
Abstract
Natural history of facultative social taxa is important for understanding the evolution of Eusociality. However, data for the social status of many species are missing. Social status of seven European Ceratina bees was studied in Cyprus. Multifemale nests were found in five species of three different subgenera: C. (Euceratina) mandibularis, C. (E.) cypriaca, C. (E.) chrysomalla, C. (Dalyatina) parvula and C. (Neoceratina) bispinosa. No social nests were found in C. (E.) dallatoreana and C. (N.) schwarzi, in which the sample size was also small. We compared nest productivity between solitary and multifemale nests in C. cypriaca, C. mandibularis, and C. parvula. It is apparent that multifemale nests have a higher number of brood cells than solitary nests. Per capita productivity was equal in both nest types. We can conclude that facultative sociality is common in Ceratina bees of subtropical climate and social nesting is the successful strategy.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Biologists were always fascinated by social insect societies (Keller and Chapuisat 2010). The origin of eusociality is one of the key puzzles of evolutionary biology (Wilson and Hölldobler 2005; Keller and Chapuisat 2010; Kennedy et al. 2017), and it was one of the problematic points for Darwin´s original theory. Although social insects are highly studied, the research has focused mainly on a few species. These species are usually highly social; therefore being useful for studying many questions concerning cooperation and conflict between individuals, however, they are unsuitable for the study of the origin of eusociality.
With their behavioral variability, facultatively eusocial species are a unique opportunity to take a look at both sides of sociality. These species are able to manifest the solitary strategy as well as the eusocial one (Schwarz et al. 2007; Shell and Rehan 2017). The largest lineages of socially polymorphic bees are Halictidae, Xylocopinae and Euglossini (Schwarz et al. 2007; Shell and Rehan 2017).
Emergence of social colonies in facultatively eusocial species can be dependent on ecological conditions, e.g. climate (Field et al. 2010; Purcell 2011). Temperate climate has often too short a season for the development of two subsequent generation of bees; therefore some species perform only solitary nesting despite retaining the ability for social nesting (Field et al. 2010). Eusocial colonies of facultatively eusocial species consist only of a few females (from two to lower tens) (Schwarz et al. 2007; Turillazzi 2013). We can compare solitary and social colonies and evaluate the success of these strategies. We might think that social colonies should have a higher productivity per female than solitary colonies, however, published evidence show that the productivity of social colonies per female is often lower than in solitary colonies (Michener 1964; Richards 2011; Rehan et al. 2014). For a proper assessment of costs and benefits of sociality, it is necessary to take the probability of nest failure into account, which is lower in social colonies (Stark 1992; Clouse 2001; Smith et al. 2007; Leadbeater et al. 2011).
Facultatively eusocial species were previously commonly considered as solitary, because sometimes it is not easy to find evidence for sociality in such species. Their frequency can be either very low in the population (Sakagami and Maeta 1987) or sociality can even occur only in some populations (Field et al. 2010) or during a limited time of a year (Dew et al. 2018; Mikát et al. 2021a).
Knowledge about the natural history of facultatively eusocial groups is essential for identifying key factors influencing the origin of eusociality. Origin of eusociality can be influenced by ecological pressures and relatedness among individuals (Keller and Chapuisat 2010; Purcell 2011; Kocher and Paxton 2014). However, the evolution of eusociality is strongly influenced by the natural history traits of each taxon (Andersson 1984; Ross et al. 2013). For example, facultatively eusocial groups with progressive provisioning such as Stenogastrinae wasps and Allodapine bees have not reversed to strictly solitary behavior (Chenoweth et al. 2007; Turillazzi 2013). On the other hand, mass provisioning groups such as Halictid bees or carpenter bees from the genera Ceratina or Xylocopa frequently reverse the strategy to strict solitarity (Rehan et al. 2012; Gibbs et al. 2012; Groom and Rehan 2018). Another example is the architecture of nests in Xylocopa. There are different tendencies in groups with linear and branched nests in Xylocopa. Species with linear nest building have the tendency to nest eusocial, but species with branched nest building have the tendency to build communal nests (Steen and Schwarz 2000). While the natural history of facultative eusocial taxa is the important knowledge for understanding the social evolution, it is still insufficient or unknown for most of the taxa. Small carpenter bees (Ceratina) are understudied in nesting and social biology compared to other facultatively eusocial species.
Small carpenter bees (Ceratina) belong to the subfamily Xylocopinae in the family Apidae and they are an exemplary taxon for the study of facultative eusociality (Sakagami and Maeta 1977; Hogendoorn and Velthuis 1999; Groom and Rehan 2018). Facultative eusociality is a primary state and the strictly solitary strategy is considered a reversion from facultative eusociality (Rehan et al. 2012). Ceratina bees nest in broken twigs. They build linear nests containing several brood cells (Michener 1990) which are mass provisioned (Rehan 2020). When provisioning is finished, the mother usually guards her offspring until adulthood (Sakagami and Maeta 1977; Rehan and Richards 2010; Mikát et al. 2016) and after that feeds mature offspring with pollen and nectar (Sakagami and Maeta 1977; Mikát et al. 2017; Rehan 2020) .
Obligatory eusocial Ceratina species do not exist. On the other hand, some reversions to strict solitarity are known (Michener 1990; Rehan and Richards 2010; Groom and Rehan 2018). Although most species are able to develop eusocial colonies, their proportion in the population is small—between 1 and 30% (Sakagami and Maeta 1984; Okazaki 1987; Rehan et al. 2010, 2015). Generally, tropical species have a higher tendency to manifest sociality than temperate species (Groom and Rehan 2018).
Social colonies are very small and usually contain only two females (Okazaki 1987; Rehan et al. 2010, 2015; Rehan 2020). Sometimes, there can be up to four residents (Sakagami and Maeta 1984; Michener 1990). More than one female in a colony can lay eggs, though usually one is reproductively dominant (Sakagami and Maeta 1984; Hogendoorn and Velthuis 1999).
