from exponential growth to stationary phase), and how adaptation to a stressor modifies stress tolerance (e.g., how adaptation to growth at 37 °C modifies antifungal drug susceptibility).
Early studies in this field revealed the “acquired stress tolerance” phenomenon. This means that adaptation to mild stress increases the tolerance of the fungus against a stronger stressor. The acquired thermotolerance of Saccharomyces cerevisiae is a classic example: This yeast is unable to survive at 55 °C if it was cultured at its optimal temperature (28–30 °C) previously. However, it can efficiently adapt to this heat stress after pretreatment at 37 °C (mild heat stress) (Lindquist 1986). Acquired thermotolerance has been observed in several fungi, such as Neurospora crassa, Schizosaccharomyces pombe and C. albicans (Plesofsky-Vig and Tbrambl 1985; De Virgilio et al. 1990; Argüelles 1997), but this phenomenon is not restricted to heat stress. Acquired stress tolerance has been recorded in very different fungi under very different stresses, including oxidative, osmotic, carbon limitation stresses or antifungal drug-induced stresses (Lewis et al. 1995; Izawa et al. 1996; Marr et al. 1998; Brickner et al. 2007; Kundu et al. 2007; Zacharioudakis et al. 2007; Berry and Gasch 2008; Calo et al. 2014). There are two interesting features of acquired stress tolerance (Brown et al. 2014, 2017): 1) The effect of pretreatment can be long-lasting (“stress response memory”) (i.e., the increased stress tolerance can sometimes remain for a surprisingly long time even if the cells, after pretreatment, are transferred to the conditions used before). 2) Adaptation to a mild stressor can lead to increased tolerance against other very different stressors (“stress cross-protection”).
Stress response memory
The molecular background of stress response memory is complex and involves a wide range of different mechanisms. The accumulation of stress metabolites (e.g., glycerol, trehalose, mannitol, glutathione) can increase stress tolerance, but their concentration can rapidly decrease after the effect of stressors has passed (Argüelles 1997; Klipp et al. 2005). Stress-activated proteins (e.g., heat shock proteins, antioxidant enzymes, biosynthetic enzymes of stress metabolites or multidrug transporters), however, can preserve their elevated levels and activities for a longer time; thereafter, the cells return to “unstressed” conditions, resulting in longer retention of acquired tolerance (Jamieson et al. 1996, Marr et al. 1998; Leach et al. 2012; You et al. 2012). As an example, NaCl treatment increased the amount and activity of cytosolic Ctt1p catalase, leading to increased oxidative stress tolerance in S. cerevisiae (Guan et al. 2012). When the culture was transferred to standard conditions, cells preserved their elevated catalase contents and, as a consequence, their increased oxidative stress tolerance for 4–5 generations. This phenomenon did not require elevated ctt1 mRNA levels or nascent protein synthesis; mother cells simply distributed their long-lasting Ctt1p to daughter cells from generation to generation. In this way, (daughter) cells that never experienced stress before showed acquired oxidative stress tolerance (Guan et al. 2012). A faster transcriptional response to stress after stress pretreatment has also been reported in many experiments (“transcriptional memory”) (D'Urso and Brickner 2017). Pretreatment with galactose increases the speed of the transcriptional response to another galactose treatment, and in a similar way, adaptation to inositol starvation results in a faster transcriptional response to a forthcoming withdrawal of inositol in S. cerevisiae (Brickner et al. 2007; Kundu et al. 2007; Zacharioudakis et al. 2007). In the previously mentioned experiment, NaCl pretreatment not only increased oxidative stress tolerance but also led to a faster transcriptional response to oxidative stress (Guan et al. 2012). “Transcriptional memory” is commonly explained by epigenetic regulation (Tribus et al. 2005; Laskar et al. 2015) and usually involves chromatin remodeling and transcriptional poising (D'Urso and Brickner 2017). In both cases, a specific transcription factor binds to the promoter after pretreatment, which leads to altered histone methylation or recruitment of the preinitiation complex in the appropriate region for generations (for details see D'Urso and Brickner; 2017). Noncoding RNAs are also involved in the regulation of stress responses and represent another important source of heritable changes in transcriptional activities (Hirota et al. 2008; Leong et al. 2014). In the human pathogenic Mucor circinelloides, as in other fungi, tacrolimus and rapamycin form complexes with FKBP12 prolyl isomerase. These complexes inhibit calcineurin and Tor kinase, which explain the antifungal activity of tacrolimus and rapamycin, respectively (Heitman et al. 1991; Liu et al. 1991). Mucor circinelloides is able to silence FKBP12 transcription in a small interfering RNA (siRNA)-dependent process, which makes the fungus resistant to these drugs even after growth on drug-free media for several generations (“epimutation”) (Calo et al. 2014).
A special case of the long-lasting effect of pretreatment is the altered stress tolerance of dormant or germinating conidia formed under different culture conditions (Rangel et al. 2015;the conidiogenous mycelia, heat, Hagiwara et al. 2017; de Vries et al. 2017). In this case, the stress tolerance shows alterations only in the next generation (i.e., conidia), but due to conidial dormancy, it has long-lasting consequences. Several factors affecting mycelia before or during conidiogenesis can influence the properties of conidia. In the case of entomopathogenic fungi such as Metarhizium robertsii or M. anisopliae, the effect of nutrient composition, temperature and pH of the media used for culturing the conidiogenous mycelia, heat, hypoxic or osmotic stress treatment applied during cultivation or even illumination can be important (Hallsworth and Magan 1994; Rangel et al. 2008a, 2008b, 2011, 2015). These factors can determine the UV, oxidative or heat tolerance of conidia, the speed or efficiency of their germination and their mycotoxin content or virulence (Hallsworth and Magan 1994; Rangel et al. 2015; Hagiwara et al. 2017). The enhanced stress tolerance of dormant conidia formed by pretreated mycelia is usually explained by the elevated accumulation of multifunctional stress metabolites (e.g., trehalose, glycerol, mannitol, erythritol, arabitol) and pigments (Hallsworth and Magan 1994, 1995, 1996; Rangel et al. 2015). In addition to its metabolite content, the transcriptome of conidia also depends on the culture conditions. The altered abundance of certain mRNAs related to stress protection caused by pretreatment can affect the stress tolerance attributes of germinating conidia (Sakamoto et al. 2009; Hagiwara et al. 2016a, b).
Stress cross-protection
Stress cross-protection is also the result of very different mechanisms. Stress metabolites are multifunctional and can protect cells against many different stressors. As an example, glutathione is not only a widespread and abundant antioxidant in fungi but also reacts with xenobiotics or toxic aldehydes formed during metabolism, forms inert complexes with heavy metals such as Cd2+, protects proteins during desiccation, is important in the stabilization of membranes and serves as an S, N or energy/C source during nutrient starvation (Pócsi et al. 2004). Meanwhile, trehalose prevents lipid peroxidation, protects proteins against denaturation and aggregation, helps cells survive desiccation or freeze–thaw stress or can be used as a stored energy/C source (Eleutherio et al. 2015; Tapia et al. 2015; Magalhães et al. 2018). Therefore, the accumulation of stress metabolites (e.g., in conidia) or the enzymes responsible for their production (e.g., in vegetative cells) after pretreatment can enhance stress tolerance against other stressors during subsequent treatment. In addition, very different stressors frequently cause very similar physiological changes in the cells. Oxidative stress can be induced by oxidative agents, xenobiotics, Fenton or even non-Fenton metal ions; different types of radiation, desiccation, carbon starvation, iron limitation, and elevated temperature can also lead to unbalanced redox homeostasis (Lushchak 2010; Brown et al. 2017). Injuries of the cell wall and cell membrane components (“cell integrity stress”) or denaturation of proteins are also common consequences of different stress treatments (Hohmann and Mager 2003a, b; Gasch 2007; Brown et al. 2017). Therefore, upregulation of processes aiming to maintain redox homeostasis, the integrity of cells and protect proteins from denaturation and aggregation or degrade the injured proteins can also contribute to cross-protection. However, experimental and theoretical approaches suggest that stress cross-protection is more than a simple consequence of the multifunctionality of stress metabolites or the overlapping consequences of different stressors. Instead, it is an important element of fungal strategies to survive in a continuously changing environment. The existence of the core stress response (environmental stress response) in fungi is a good example.
The core stress response means that a set of stress genes responds in the same manner to very different stressors (Gasch 2007; Pradhan et al. 2021). During the core stress response, many genes involved in vegetative growth are downregulated, and among others, several stress genes are upregulated (Gasch et al. 2000, 2007; Chen et al. 2003; Enjalbert et al. 2006; Roetzer et al. 2008; Antal et al. 2020). Upregulated stress genes have versatile functions, and their upregulation can explain stress cross-protection (Gasch et al. 2007). The core stress response has been described in S. cerevisiae (Gasch et al. 2000), S. pombe (Chen et al. 2003), Candida glabrata (Roetzer et al. 2008), and C. albicans (Enjalbert et al. 2006; Brown et al. 2020) and may also exist in other fungi (Gasch 2007; Antal et al. 2020). The number of core stress response genes and their regulation depend on the species. In S. cerevisiae, in which the most core stress response genes were identified (approximately 900 genes), this part of the stress response is mainly under the control of the functionally redundant Msn2p and Msn4p transcription factors (Gasch et al. 2000, 2007). In C. glabrata, C. albicans and S. pombe, where many fewer core stress response genes were identified, these genes are regulated by the Msn2p transcription factor, the Hog1 stress-activated protein kinase and Cap1p transcription factor, and the Hog1 ortholog Sty1p, respectively (Chen et al. 2003; Enjalbert et al. 2006; Roetzer et al. 2008).
There are two theories explaining the existence of a core stress response in fungi:
-
(1)
Continual changes are not necessarily equal to chaotic changes. Stresses can follow each other in a kind of order: e.g., an increase in the temperature can lead to drying of wet habitats, which means the heat stress is followed by an osmotic stress and later, when cells are no longer in water, by oxidative and starvation stresses. Therefore, mechanisms that upregulate some stress response elements to cope with the actual stressor and others to prepare for the most likely subsequent stress(es) can be evolutionarily advantageous (“adaptive prediction”/ “anticipatory behavior”) (Mitchell et al. 2009; Brown et al. 2019).