There is a high diversity of Ceratina bees in the Meditarrean region with 35 species belonging to five subgenera (Terzo 1998; Terzo and Rasmont 2011). However, nesting biology and sociality in most of these species is unknown especially in comparison to North American and Japanese species (Sakagami and Maeta 1977, 1987; Vickruck et al. 2011; Lawson et al. 2018). The goal of our study was to increase our knowledge about the sociality of Meditarrean species. We performed a study in Cyprus, because eight Ceratina species live there and C. cucurbitina, which dominates in most of the Mediterranean region, is absent (Terzo and Rasmont 2011; Varnava et al. 2020).
Ceratina behavior strongly differs between different nesting phases. First, a female excavates a burrow (new founding nests), later she provisions brood cells (active brood nests) and after that she guards the offspring until adulthood (full brood nests) and lastly feeds mature offspring (full-mature and mature brood nests)(Mikát et al. 2021b). Individuals also use old nests or newly excavated burrows for overwintering (Mikát and Straka 2021). Active brood nests and full brood nests are useful for the evaluation of social status (Rehan et al. 2015). At these stages, juveniles in brood cells are present together with adults. We considered the nest social, when multiple females were present in active or full brood nest stage, consistently with previous works on Ceratina bees (Sakagami and Maeta 1977, 1984, 1987; Okazaki 1987; Rehan et al. 2009, 2015; Groom and Rehan 2018).
We discovered multifemale nests in five species and examined the frequency of multifemale nesting and the number of females in multifemale nests in each species. Moreover, we examined the difference of nest productivity and architecture between solitary and multifemale nests in three species where multifemale nests were most commonly present.
2 Methods
Study area
All samples were collected in Cyprus. Most of the samples were collected around Lefkara village (34.8744575N, 33.3113722E) and between villages Mathiatis and Agios Theodoros. However, other nests were collected in other parts of Cyprus, specifically around Paphos and Limassol. Coordinates of all sampled nests are showed in Online resource 2.
Sampling process
We sampled nests in four periods through the nesting season: 25–30 May 2018, 15–28 June 2019, 6–12 August 2019 and 17–22 September 2019. We collected nests from semi-natural nesting opportunities, such as cut or otherwise disturbed twigs next to roads. However, nests were relatively scarce in the landscape. Therefore, we cut additional twigs to increase nesting opportunities. We performed the cutting of twigs in March, June and August 2019. The most common nesting substrate was blackberry (Rubus spp.), and nests were also frequently collected from fennel (Foeniculum vulgare) and a giant fennel (Ferula communis).
Dissecting of nests
Broken twigs containing a possible nest entrance (presented as a hole in the disturbed area) of Ceratina were collected. It is not possible to determine whether the nest truly is present inside of the twig; therefore we collected all twigs which could potentially contain a nest and the presence of nest was confirmed during the dissection process. We collected nests after 6 p.m. to ensure that all inhabitants were back from foraging trips and therefore inside nests. The nest entrance was closed by a tape forthwith of collection. Nests were opened with clippers. We noted these nest features: number, sex, and age of adult individuals, number of brood cells, number and stage of development of immature offspring, presence of parasites, and nesting substrate. We measured the following parameters of nest: length of nest, length of entrance burrow, width of nest, width of pith and width of twig. In total, we sampled 20 nests of C. bispinosa Handlirsch, 338 nests of C. cypriaca Mavromoustakis, 192 nests of C. dallatoreana Friese, 131 nests of C. chrysomalla Gerstaecker, 203 nests of C. mandibularis Friese, 102 nests of C. parvula Smith and 45 nests of C. schwarzi Kocourek.
There is no existing taxonomic key for species identification of Cypriot Ceratina. We used publications from Daly (1983) and Terzo (1998) for species determination. Collected individuals were also compared to determined specimens deposited in museum in Linz, Austria.
Collected nests were in different stages: burrows, which contained only adult(s) without any sign of present or past brood cells. These burrows are either newly founded nests, burrows prepared for overwintering or temporary shelters only. However, in the case of C. dallatoreana, we distinguished burrows with nectar storage at the bottom of the nest and standard burrows without this storage. Other types of nests were active brood nests, which contained brood cells and the outermost brood cell contained egg or was recently provisioned with pollen supplies; full brood nest, which contained larva or pupa in the outermost brood cell and all brood cell partitions were preserved; full-mature brood nests, which contained immature stages and also newly enclosed adults which damaged brood cell partitions; and mature brood nests which contained only mature offspring.
Analysis of phenology
Nests which contained at least one adult female were included in this analysis. Therefore, nests with only males, juveniles or parasites were not included in the analysis because the goal of this analysis was to evaluate the behavior of females through the season. We calculated the proportion of different nest stages.
Comparison between solitary and social nests
We considered multifemale nests as social and single-female nest as solitary. We included only active brood nests and full brood nests in this analysis. We excluded orphaned nests (nests without a female), because these nests cannot be classified in any category as no female is present there. For most analyses, we analyzed a pooled dataset with those with active or full brood nests, because there was no significant difference in the number of brood cells in most species between active brood nests or full brood nests (see Results). The number of active and full brood nests together was 11 for C. bispinosa, 77 for C. chrysomalla, 164 for C. cypriaca, 65 for C. dallatoreana, 115 for C. mandibularis, 67 for C. parvula and 14 for C. schwarzi. For the analyses the number of brood cells or empty cells for were used for calculations; we excluded nests parasitized by ichneumonid or Gasteruption parasites because these parasites damaged the nest and the number of brood cells was therefore unknown. The statistical analyses were performed in R software 3.6.1. (R. Core Developmental Team 2016). Chi square tests were used for testing the proportions of social nests in active brood nests vs. full brood nests and between different parts of the season. Wilcoxon tests were used for testing the effect of sociality on number of brood cells, number of brood cells per female and length of nests, because data were not normally distributed. The effect of sociality on the proportion of empty cells was used by generalized linear model of binomial family.