-
(2)
In conditions suitable for fast growth (e.g., in the presence of a sufficient amount of glucose as an energy/C source and at optimal or close to optimal pH, temperature, aeration, osmotic concentration, redox state, etc.), a quick invasion of the area where the fungus lives (before the competitors do so) can also be an evolutionarily preferred strategy. Upregulation of any biochemical/cellular processes that support fast growth and downregulation of processes that are not necessary for fast growth (e.g., unessential parts of stress protection machinery) can be meaningful elements of this strategy (“fast growth with reduced stress protection”). When we use fast-growing cultures as a reference, any stress that prevents intensive growth can promote fungi to abandon this strategy. As a consequence, vegetative growth will be downregulated, and many processes that were downregulated before including stress protection mechanisms will be upregulated. From this perspective, stress (pretreatment) does not increase the stress tolerance of the fungus to prepare for subsequent stress, but adaptation to unique fast-growing conditions decreased it previously (Antal et al. 2020).
Stress cross-protection is not necessarily symmetric. As an example, heat stress increases the oxidative stress tolerance of both S. cerevisiae and C. albicans; however, oxidative stress does not protect against subsequent thermal stress (Enjalbert et al. 2003; Mitchell et al. 2009). Asymmetric responses can be explained with the abovementioned adaptive prediction theory (Mitchell et al. 2009; Brown et al. 2019): Intensive fermentation leads to increased temperature and quick depletion of glucose, which switches on respiration in S. cerevisiae. Elevated respiration increases the formation of reactive oxygen species, which should be controlled by upregulated oxidative stress protection (Mitchell et al. 2009). Meanwhile, in the case of C. albicans, it is speculated that adaptation mechanisms to fever caused by inflammatory responses should protect cells not only against high temperature but also against oxidative attack by phagocytic cells (Brown et al. 2014). In both cases, the increase in temperature predicts subsequent oxidative stress, but oxidative stress is not a sign of forthcoming heat stress.
Stress cross-protection is not an obligatory property of stress responses. Adaptation to a stressor can either increase or decrease (“stress cross-sensitization”) or simply not modify stress tolerance against other stressors. Although pretreatment of C. albicans with thermal stress leads to elevated oxidative protection, it does not increase osmotic or cell wall stress tolerance (Enjalbert et al. 2003). Carbon limitation stress-induced oxidative stress tolerance in S. cerevisiae (Maris et al. 2001), while iron limitation stress increased oxidative stress sensitivity of the human pathogenic Aspergillus fumigatus (Kurucz et al. 2018; see also Fig. 2C and 2D). Growth on lactate (carbon limitation) induced osmotic and cell wall stress tolerance and decreased caspofungin, tunicamycin and amphotericin B susceptibility of C. albicans; however, it reduced miconazole tolerance (Ene et al. 2012). In the case of C. glabrata, growth on lactate (or other physiologically relevant carbon sources such as acetate or oleate) reduced amphotericin B susceptibility but increased oxidative stress sensitivity (Chew et al. 2019). In the case of Cryptococcus neoformans, iron limitation increased caspofungin, tunicamycin and amphotericin B antifungal drug susceptibility but decreased susceptibility to voriconazole and itraconazole (Lai et al. 2016).
Stress cross-sensitization, the sometimes asymmetric nature of stress cross-protection and the occasionally conflicting results of different research teams (i.e., during one experiment, there was stress cross-protection, and during another, very similar experiment there was not; Święciło 2016) suggest that the interaction between the two stressors cannot be explained completely with the core stress response. Instead, the consequences of the interaction between the applied stressors highly depend on the type and strength of the stressors and even on the culture conditions (Święciło 2016; Antal et al. 2020). Some of these interactions can be explained by the biochemical and/or ultrastructural consequences of the pretreatment (Ene et al. 2012; Kurucz et al. 2018; Chew et al. 2019), the evolutionarily designed workings of the cellular regulatory network (adaptive prediction; Mitchell et al. 2009) or simply by the fact that even the most versatile regulatory network cannot guarantee successful adaptation to all possible stress combinations (stress pathway interference; Kaloriti et al. 2014).
Conclusions for the future of biology
Stress pathway interference, stress response memory, stress cross-protection or sensitization phenomena demonstrate that stress responses developed during a combined stress experiment are not the simple summation of stress responses observed during single stress treatments. The few omical studies carried out in this field clearly support this view (Kaloriti et al. 2014, Owens et al. 2014, Kurucz et al. 2018; see also Fig. 2B and 2D). Combinatorial stresses represent new types of stress for fungi that require new strategies to cope with them. Based on the knowledge collected from single stress treatment experiments, we cannot predict how fungi will respond to a combination of stresses or whether this combination will be more harmful than single stress treatments alone. Despite the clear achievements in this field, we are still far from understanding how the cellular regulatory network of fungi functions or the rationale underlying their stress response. Nevertheless, the results of the last decade have brought new ideas. We have realized that if we want to understand how fungi adapt to a certain habitat (e.g., the human body) and we want to find points of weakness of these adaptations, we have to understand how they cope with the combination of stresses rather than with single separate stressors. When we study in vitro how fungi behave in a complex environment, the results can be misleading since a stress that is harmful in vitro can, in combination with other stresses, be negligible in a real habitat. The opposite can also occur. Compounds that proved unsuitable to interfere with fungal growth in vitro or exerted effects that were not selective enough can kill microbes efficiently and selectively in vivo. Results of the combinatorial stress response studies, therefore, can initiate substantial changes in the methodology of stress microbiology in the near future. New, efficient methods are foreseeable to be developed to characterize drug susceptibility under stress. The classic culture conditions that allow fungi to grow rapidly (“stress-free conditions”) will be replaced with conditions that more closely resemble the in vivo ones when stress responses are studied. Above all, the scientific significance of studies aiming to record stress responses in vivo will increase.