Morphological characteristics of females
We measured head width as the distance between the two widest points on the head. We dissected female abdomens and extracted ovaria. Ovarian development was measured as the sum of length of the three largest oocytes (Rehan et al. 2010). Wing wear was evaluated by the relative score developed by Mueller and Wolf-Mueller (1993). This relative score recognizes seven levels of wear. When wear was between levels described by Mueller and Wolf-Mueller (1993), we evaluated it as a half-level wear. Measurements were performed on a Nikon SMZ 745 stereomicroscope.
We successfully performed measurements only for C. mandibularis, C. cypriaca and C. chrysomalla, because C. parvula and C. bispinosa were too small to successfully perform the dissection. Unfortunately, dissection was also unsuccessful in some individuals from analyzed species due to the damage to ovaria or unsuccessful location.
3 Results
3.1 Phenology
C. cypriaca
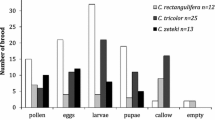
Proportion of active brood nests and full brood nests was relatively stable throughout the nesting season. The most different month was September, when the proportion of active brood nests was the lowest and the proportion of mature brood nests the highest (Figure 1).

Phenology of Ceratina species in Cyprus.
C. ballatoreana
The proportion of active brood nests was high in May (40%, 8/20) and June (54%, 15/28), it decreased in August and no active brood nests were found in September. The main peak of full brood nests was in June and August. The proportion of burrows without nectar storage was stable throughout the season. However, the proportion of burrows with nectar storage was highest in September (77%, 54/70) and relatively common in August and May. No burrows with nectar storage were found in June.
C. chrysomalla
Proportion of active brood nests was relatively stable between May and August. However, no active brood nests were found in September. The highest proportion of full brood nests was in June and September.
C. mandibularis
Proportion of active brood nests and full brood nests was stable throughout the nesting season. Mature brood nests and full-mature brood nests occurred from June, but their proportion was stable between June and September.
C. parvula
Active brood nests were very common in May (58%, 7/12) and August (68%, 15/22), and their proportion was the lowest in September (10%, 2/21). Full brood nests occurred from June to September and their proportion was highest in September.
C. bispinosa and C. schwarzi
Not enough data.
3.2 Number of brood cells provisioned
The average number of brood cells provisioned was 4.11 (N=9, SD=3.25, range 1–8) for C. bispinosa, 1.67 (N=129, SD=1.41, range 1–9) for C. cypriaca, 3.42 (N=49, SD=2.17, range 1–11) for C. dallatoreana, 4.49 (N=63, SD=2.44, range 1–11) for C. chrysomalla, 2.79 (N=97, SD=1.96, range 1–11) for C. mandibularis, 5.11 (N=65, SD=3.66, range 1–20) for C. parvula and 2.9 (N=10, SD=1.60, range 1–5) for C. schwarzi. There was no significant difference in the number of brood cells provisioned between active brood nests and full brood nests for most species (Table I). Significant difference in brood cells provisioned between active brood nests and full brood nests was in C. cypriaca (Table I), where active brood nests had a higher number of brood cells provisioned (2.50, N=40, SD=2.11, range 1–9) than full brood nests (1.30, N=89, SD=0.66, range 1– 4).
3.3 Evidence for social nesting
We found five nest categories: solitary nests (one adult female present), multifemale nests (at least two females present), nests where adult male and female were present, orphaned nest (no adult female was present) and nests where one living adult female and one dead adult female was present. Here, we note only the proportion of multifemale nests, and number of nests of all categories are summarized in Table II for each species.
C. bispinosa
Proportion of multifemale nests was 27.27% (3/11, Table II). Other nests were solitary. Two multifemale nests contained two females, one contained three females and two males. Multifemale nests were found in May, June and September.
C. cypriaca
Proportion of multifemale nests was 6.21% (10/161). There was no significant difference in the proportion of social nests between active brood nests and full brood nests (Chi square test, X^2 = 2.20, df = 1, p = 0.13, Table III). Nine multifemale nests contained two females, one contained two females and one male . Multifemale nests were found in June, August and September.
C. dallatoreana
We found only solitary or orphaned nests (N=65).
C. chrysomalla
Proportion of multifemale nests was 3.07% (2/65). Other nests were solitary. Multifemale nests contained two females.
C. mandibularis
Proportion of multifemale nests was 18.01% (20/111). There was a significant difference in the proportion of multifemale nests between active brood nests and full brood nests (Chi square test, X^2 =17.57, df = 1, p = 2.759e-05). In active brood nests, the proportion of multifemale nests was 35.29% (18/51), and in full brood nests, it was only 3.33% (2/60). Most of the multifemale nests (70%, 14/20) contained two females. However, we also found one nest with three females, four nests with four females, and one nest with five females. Multifemale nests were found in all studied periods, proportion of social nests did not significantly differ between periods (Chi square test, X^2 = 0.51, df = 3, p = 0.91).
C. parvula
Proportion of multifemale nests was 6.15% (4/65). Other nests were solitary. Two multifemale nests contained two females, one contained four females, and one contained six females. Multifemale nests were found in August and September.
C. schwarzi
We found only solitary nests (N=14).
3.4 Differences between solitary and multifemale nests
Number of brood cells
Number of brood cells provisioned was significantly higher in multifemale nests in all three species with enough nests (Wilcoxon test: C. mandibularis: W = 1273.5, p = 4.068e-06, C. cypriaca: W = 879, p = 8.603e-05, C. parvula: W = 243, p=0.0009, Figure 2, Table IV). However, the number of brood cells per female (per capita productivity) was not significantly different between solitary and social nests in any species with sufficient sample size (Wilcoxon test, C. mandibularis: W = 735.5, p = 0.75, C. cypriaca: W = 607, p = 0.43, C. parvula: W = 134.5, p = 0.74).