References
Abad A, Fernández-Molina JV, Bikandi J, Ramírez A, Margareto J, Sendino J, Hernando FL, Pontón J, Garaizar J, Rementeria A (2010) What makes aspergillus fumigatus a successful pathogen? genes and molecules involved in invasive aspergillosis. Rev Iberoam Micol 27:155–182. https://doi.org/10.1016/j.riam.2010.10.003
Antal K, Gila BC, Pócsi I, Emri T (2020) General stress response or adaptation to rapid growth in Aspergillus nidulans? Fungal Biol 124:376–386. https://doi.org/10.1016/j.funbio.2019.10.009
Argüelles JC (1997) Thermotolerance and trehalose accumulation induced by heat shock in yeast cells of Candida albicans. FEMS Microbiol Lett 146:65–71. https://doi.org/10.1111/j.1574-6968.1997.tb10172.x
Atay O, Skotheim JM (2017) Spatial and temporal signal processing and decision making by MAPK pathways. J Cell Biol 216:317–330. https://doi.org/10.1083/jcb.201609124
Berry DB, Gasch AP (2008) Stress-activated genomic expression changes serve a preparative role for impending stress in yeast. Mol Biol Cell 19:4580–4587. https://doi.org/10.1091/mbc.e07-07-0680
Brickner DG, Cajigas I, Fondufe-Mittendorf Y, Ahmed S, Lee PC, Widom J, Brickner JH (2007) H2A.Z-mediated localization of genes at the nuclear periphery confers epigenetic memory of previous transcriptional state. PLoS Biol 5:e81. https://doi.org/10.1371/journal.pbio.0050081
Brown AJ, Budge S, Kaloriti D, Tillmann A, Jacobsen MD, Yin Z, Ene IV, Bohovych I, Sandai D, Kastora S, Potrykus J, Ballou ER, Childers DS, Shahana S, Leach MD (2014) Stress adaptation in a pathogenic fungus. J Exp Biol 217:144–155. https://doi.org/10.1242/jeb.088930
Brown AJP, Cowen LE, di Pietro A, Quinn J (2017) Stress adaptation. Microbiol Spectr 5(4):5–4. https://doi.org/10.1128/microbiolspec.FUNK-0048-2016
Brown AJP, Gow NAR, Warris A, Brown GD (2019) Memory in fungal pathogens promotes immune evasion, colonisation, and infection. Trends Microbiol 27:219–230. https://doi.org/10.1016/j.tim.2018.11.001
Brown AJP, Larcombe DE, Pradhan A (2020) Thoughts on the evolution of core environmental responses in yeasts. Fungal Biol 124:475–481. https://doi.org/10.1016/j.funbio.2020.01.003
Calo S, Shertz-Wall C, Lee SC, Bastidas RJ, Nicolás FE, Granek JA, Mieczkowski P, Torres-Martínez S, Ruiz-Vázquez RM, Cardenas ME, Heitman J (2014) Antifungal drug resistance evoked via RNAi-dependent epimutations. Nature 513:555–558. https://doi.org/10.1038/nature13575
Campitelli M, Zeineddine N, Samaha G, Maslak S (2017) Combination antifungal therapy: a review of current data. J Clin Med Res 9:451–456. https://doi.org/10.14740/jocmr2992w
Chen D, Toone WM, Mata J, Lyne R, Burns G, Kivinen K, Brazma A, Jones N, Bähler J (2003) Global transcriptional responses of fission yeast to environmental stress. Mol. Biol. Cell 14:214–229. https://doi.org/10.1091/mbc.e02-08-0499
Cheung YY, Hui M (2017) Effects of echinocandins in combination with nikkomycin z against invasive Candida albicans bloodstream isolates and the fks mutants. Antimicrob Agents Chemother 61:e00619-17. https://doi.org/10.1128/AAC.00619-17
Chew SY, Ho KL, Cheah YK, Sandai D, Brown AJP, Than LTL (2019) Physiologically relevant alternative carbon sources modulate biofilm formation, cell wall architecture, and the stress and antifungal resistance of Candida glabrata. Int J Mol Sci 20:3172. https://doi.org/10.3390/ijms20133172
Cooney NM, Klein BS (2008) Fungal adaptation to the mammalian host: it is a new world, after all. Curr Opin Microbiol 11:511–516. https://doi.org/10.1016/j.mib.2008.09.018
De Virgilio C, Simmen U, Hottiger T, Boller T, Wiemken A (1990) Heat shock induces enzymes of trehalose metabolism, trehalose accumulation, and thermotolerance in Schizosaccharomyces pombe, even in the presence of cycloheximide. FEBS Lett 273:107–110. https://doi.org/10.1016/0014-5793(90)81062-s
de Vries RP, Riley R, Wiebenga A, Aguilar-Osorio G, Amillis S, Uchima CA, Anderluh G, Asadollahi M, Askin M, Barry K, Battaglia E, Bayram Ö, Benocci T, Braus-Stromeyer SA, Caldana C, Cánovas D, Cerqueira GC, Chen F, Chen W, Choi C, Clum A, Dos Santos RA, Damásio AR, Diallinas G, Emri T, Fekete E, Flipphi M, Freyberg S, Gallo A, Gournas C, Habgood R, Hainaut M, Harispe ML, Henrissat B, Hildén KS, Hope R, Hossain A, Karabika E, Karaffa L, Karányi Z, Kraševec N, Kuo A, Kusch H, LaButti K, Lagendijk EL, Lapidus A, Levasseur A, Lindquist E, Lipzen A, Logrieco AF, MacCabe A, Mäkelä MR, Malavazi I, Melin P, Meyer V, Mielnichuk N, Miskei M, Molnár ÁP, Mulé G, Ngan CY, Orejas M, Orosz E, Ouedraogo JP, Overkamp KM, Park HS, Perrone G, Piumi F, Punt PJ, Ram AF, Ramón A, Rauscher S, Record E, Riaño-Pachón DM, Robert V, Röhrig J, Ruller R, Salamov A, Salih NS, Samson RA, Sándor E, Sanguinetti M, Schütze T, Sepčić K, Shelest E, Sherlock G, Sophianopoulou V, Squina FM, Sun H, Susca A, Todd RB, Tsang A, Unkles SE, van de Wiele N, van Rossen-Uffink D, Oliveira JV, Vesth TC, Visser J, Yu JH, Zhou M, Andersen MR, Archer DB, Baker SE, Benoit I, Brakhage AA, Braus GH, Fischer R, Frisvad JC, Goldman GH, Houbraken J, Oakley B, Pócsi I, Scazzocchio C, Seiboth B, vanKuyk PA, Wortman J, Dyer PS, Grigoriev IV (2017) Comparative genomics reveals high biological diversity and specific adaptations in the industrially and medically important fungal genus Aspergillus. Genome Biol 18:28. https://doi.org/10.1186/s13059-017-1151-0
D’Urso A, Brickner JH (2017) Epigenetic transcriptional memory. Curr Genet 63:435–439. https://doi.org/10.1007/s00294-016-0661-8
Eleutherio E, Panek A, De Mesquita JF, Trevisol E, Magalhães R (2015) Revisiting yeast trehalose metabolism. Curr Genet 61:263–274. https://doi.org/10.1007/s00294-014-0450-1
Emri T, Majoros L, Tóth V, Pócsi I (2013) Echinocandins: production and applications. Appl Microbiol Biotechnol 97:3267–3284. https://doi.org/10.1007/s00253-013-4761-9
Ene IV, Adya AK, Wehmeier S, Brand AC, MacCallum DM, Gow NA, Brown AJ (2012) Host carbon sources modulate cell wall architecture, drug resistance and virulence in a fungal pathogen. Cell Microbiol 14:1319–1335. https://doi.org/10.1111/j.1462-5822.2012.01813.x
Enjalbert B, Nantel A, Whiteway M (2003) Stress-induced gene expression in Candida albicans: absence of a general stress response. Mol Biol Cell 14:1460–1467. https://doi.org/10.1091/mbc.e02-08-0546
Enjalbert B, Smith DA, Cornell MJ, Alam I, Nicholls S, Brown AJ, Quinn J (2006) Role of the hog1 stress-activated protein kinase in the global transcriptional response to stress in the fungal pathogen Candida albicans. Mol Biol Cell 17:1018–1032. https://doi.org/10.1091/mbc.e05-06-0501
Gasch AP (2007) Comparative genomics of the environmental stress response in ascomycete fungi. Yeast 24:961–976. https://doi.org/10.1091/mbc.e05-06-0501
Gasch AP, Spellman PT, Kao CM, Carmel-Harel O, Eisen MB, Storz G, Botstein D, Brown PO (2000) Genomic expression programs in the response of yeast cells to environmental changes. Mol Biol Cell 11:4241–4257. https://doi.org/10.1091/mbc.11.12.4241
Guan Q, Haroon S, Bravo DG, Will JL, Gasch AP (2012) Cellular memory of acquired stress resistance in Saccharomyces cerevisiae. Genetics 192:495–505. https://doi.org/10.1091/mbc.11.12.4241
Hagiwara D, Sakamoto K, Abe K, Gomi K (2016) Signaling pathways for stress responses and adaptation in Aspergillus species: stress biology in the post-genomic era. Biosci Biotechnol Biochem 80:1667–1680. https://doi.org/10.1080/09168451.2016.1162085
Hagiwara D, Takahashi H, Kusuya Y, Kawamoto S, Kamei K, Gonoi T (2016) Comparative transcriptome analysis revealing dormant conidia and germination associated genes in Aspergillus species: an essential role for AtfA in conidial dormancy. BMC Genomics 17:358. https://doi.org/10.1186/s12864-016-2689-z
Hagiwara D, Sakai K, Suzuki S, Umemura M, Nogawa T, Kato N, Osada H, Watanabe A, Kawamoto S, Gonoi T, Kamei K (2017) Temperature during conidiation affects stress tolerance, pigmentation, and trypacidin accumulation in the conidia of the airborne pathogen Aspergillus fumigatus. PLoS ONE 12:e0177050. https://doi.org/10.1371/journal.pone.0177050
Hallsworth JE (2018) Stress-free microbes lack vitality. Fungal Biol 122:379–385. https://doi.org/10.1016/j.funbio.2018.04.003
Hallsworth JE, Magan N (1995) Manipulation of intracellular glycerol and erythritol enhances germination of conidia at low water availability. Microbiology 141:1109–1115. https://doi.org/10.1099/13500872-141-5-1109