Comparison of productivity of solitary and social nests. Effect is significant for all three species (Wilcoxon test: C. mandibularis: W=1273.5, p=4.068e-06, C. cypriaca: W = 879, p=8.603e-05, C. parvula: W = 243, p=0.0009).
Length of nest
Social nests were significantly longer than solitary nests in C. mandibularis (Wilcoxon test, W = 1147.5, p = 0.035, Table IV) and C. parvula (Wilcoxon test, W = 212, p = 0.015), but not in C. cypriaca (Wilcoxon test, W = 937, p = 0.16).
Proportion of empty cells
Proportion of empty cells was higher in solitary nests (29%, Table IV) than in multifemale nests (17%) in C. mandibularis. The difference was statistically significant (GLM binomial, deviance = 13.53, df = 94, residual deviance = 94.11, p = 0.0002). However, this difference was not significant in C. cypriaca (GLM binomial, deviance = 0.59, df = 126, residual deviance = 105.54, p = 0.44) or C. parvula (GLM binomial, deviance = 1.05, df = 63, residual deviance = 18.75, p = 0.30).
3.5 Reproductive status of females
C. chrysomalla
We analyzed females from two social nests. Females in social nests differed in ovarian development in both nests (in the first nest, sum of the three largest oocytes was 4.63 mm for the first female and 1.53 mm for the second female, in the second nest the sum of three largest oocytes was 6.86 mm for the first female and 1.55 mm for the second female). Females with higher ovarian development had higher wing wear in both cases; however, in one nest, the larger female had a larger ovarian development and in other smaller female had larger ovarian development.
C. cypriaca
We analyzed females from seven nests, and all contained two females. The female with the larger ovarian development had the sum of the three largest oocytes 2.38 mm on average (N=7, SD = 1.89, range 0.95–6.15), the female with the smaller ovarian development had the sum of the three largest oocytes 1.22 mm (N = 7, SD = 0.66, range 0.56–2.36) on average. The difference was not statistically significant (Wilcoxon test, N = 14, W = 37, p = 0.12).
Females with larger ovarian development had larger wing wear in four cases, smaller wing wear in two cases, and in one case there was no difference in wing wear between females. Head width was not significantly different between females with larger and smaller ovarian development (paired t-test, N = 7 nests, t = 0.72, p value = 0.49).
C. mandibularis
All multifemale nests contain one female with extensive wing wear (score 1 and larger) in maximum. However, 27.77% (5/18) of nests contained only unworn females, and two other nests contained females from which one has only a slight wing wear (score 0.5).
Almost all females in social nests had at least some ovarian development. Ovaries usually differed between females in multifemale nests, although some nests contained multiple females with highly developed ovaries. The female with the most developed ovaries was usually the female with the largest wing wear. In 83.33% (10/12) of nests where females differed in wing wear, the female with the most developed ovaries had the highest wing wear. On the other hand, the largest female in the nest did not usually have the biggest ovarian development. In five nests, females did not differ in wing wear. In 29.41% (5/17) nests, the female with the largest head width also had the largest ovarian development, in 35.29% (6/17) of nests another female had the largest ovarian development and in 35.29% (6/17) of nests there was no difference in head width between females. Within nest variance in ovarian development was not related to the variance of within nest variance of wing wear (linear model, N = 17, F = 0.94, p = 0.35) nor head width (linear model, N = 17, F = 0.27, p = 0.61).
3.6 Branched nest architecture
In C. cypriaca, we found one nest (out of 338 nests) with two branches within the nest. This nest was solitary and contained brood cells in both branches. It was present in the stem of Ferula with 2.5 cm in diameter. C. cypriaca nests were frequently found in Ferula—the proportion of nests in Ferula was 68.64% (232/338). Although Ferula stems had 1.75 cm in diameter on average, we did not encounter more branched nests.
4 Discussion
4.1 Presence of sociality
Although Ceratina bees were traditionally considered as solitary and most nests are indeed solitary, the possibility of multifemale nesting was detected in multiple studied species (Michener 1974, 1990, 2007; Sakagami and Maeta 1977; Rehan et al. 2010). The most recent work reviewing the presence of sociality in the genus Ceratina (Groom and Rehan 2018) states that social nesting was present 26 out of 34 studied species. However, European species studied to this date are considered to be solitary or biparental (Mikát et al. 2016, 2019a, b). The only exception is C. chalybea, where unusual sociality with males as helpers occurs (Mikát et al. 2021a). Social biology of southern European species was mostly unexplored to this date. We examined the social strategy in seven species, and we detected social nests in five of them: C. mandibualis, C. cypriaca, C. chrysomalla, C. parvula and C. bispinosa. We have not detected social nests in two species: C. dallatoreana and C. schwarzi. However, the number of nests analyzed is too low to interpret the social status in C. schwarzi. For C. dallatoreana, the number of examined nests (56) can be insufficient if sociality is very rare, though Daly (1966) also reported C. dallatoreana as a solitary species. In our opinion, C. dallatoreana can be obligatory solitary, although more data from multiple regions would be useful.
Our results show that social nesting of Ceratina is also more common in Europe than previously presumed and probably most of the species can be facultatively eusocial. Therefore, Ceratina bees are equally important for studying the social evolution as other Xylocopinae bees such as Xylocopa or Allodapine bees or also Halictid bees (Hogendoorn and Velthuis 1999; Schwarz et al. 2007). It is interesting that the knowledge about natural history and social biology of Ceratina bees is less than in previously mentioned groups, although the collection of data is much easier.