Hallsworth JE, Magan N (1996) Culture age, temperature, and pH affect the polyol and trehalose contents of fungal propagules. Appl Environ Microbiol 62:2435–2442. https://doi.org/10.1128/AEM.62.7.2435-2442.1996
Heitman J, Movva NR, Hall MN (1991) Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science 253:905–909. https://doi.org/10.1126/science.1715094
Hirota K, Miyoshi T, Kugou K, Hoffman CS, Shibata T, Ohta K (2008) Stepwise chromatin remodelling by a cascade of transcription initiation of non-coding RNAs. Nature 456:130–134. https://doi.org/10.1038/nature07348
Hohmann S, Mager WH (2003) Yeast stress responses. Topics in Current Genetics, Springer, Berlin
Hohmann S, Mager WH (2003) Introduction – What is stress? In: Hohmann S, Mager WH (eds) Yeast stress responses. Springer, Berlin, pp 1–9
Izawa S, Inoue Y, Kimura A (1996) Importance of catalase in the adaptive response to hydrogen peroxide: analysis of acatalasaemic Saccharomyces cerevisiae. Biochem J 320:61–67. https://doi.org/10.1042/bj3200061
Kaloriti D, Tillmann A, Cook E, Jacobsen M, You T, Lenardon M, Ames L, Barahona M, Chandrasekaran K, Coghill G, Goodman D, Gow NA, Grebogi C, Ho HL, Ingram P, McDonagh A, de Moura AP, Pang W, Puttnam M, Radmaneshfar E, Romano MC, Silk D, Stark J, Stumpf M, Thiel M, Thorne T, Usher J, Yin Z, Haynes K, Brown AJ (2012) Combinatorial stresses kill pathogenic Candida species. Med Mycol 50:699–709. https://doi.org/10.1042/bj3200061
Kaloriti D, Jacobsen M, Yin Z, Patterson M, Tillmann A, Smith DA, Cook E, You T, Grimm MJ, Bohovych I, Grebogi C, Segal BH, Gow NA, Haynes K, Quinn J, Brown AJ (2014) Mechanisms underlying the exquisite sensitivity of Candida albicans to combinatorial cationic and oxidative stress that enhances the potent fungicidal activity of phagocytes. mBio 5:e01334-14. https://doi.org/10.1128/mBio.01334-14
Klipp E, Nordlander B, Krüger R, Gennemark P, Hohmann S (2005) Integrative model of the response of yeast to osmotic shock. Nat Biotechnol 23:975–982. https://doi.org/10.1038/nbt1114
Kos I, Patterson MJ, Znaidi S, Kaloriti D, da Silva Dantas A, Herrero-de-Dios CM, d’Enfert C, Brown AJ, Quinn J (2016) Mechanisms underlying the delayed activation of the cap1 transcription factor in Candida albicans following combinatorial oxidative and cationic stress important for phagocytic potency. mBio 7:e00331. https://doi.org/10.1128/mBio.00331-16
Kovács R, Nagy F, Tóth Z, Bozó A, Balázs B, Majoros L (2019) Synergistic effect of nikkomycin Z with caspofungin and micafungin against Candida albicans and Candida parapsilosis biofilms. Lett Appl Microbiol 69:271–278. https://doi.org/10.1111/lam.13204
Kundu S, Horn PJ, Peterson CL (2007) SWI/SNF is required for transcriptional memory at the yeast GAL gene cluster. Genes Dev 21:997–1004. https://doi.org/10.1101/gad.1506607
Kurucz V, Krüger T, Antal K, Dietl AM, Haas H, Pócsi I, Kniemeyer O, Emri T (2018) Additional oxidative stress reroutes the global response of Aspergillus fumigatus to iron depletion. BMC Genomics 19:357. https://doi.org/10.1186/s12864-018-4730-x
Lai YW, Campbell LT, Wilkins MR, Pang CN, Chen S, Carter DA (2016) Synergy and antagonism between iron chelators and antifungal drugs in Cryptococcus. Int J Antimicrob Agents 48:388–394. https://doi.org/10.1016/j.ijantimicag.2016.06.012
Laskar SKS, Bhattacharyya MK, Nair AS, Dhar P, Bhattacharyya S (2015) Heat stress-induced cup9-dependent transcriptional regulation of SIR2. Mol Cell Biol 35:437–450. https://doi.org/10.1128/MCB.01046-142
Leach MD, Tyc KM, Brown AJ, Klipp E (2012) Modelling the regulation of thermal adaptation in Candida albicans, a major fungal pathogen of humans. PLoS One 7:e32467. https://doi.org/10.1371/journal.pone.0032467
Leong HS, Dawson K, Wirth C, Li Y, Connolly Y, Smith DL, Wilkinson CR, Miller CJ (2014) A global non-coding RNA system modulates fission yeast protein levels in response to stress. Nat Commun. 5:3947. https://doi.org/10.1038/ncomms4947
Lewis JG, Learmonth RP, Watson K (1995) Induction of heat, freezing and salt tolerance by heat and salt shock in Saccharomyces cerevisiae. Microbiology 141:687–694. https://doi.org/10.1099/13500872-141-3-687
Lima SL, Colombo AL, de Almeida Junior JN (2019) Fungal cell wall: emerging antifungals and drug resistance. Front Microbiol 10:2573. https://doi.org/10.3389/fmicb.2019.02573
Lindquist S (1986) The heat-shock response. Annu Rev Biochem 55:1151–1191. https://doi.org/10.1146/annurev.bi.55.070186.005443
Liu J, Farmer JD Jr, Lane WS, Friedman J, Weissman I, Schreiber SL (1991) Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP-FK506 complexes. Cell 66:807–815. https://doi.org/10.1016/0092-8674(91)90124-h
Lushchak VI (2010) Oxidative stress in yeast. Biochemistry (Mosc) 75:281–296. https://doi.org/10.1134/s0006297910030041
Magalhães RSS, Popova B, Braus GH, Outeiro TF, Eleutherio ECA (2018) The trehalose protective mechanism during thermal stress in Saccharomyces cerevisiae: the roles of Ath1 and Agt1. FEMS Yeast Res. https://doi.org/10.1093/femsyr/foy066
Maris AF, Assumpção AL, Bonatto D, Brendel M, Henriques JA (2001) Diauxic shift-induced stress resistance against hydroperoxides in Saccharomyces cerevisiae is not an adaptive stress response and does not depend on functional mitochondria. Curr Genet 39:137–149. https://doi.org/10.1007/s002940100194
Marr KA, Lyons CN, Rustad TR, Bowden RA, White TC (1998) Rapid, transient fluconazole resistance in Candida albicans is associated with increased mRNA levels of CDR. Antimicrob Agents Chemother 42:2584–2589. https://doi.org/10.1128/AAC.42.10.2584
Mitchell A, Romano GH, Groisman B, Yona A, Dekel E, Kupiec M, Dahan O, Pilpel Y (2009) Adaptive prediction of environmental changes by microorganisms. Nature 460:220–224. https://doi.org/10.1038/nature08112
Morano KA, Grant CM, Moye-Rowley WS (2012) The response to heat shock and oxidative stress in Saccharomyces cerevisiae. Genetics 190:1157–1195. https://doi.org/10.1534/genetics.111.128033
Nüsse O (2011) Biochemistry of the phagosome: the challenge to study a transient organelle. ScientificWorldJournal 11:2364–2381. https://doi.org/10.1100/2011/741046
Owens RA, Hammel S, Sheridan KJ, Jones GW, Doyle S (2014) A proteomic approach to investigating gene cluster expression and secondary metabolite functionality in Aspergillus fumigatus. PLoS One 9:e106942. https://doi.org/10.1371/journal.pone.0106942
Pais P, Galocha M, Teixeira MC (2019) Genome-wide response to drugs and stress in the pathogenic yeast Candida glabrata. Prog Mol Subcell Biol 58:155–193. https://doi.org/10.1007/978-3-030-13035-0_7
Plesofsky-Vig N, Tbrambl R (1985) Heat shock response of Neurospora crassa: protein synthesis and induced thermotolerance. J Bacteriol 162:1083–1091. https://doi.org/10.1128/JB.162.3.1083-1091.1985
Pócsi I, Prade RA, Penninckx MJ (2004) Glutathione, altruistic metabolite in fungi. Adv Microb Physiol 49:1–76. https://doi.org/10.1016/S0065-2911(04)49001-8
Pradhan A, Ma Q, de Assis LJ, Leaves I, Larcombe DE, Rodriguez Rondon AV, Nev OA, Brown AJP (2021) Anticipatory stress responses and immune evasion in fungal pathogens. Trends Microbiol 29:416–427. https://doi.org/10.1016/j.tim.2020.09.010
Rangel DEN, Alston DG, Roberts DW (2008) Effects of physical and nutritional stress conditions during mycelial growth on conidial germination speed, adhesion to host cuticle, and virulence of Metarhizium anisopliae, an entomopathogenic fungus. Mycol Res 112:1355–1361. https://doi.org/10.1016/j.mycres.2008.04.011
Rangel DEN, Anderson AJ, Roberts DW (2008) Evaluating physical and nutritional stress during mycelial growth as inducers of tolerance to heat and UV-B radiation in Metarhizium anisopliae conidia. Mycol Res 112:1362–1372. https://doi.org/10.1016/j.mycres.2008.04.013
Rangel DEN, Fernandes EK, Braga GU, Roberts DW (2011) Visible light during mycelial growth and conidiation of Metarhizium robertsii produces conidia with increased stress tolerance. FEMS Microbiol Lett 315:81–86. https://doi.org/10.1111/j.1574-6968.2010.02168.x
Rangel DEN, Braga GUL, Fernandes ÉKK, KeyserHallsworth CAJE, Roberts DW (2015) Stress tolerance and virulence of insect-pathogenic fungi are determined by environmental conditions during conidial formation. Current Genetics 61:383–404. https://doi.org/10.1007/s00294-015-0477-y