Ceratina bees have been intensively studied mainly in temperate regions, where solitary species are common (Lawson et al. 2018; Groom and Rehan 2018), but data from the tropics showed that facultatively social species are most common in that region (Rehan et al. 2009, 2015; Groom and Rehan 2018). Cyprus, where we performed our research, lays in the subtropical zone and we showed that facultatively social species prevail there as well. Similarly, Japanese species which live in the warm-temperate or even the subtropical climate are also facultatively eusocial (Sakagami and Maeta 1977, 1987). Generally, the possibility for multiple broods per year is supposed to be an important factor for social nesting (Field et al. 2010; Purcell 2011). Ceratina bees have a long nesting season in Cyprus. Three species, C. mandibularis, C. parvula and C. cypricaca had active brood nests even in the late September, and in all these species, multifemale nests were documented. On the other hand, we have not observed any active brood nests in September in C. dallatoreana which had only solitary nests in our observation and in C. chrysomalla, which has a low proportion of social nests. The Japanese subtropical species C. okinawana in which multifemale nests occur has three peaks of reproduction per year (Okazaki 1987). Therefore, the season in subtropical climate seems to be sufficiently long for multiple broods per year.
4.2 Features of social nests
Number of individuals
Ceratina bees build linear nests; therefore females need to pass each other when one of them leaves or enters the nest. Linear nests do not allow multiple females to pass by each other easily; therefore the number of individuals per nest is constrained by nest architecture in Ceratina (Rehan et al. 2009). Two females in Ceratina nests perform an effective division of labor. One performs foraging, and the other performs guarding of the nest (Sakagami and Maeta 1984; Hogendoorn and Velthuis 1999; Rehan et al. 2010). However, more females probably cannot work effectively, because only one brood cell is provisioned at a time. Nest architecture of Ceratina nests is probably constrained by the increase in colony size in Ceratina bees more than in halictid bees, where multiple brood cells can be provisioned (Schwarz et al. 2007; Michener 2007), or in allodapine bees with progressive provisioning (Schwarz et al. 2007).
Social Ceratina nests usually contain only two females and can contain up to four females (Sakagami and Maeta 1984; Okazaki 1987; Rehan et al. 2010). Although two-female nests were the most common pattern in social nests in all social species in this study, we repeatedly detected nests with more individuals. We found even six females in a nest of C. parvula. Moreover, we found one nest with five females and four females frequently occurred in the nests of C. mandibularis. Therefore, these species can build unusually large societies in comparison to other Ceratina bees.
Males
In rare cases, we found adult males present in active brood nests and full brood nests. We observed male-female pair in C. mandibularis and C. cypariaca. Moreover, we found adult males in multifemale nests in C. cypriaca and C. bispinosa. These males can be accidental visitors and/or also male helpers which philopatrically stay in the nest from the previous nest clutch. We suppose that none of the studied species have regular biparental strategy, which is known in C. nigrolabiata (Mikát et al. 2019b). Also, male behavior is different from C. chalybea, where young males prevail in social nests (Mikát et al. 2021a). However, larger datasets, nest observation, and genetic tests are necessary for uncovering the role of males in these nests.
Nest productivity
Social nesting should be beneficial, if nesting productivity per individual increases. Surprisingly, empirical studies commonly show a decreasing productivity per individual (Michener 1964; Richards 2011; Prager 2014). Studies performed on Ceratina bees up to this date showed different results. Number of brood cells per female does not differ in C. japonica (Sakagami and Maeta 1984) but significantly decreases in social nests of C. australensis (Rehan et al. 2014; Dew et al. 2018), and C. chalybea (Mikát et al. 2021a). We found that social nests are more productive than solitary in three species with enough data, but the number of brood cells per female did not differ between solitary and multifemale nests. As per capita productivity is the same for both strategies, their coexistence is not surprising. However, the benefit of social nesting is probably higher than measured only by a number of brood cells provisioned per female, because social nests have smaller probability of nest failure (Smith et al. 2007). Therefore studies measuring the productivity primarily as a number of brood cells per female can underestimate the benefits of social nesting. Studies, which take also other factors such as brood or nest survival into account, showed commonly increasing per capita productivity in small societies (Hogendoorn and Zammit 2001). Social nests usually have a higher colony survival than solitary nests (Hogendoorn and Zammit 2001; Smith et al. 2007). In Ceratina bees, orphaned solitary nests are usually destroyed by natural enemies (Mikát et al. 2016), and social nests should have a lower probability of nest orphanage, although this phenomenon was not tested in Ceratina bees to this date.
Nest architecture
Social nests were generally longer than solitary nests, although the difference was significant only for C. parvula and C. mandibularis and not for C. cypriaca. As multifemale nests have higher productivity, it makes sense that they need more space for brood cells. There can be two reasons for this difference: higher probability of establishing social nests in larger burrows or enlarging and cleaning the burrow after the first brood clutch.
Very good nest structure feature is a distribution of empty cells in the nest. We found that empty cells are more often present in solitary nests than in social nests in C. mandibularis. Empty cells are considered to be a protection against natural enemies (Tepedino et al. 1979; Münster-Swendsen and Calabuig 2000) and more effectively guarded multifemale nests probably do not need this protection. However, we have not found a difference in empty cells proportion in C. cypriaca nor C. parvula. Empty cells are scarce in both species; therefore they probably rely on other types of nest protection.
Dominance hierarchies
In multifemale nests, usually multiple females had at least some ovarian development. Therefore, we can suppose that reproductive skew is incomplete and more females can participate in reproduction. Incomplete reproductive skew was documented for Japanese Ceratina from the subgenus Ceratinidia (Maeta and Sakagami 1995; Hogendoorn and Velthuis 1999) and it is generally common in small insect societies (Sherman et al. 1995). In contrast, in Xylocopa bees, a complete dominance of one female on reproduction was documented (Hogendoorn and Velthuis 1999). However, ovarian development itself is not a sufficient indicator of reproduction (Cini et al. 2013), and analysis of relatedness is necessary to uncover the reproductive dominance.
We found no relationship between the head width of female and the ovarian development. Therefore, we can suppose that the size is not important for establishing dominance hierarchies in these species. This is different from the situation in C. japonica, where the larger females usually have more developed ovaries (Sakagami and Maeta 1984). Our results show that reproductive dominance is probably related to age—females with higher wing wear also usually have more developed ovaries. Age-based and not size-based reproductive hierarchies are also known in Stenogastrinae wasps (Bridge and Field 2007). In Xylocopa bees, age is an important determinant of reproductive dominance, and an older female is usually dominant; however, too senescent females can be reproductive subordinates (Hogendoorn and Velthuis 1999). Size is important for establishing the dominance hierarchies in Xylocopa, when females are of the same generation (Hogendoorn and Velthuis 1999). As social nests are relatively scarce, our sample size is small for a deeper insight into reproductive hierarchies in our studied species. Later studies with a larger sample size are necessary for a better understanding of the reproductive division of labor in Ceratina bees.
4.3 Nesting biology of Ceratina in Cyprus
Nest guarding
Although Ceratina bees usually guard their nest until adulthood, facultative nest abandonment has been documented in two European Ceratina species (Mikát et al. 2016, 2019a). We found only a few full brood nests without a female in the most currently studied species. About 25% of full brood nests without female were found in C. chrysomalla and C. dallatoreana. However, we have not found a clear distinction in nest architecture between nests with mother and without mother in these species in contrast to the situation in C. chalybea and C. chalcites, where abandoned nests are closed by a filling plug that is much thicker than the usual nest partition (Mikát et al. 2016, 2019a). Therefore, we suppose that female always tries to guard her offspring until adulthood in all species here, and she does not perform facultative nest abandonments. However, further research of guarding strategy is necessary for the confirmation of obligate nest guarding for C. dallatoreana and C. chrysomalla.
Branched nests
Branched nests are common in Xylocopa bees (Gerling et al. 1983; Steen and Schwarz 2000). However, it has not been documented for Ceratina bees yet (Eardley and Daly 2007; Vickruck et al. 2011). Ceratina bees usually nest in relatively narrow twigs, where there is not enough space for a branched nest. However, C. cypriaca commonly uses broken Ferula stalks with about 2 cm in diameter where there can be enough space for a branched nest. We have documented only one branched nest yet. This situation can be either a mistake or a regular strategy of species. But this phenomenon deserves future research due to a possible influence of nesting architecture on social organization (Steen and Schwarz 2000).
5 Conclusion
We showed that facultative sociality prevails in Ceratina species in southern Europe (Cyprus). However, social nests are not common in facultatively social species, and their proportion was between 3 and 27% of nests, consistent with to what is known from previous studies about facultatively social Ceratina. However, we detected nests with 5 or 6 females, but not more than 4 females nesting together was recorded in Ceratina bees to this date. Social and solitary nesting are probably two similarly effective strategies, as per capita productivity is similar for both strategies. Usually, multiple females with developed ovaries are present in multifemale nests; therefore the reproductive division of labor is probably not complete. Reproductive dominance is probably related to the age, but not to the size of a female.
Data availability
Dataset generated during this research will be published with manuscript as supplementary material
References
Andersson M (1984) The evolution of eusociality. Annu Rev Ecol Syst 15:165–189
Bridge C, Field J (2007) Queuing for dominance: gerontocracy and queue-jumping in the hover wasp Liostenogaster flavolineata. Behav Ecol Sociobiol 61:1253–1259. https://doi.org/10.1007/s00265-007-0355-9
Chenoweth LB, Tierney SM, Smith JA, et al (2007) Social complexity in bees is not sufficient to explain lack of reversions to solitary living over long time scales. BMC Evol Biol 7:246. https://doi.org/10.1186/1471-2148-7-246
Cini A, Meconcelli S, Cervo R (2013) Ovarian indexes as indicators of reproductive investment and egg-laying activity in social insects: a comparison among methods. Insectes Sociaux 60:393–402
Clouse R (2001) Some effects of group size on the output of beginning nests of Mischocyttarus mexicanus (Hymenoptera: Vespidae). Fla Entomol 84:418
Daly HV (1983) Taxonomy and ecology of Ceratinini of North Africa and the Iberian peninsula (Hymenoptera: Apoidea). Syst Entomol 8:29–62. https://doi.org/10.1111/j.1365-3113.1983.tb00466.x
Daly HV (1966) Biological studies on Ceratina dallatorreana, an alien bee in California which reproduces by parthenogenesis (Hymenoptera: Apoidea). Ann Entomol Soc Am 59:1138–1154
Dew RM, Shell WA, Rehan SM (2018) Changes in maternal investment with climate moderate social behaviour in a facultatively social bee. Behav Ecol Sociobiol 72:69. https://doi.org/10.1007/s00265-018-2488-4
Eardley C, Daly HV (2007) Bees of the Genus Ceratina Latreille in Southern Africa (Hymenoptera, Apidae). Entomofauna, Supplement 13, 96pp