Rangel DEN, Finlay RD, Hallsworth JE, Dadachova E, Gadd GM (2018) Fungal strategies for dealing with environment-and agriculture-induced stresses. Fungal Biol 122:602–612. https://doi.org/10.1016/j.funbio.2018.02.002
Roetzer A, Gregori C, Jennings AM, Quintin J, Ferrandon D, Butler G, Kuchler K, Ammerer G, Schüller C (2008) Candida glabrata environmental stress response involves Saccharomyces cerevisiae Msn2/4 orthologous transcription factors. Mol Microbiol 69:603–620. https://doi.org/10.1111/j.1365-2958.2008.06301.x
Sakamoto K, Iwashita K, Yamada O, Kobayashi K, Mizuno A, Akita O, Mikami S, Shimoi H, Gomi K (2009) Aspergillus oryzae atfA controls conidial germination and stress tolerance. Fungal Genet Biol 46:887–897. https://doi.org/10.1016/j.fgb.2009.09.004
Selye H (1936) Syndrome produced by diverse nocuous agents. Nature 138:32
Selye H (1976) Stress in health and disease. Butterworth, Boston
Sugar AM, Liu XP (1998) Interactions of itraconazole with amphotericin B in the treatment of murine invasive candidiasis. J Infect Dis 177:1660–1663. https://doi.org/10.1086/515319
Święciło A (2016) Cross-stress resistance in saccharomyces cerevisiae yeast–new insight into an old phenomenon. Cell Stress Chaperones 21:187–200. https://doi.org/10.1007/s12192-016-0667-7
Tapia H, Young L, Fox D, Bertozzi CR, Koshland D (2015) Increasing intracellular trehalose is sufficient to confer desiccation tolerance to Saccharomyces cerevisiae. Proc Natl Acad Sci USA 112:6122–6127. https://doi.org/10.1073/pnas.1506415112
Thammavongs B, Denou E, Missous G, Guéguen M, Panoff JM (2008) Response to environmental stress as a global phenomenon in biology: the example of microorganisms. Microbes Environ 23:20–23. https://doi.org/10.1264/jsme2.23.20
Tóth V, Antal K, Gy Gyémánt, Miskei M, Pócsi I, Emri T (2009) Optimization of coprogen production in Neurospora crassa. Acta Biol Hung 60:321–328. https://doi.org/10.1556/ABiol.60.2009.3.9
Tóth V, Szilágyi M, Anton F, Leiter É, Pócsi I, Emri T (2013) Interactions between naturally occurring antifungal agents. Acta Biol Hung 64:519–521. https://doi.org/10.1556/ABiol.64.2013.4.11
Tribus M, Galehr J, Trojer P, Brosch G, Loidl P, Marx F, Haas H, Graessle S (2005) HdaA, a major class 2 histone deacetylase of Aspergillus nidulans, affects growth under conditions of oxidative stress. Eukaryot Cell 4:1736–1745
van Munster JM, Daly P, Delmas S, Pullan ST, Blythe MJ, Malla S, Kokolski M, Noltorp ECM, Wennberg K, Fetherston R, Beniston R, Yu X, Dupree P, Archer DB (2014) The role of carbon starvation in the induction of enzymes that degrade plant-derived carbohydrates in Aspergillus niger. Fungal Genet Biol 72:34–47. https://doi.org/10.1128/EC.4.10.1736-1745.2005
Walker LA, Gow NAR, Munro CA (2010) Fungal echinocandin resistance. Fungal Genet Biol 47:117–126. https://doi.org/10.1016/j.fgb.2009.09.003
Wambaugh MA, Denham ST, Ayala M, Brammer B, Stonhill MA, Brown JC (2020) Synergistic and antagonistic drug interactions in the treatment of systemic fungal infections. Elife 9:e54160. https://doi.org/10.7554/eLife.54160
You T, Ingram P, Jacobsen MD, Cook E, McDonagh A, Thorne T, Lenardon MD, de Moura AP, Romano MC, Thiel M, Stumpf M, Gow NA, Haynes K, Grebogi C, Stark J, Brown AJ (2012) A systems biology analysis of long and short-term memories of osmotic stress adaptation in fungi. BMC Res Notes 5:258. https://doi.org/10.1186/1756-0500-5-258
Zacharioudakis I, Gligoris T, Tzamarias D (2007) A yeast catabolic enzyme controls transcriptional memory. Curr Biol 17:2041–2046. https://doi.org/10.1186/1756-0500-5-258
Funding
Open access funding provided by University of Debrecen. The research was financed by the National Research, Development and Innovation Office (Hungary) project K131767.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Emri, T., Forgács, K. & Pócsi, I. Biologia futura: combinatorial stress responses in fungi. BIOLOGIA FUTURA 73, 207–217 (2022). https://doi.org/10.1007/s42977-022-00121-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42977-022-00121-8