Field J, Paxton RJ, Soro A, Bridge C (2010) Cryptic plasticity underlies a major evolutionary transition. Curr Biol 20:2028–2031. https://doi.org/10.1016/j.cub.2010.10.020
Gerling D, Hurd PD, Hefetz A (1983) Comparative behavioral biology of two Middle East species of carpenter bees (Xylocopa Latreille)(Hymenoptera: Apoidea). Smithsonian Institution Press
Gibbs J, Brady SG, Kanda K, Danforth BN (2012) Phylogeny of halictine bees supports a shared origin of eusociality for Halictus and Lasioglossum (Apoidea: Anthophila: Halictidae). Mol Phylogenet Evol 65:926–939. https://doi.org/10.1016/j.ympev.2012.08.013
Groom SVC, Rehan SM (2018) Climate-mediated behavioural variability in facultatively social bees. Biol J Linn Soc 125:165–170. https://doi.org/10.1093/biolinnean/bly101
Hogendoorn K, Velthuis HHW (1999) Task allocation and reproductive skew in social mass provisioning carpenter bees in relation to age and size. Insectes Sociaux 46:198–207. https://doi.org/10.1007/s000400050135
Hogendoorn K, Zammit J (2001) Benefits of cooperative breeding through increased colony survival in an allodapine bee. Insectes Sociaux 48:392–397
Keller L, Chapuisat M (2010) Eusociality and Cooperation. In: eLS. American Cancer Society
Kennedy P, Baron G, Qiu B, et al (2017) Deconstructing superorganisms and societies to address big questions in biology. Trends Ecol Evol 32:861–872. https://doi.org/10.1016/j.tree.2017.08.004
Kocher SD, Paxton RJ (2014) Comparative methods offer powerful insights into social evolution in bees. Apidologie 45:289–305. https://doi.org/10.1007/s13592-014-0268-3
Lawson SP, Shell WA, Lombard SS, Rehan SM (2018) Climatic variation across a latitudinal gradient affect phenology and group size, but not social complexity in small carpenter bees. Insectes Sociaux 1–10
Leadbeater E, Carruthers JM, Green JP, et al (2011) Nest inheritance is the missing source of direct fitness in a primitively eusocial insect. Science 333:874–876. https://doi.org/10.1126/science.1205140
Maeta Y, Sakagami S (1995) Oophagy and egg replacement in artificially induced colonies of a basically solitary bee, Ceratina (Ceratinidia) okinawana (Hymenoptera, Anthophoridae, Xylocopinae), with a comparison of social behavior among Ceratina, Xylocopa and the Halictine bees. Jpn J Entomol 63:347–375
Michener CD (1974) The social behavior of the bees: A comparative study. Harvard University Press
Michener CD (1964) Reproductive efficiency in relation to colony size in hymenopterous societies. Insectes Sociaux 11:317–341. https://doi.org/10.1007/BF02227433
Michener CD (1990) Castes in xylocopine bees. In: Social Insects. Springer, pp 123–146
Michener CD (2007) The bees of the world, 2nd edn. The Johns Hopkins University Press
Mikát, M., Benda, D., Straka, J., 2021. Unrelated males in societies of a facultatively social bee. J. Apic. Res. 0, 1–12. https://doi.org/10.1080/00218839.2021.1994263
Mikát M, Benda D, Straka J (2019a) Maternal investment in a bee species with facultative nest guarding and males heavier than females. Ecol Entomol 44(6):823-832 https://doi.org/10.1111/een.12768
Mikát M, Černá K, Straka J (2016) Major benefits of guarding behavior in subsocial bees: implications for social evolution. Ecol Evol 6:6784–6797. https://doi.org/10.1002/ece3.2387
Mikát M, Franchino C, Rehan SM (2017) Sociodemographic variation in foraging behavior and the adaptive significance of worker production in the facultatively social small carpenter bee, Ceratina calcarata. Behav Ecol Sociobiol 71:135. https://doi.org/10.1007/s00265-017-2365-6
Mikát M, Janošík L, Černá K, et al (2019b) Polyandrous bee provides extended offspring care biparentally as an alternative to monandry based eusociality. Proc Natl Acad Sci 116:6238–6243. https://doi.org/10.1073/pnas.1810092116
Mikát M, Matoušková E, Straka J (2021) Nesting of Ceratina nigrolabiata , a biparental bee. Sci Rep 11:5026. https://doi.org/10.1038/s41598-021-83940-4
Mikát M, Straka J (2021) Overwintering strategy and longevity of European small carpenter bees (Ceratina). J Ethol. https://doi.org/10.1007/s10164-020-00688-6
Mueller UG, Wolf-Mueller B (1993) A method for estimating the age of bees: age-dependent wing wear and coloration in the wool-carder bee Anthidium manicatum (Hymenoptera: Megachilidae). J Insect Behav 6:529–537
Münster-Swendsen M, Calabuig I (2000) Interaction between the solitary bee Chelostoma florisomne and its nest parasite Sapyga clavicornis- empty cells reduce the impact of parasites. Ecol Entomol 25:63–70
Okazaki K (1987) Life cycle of a subtropical xylocopine bee, Ceratina okinawana, with some related problems. Kontyu 55:1–8
Prager SM (2014) Comparison of social and solitary nesting carpenter bees in sympatry reveals no advantage to social nesting. Biol J Linn Soc 113:998–1010
Purcell J (2011) Geographic patterns in the distribution of social systems in terrestrial arthropods. Biol Rev 86:475–491
R Core Team, 2016. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0
Rehan, Richards MH, Schwarz MP (2010) Social polymorphism in the Australian small carpenter bee, Ceratina (Neoceratina) australensis. Insectes Sociaux 57:403–412. https://doi.org/10.1007/s00040-010-0097-y
Rehan SM (2020) Small Carpenter Bees (Ceratina). In: Starr CK (ed) Encyclopedia of Social Insects. Springer International Publishing, Cham, pp 1–4
Rehan SM, Leys R, Schwarz MP (2012) A mid-cretaceous origin of sociality in xylocopine bees with only two origins of true worker castes indicates severe barriers to eusociality. PLoS ONE 7:e34690. https://doi.org/10.1371/journal.pone.0034690
Rehan SM, Richards MH (2010) Nesting biology and subsociality in Ceratina calcarata (Hymenoptera: Apidae). Can Entomol 142:65–74. https://doi.org/10.4039/n09-056
Rehan SM, Richards MH, Adams M, Schwarz MP (2014) The costs and benefits of sociality in a facultatively social bee. Anim Behav 97:77–85. https://doi.org/10.1016/j.anbehav.2014.08.021
Rehan SM, Richards MH, Schwarz MP (2009) Evidence of social nesting in the Ceratina of Borneo (Hymenoptera: Apidae). J Kans Entomol Soc 82:194–209. https://doi.org/10.2317/JKES809.22.1
Rehan SM, Tierney SM, Wcislo WT (2015) Evidence for social nesting in Neotropical ceratinine bees. Insectes Sociaux 62:465–469
Richards MH (2011) Colony social organisation and alternative social strategies in the eastern carpenter bee, Xylocopa virginica. J Insect Behav 24:399–411. https://doi.org/10.1007/s10905-011-9265-9
Ross L, Gardner A, Hardy N, West SA (2013) Ecology, not the genetics of sex determination, determines who helps in eusocial populations. Curr Biol 23:2383–2387
Sakagami SF, Maeta Y (1987) Multifemale nests and rudimentary castes of an" almost" solitary bee Ceratina flavipes, with additional observations on multifemale nests of Ceratina japonica (Hymenoptera, Apoidea). Kontyu Tokyo 55:391–409
Sakagami SF, Maeta Y (1977) Some presumably presocial habits of Japanese Ceratina bees, with notes on various social types in Hymenoptera. Insectes Sociaux 24:319–343. https://doi.org/10.1007/BF02223784
Sakagami SF, Maeta Y (1984) Multifemale nests and rudimentary castes in the normally solitary bee Ceratina japonica (Hymenoptera: Xylocopinae). J Kans Entomol Soc 57:639–656. https://doi.org/10.2307/25084573
Schwarz MP, Richards MH, Danforth BN (2007) Changing paradigms in insect social evolution: Insights from Halictine and Allodapine bees. Annu Rev Entomol 52:127–150. https://doi.org/10.1146/annurev.ento.51.110104.150950
Shell WA, Rehan SM (2017) Behavioral and genetic mechanisms of social evolution: insights from incipiently and facultatively social bees. Apidologie 1–18
Sherman PW, Lacey EA, Reeve HK, Keller L (1995) The eusociality continuum. Behav Ecol 6:102–108
Smith AR, Wcislo WT, O’Donnell S (2007) Survival and productivity benefits to social nesting in the sweat bee Megalopta genalis (Hymenoptera: Halictidae). Behav Ecol Sociobiol 61:1111–1120. https://doi.org/10.1007/s00265-006-0344-4
Stark RE (1992) Cooperative nesting in the multivoltine large carpenter bee Xylocopa sulcatipes Maa (Apoidea: Anthophoridae): Do helpers gain or lose to solitary females? Ethology 91:301–310. https://doi.org/10.1111/j.1439-0310.1992.tb00871.x
Steen Z, Schwarz MP (2000) Nesting and life cycle of the Australian green carpenter bees Xylocopa (Lestis) aeratus Smith and Xylocopa (Lestis) bombylans (Fabricius)(Hymenoptera: Apidae: Xylocopinae). Aust J Entomol 39:291–300
Tepedino VJ, McDonald LL, Rothwell R (1979) Defense against parasitization in mud-nesting Hymenoptera: Can empty cells increase net reproductive output? Behav Ecol Sociobiol 6:99–104. https://doi.org/10.1007/BF00292555
Terzo M (1998) Annotated list of the species of the genus Ceratina (Latreille) occuring in the Near East, with descriptions of new species (Hymenoptera: Apoidea: Xylocopinae). Linz Biol Beitr 30:719–743
Terzo M, Rasmont P (2011) Atlas of the European bees: genus Ceratina. In: Atlas Hymenopt. - Atlas Eur. Bees - STEP Proj. http://www.atlashymenoptera.net/page.asp?id=192. Accessed 25 Feb 2019
Turillazzi S (2013) The biology of hover wasps. Springer Science & Business Media
Varnava AI, Roberts SPM, Michez D, et al (2020) The wild bees (Hymenoptera, Apoidea) of the island of Cyprus. ZooKeys 924: 1–114
Vickruck JL, Rehan SM, Sheffield CS, Richards MH (2011) Nesting biology and DNA barcode analysis of Ceratina dupla and C. mikmaqi, and Comparisons with C. calcarata (Hymenoptera: Apidae: Xylocopinae). Can Entomol 143:254–262
Wilson EO, Hölldobler B (2005) Eusociality: Origin and consequences. Proc Natl Acad Sci U S A 102:13367–13371. https://doi.org/10.1073/pnas.0505858102
Acknowledgments
We are grateful to all people who helped us in nest processing or preparing nesting opportunities: Slavomír Dobrotka, Zuzana Dobrotková, Karolína Fazekašová, Jiří Houska, Jiří Janoušek, Vít Procházka, Daniela Reiterová, Vojtěch Waldhauser and Jitka Waldhauserová. We thank Androulla Varnava for advice about local conditions and discussions about bees in Cyprus.
Funding
The Grant Agency of Charles University (Grant GAUK 764119/2019) and the Specific University Research Project Integrative Animal Biology (Grant SVV 260571/2021) supported this research.
Author information
Authors and Affiliations
Contributions
Michael Mikát designed research, Michael Mikát, Daniel Benda and Tereza Fraňková performed research, Michael Mikát analyzed data, Michael Mikát wrote first draft of manuscript and Jakub Straka performed substantial changes in manuscript. All authors intensively discussed about scientific issues in manuscript and approved final version.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Ethics approval
Cyprus Ministry of Agriculture, rural development and environment permitted the research
Consent to participate
Not applicable
Consent for publication
Not applicable.
Additional information
Manuscript editor: Zachary Huang
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Mikát, M., Fraňková, T., Benda, D. et al. Evidence of sociality in European small Carpenter bees (Ceratina). Apidologie 53, 18 (2022). https://doi.org/10.1007/s13592-022-00931-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13592-022-00931-8