
Abstract
Analyzing the roles of ecology and geography on speciation and lineage diversification can shed light on the processes that generate and maintain biodiversity. Additionally, lineages rapidly diversifying across unstable habitats provide substantial challenges for resolving evolutionary histories and delimiting species. Physaria is represented in South American by six species distributed from southern Bolivia to northern central Argentina and growing in highlands of the southern-central Andes, but also along the hills and lowlands of central-eastern Argentina. This biogeographical variability, not common among other South American crucifers (Brassicaceae), prompted us to conduct different climatic niche and geographical range comparisons to study the potential roles of ecology and geography through the diversification of the group. However, the remarkable similarity between these species, coupled with the continuous variability of the diagnostic morphological characters, blurs the species boundaries. Therefore, in order to identify independent evolving lineages, we first employed species delimitation methods together with the general lineage concept of species, and used molecular sequences from nuclear ribosomal and chloroplast loci. Secondly, and in the light of the lineages obtained, we explored the roles of geography and ecology on the diversification of South American Physaria and tested for presence of phylogenetic niche-conservatism or niche-divergence patterns, as well as potential ecological speciation. Lineages identified by these delimitation methods were highly congruent with described species; nevertheless, some morphospecies were included under the same independent evolutionary lineage. Our results suggest that the climatic niche divergence along the heterogeneous landscape apparently was a major factor promoting diversification of the South American Physaria. Divergence was registered mainly on the temperature dimension, which promoted shifts between cold-temperate habitats associated with the highlands of the central-southern Andes and warm lowlands from central-eastern Argentina, i.e., the Monte and Dry Chaco ecoregions. In addition, some degree of niche divergence along the precipitation gradient was also secondarily recovered. Allopatry and dispersal capabilities also seem to be associated with the diversification of the group, presumably through the Late Pliocene-Pleistocene, and promoted by glacial cycles and climatic oscillations during the Quaternary. Results of these analyses are also discussed in a general context, which will contribute to the understanding of the evolutionary and ecological patterns of South American Brassicaceae.








Similar content being viewed by others

Data availability
Data generated or analyzed in this paper are included in the electronic supplementary material, the TreeBase repository (http://purl.org/phylo/treebase/phylows/study/TB2:S26615), and the Figshare repository (https://doi.org/10.6084/m9.figshare.12645218). New DNA sequences generated for this study are available in the GenBank repository (see corresponding GenBank numbers and vouchers in Appendix 1).
References
Ab’saber, A. N. (2000). Spaces occupied by the expansion of dry climates in South America during the Quaternary ice ages. Revista do Instituto Geológico, 21(1–2), 71–78. https://doi.org/10.5935/0100-929X.20000006.
Ackerly, D. (2003). Community assembly, niche conservatism, and adaptative evolution in changing environments. International Journal of Plant Sciences, 164, S165–S184. https://doi.org/10.1086/368401.
Agapow, P. M., Bininda-Emonds, O. R., Crandall, K. A., Gittleman, J. L., Mace, G. M., Marshall, J. C., & Purvis, A. (2004). The impact of species concept on biodiversity studies. The Quarterly Review of Biology, 79(2), 161–179. https://doi.org/10.1086/383542.
Al-Shehbaz, I. A. (2008). Brassicaceae. In F. O. Zuloaga, O. Morrone, & M. J. Belgrano (Eds.), Catalogue of the vascular plants of the southern cone (Argentina, southern Brazil, Chile, Paraguay and Uruguay), Dicotyledoneae: Acanthaceae-Fabaceae (Abarema-Schizolobium) (Vol. 2, pp. 1663–1709). Missouri: Missouri Botanical Garden Press.
Al-Shehbaz, I. A. (2012a). A generic and tribal synopsis of the Brassicaceae (Cruciferae). Taxon, 61, 931–954. https://doi.org/10.1002/tax.615002.
Al-Shehbaz, I. A. (2012b). Brassicaceae. In M. A. Anton & F. O. Zuloaga (Eds.), Flora Argentina (Vol. 8, pp. 1–270). Buenos Aires: Editorial Sigma.
Al-Shehbaz, I. A., & O’Kane Jr., S. L. (2002). Lesquerella is united with Physaria (Brassicaceae). Novon, 12(3), 319–329.
Al-Shehbaz, I. A., & Prina, A. O. (2009). Physaria okanensis (Brassicaceae), a new species from Catamarca, Argentina. Novon, 19(3), 284–285.
Anacker, B. L., & Strauss, S. Y. (2014). The geography and ecology of plant speciation: range overlap and niche divergence in sister species. Proceedings of the Royal Society of London B: Biological Sciences, 281(1778), 20132980. https://doi.org/10.1098/rspb.2013.2980.
Ané, C., Larget, B., Baum, D. A., Smith, S. D., & Rokas, A. (2007). Bayesian estimation of concordance among gene trees. Molecular Biology and Evolution, 24, 412–426. https://doi.org/10.1093/molbev/msl170.
Antonelli, A., & Sanmartín, I. (2011). Why are there so many plant species in the Neotropics? Taxon, 60(2), 403–414. https://doi.org/10.1126/science.1194585.
Antonelli, A., Nylander, J. A., Persson, C., & Sanmartín, I. (2009). Tracing the impact of the Andean uplift on Neotropical plant evolution. Proceedings of the National Academy of Sciences, 106(24), 9749–9754. https://doi.org/10.1073/pnas.0811421106.
Arana, M. D., Martinez, G. A., Oggero, A. J., Natale, E. S., & Morrone, J. J. (2017). Map and shapefile of the biogeographic provinces of Argentina. Zootaxa, 4341(3), 420–422. https://doi.org/10.11646/zootaxa.4341.3.6.
Arrigoni, R., Berumen, M. L., Chen, C. A., Terraneo, T. I., Baird, A. H., Payri, C., & Benzoni, F. (2016). Species delimitation in the reef coral genera Echinophyllia and Oxypora (Scleractinia, Lobophylliidae) with a description of two new species. Molecular Phylogenetic and Evolution, 105, 146–159. https://doi.org/10.1016/j.ympev.2016.08.023.
Baldwin, B. G. (1992). Phylogenetic utility of the internal transcribed spacers of nuclear ribosomal DNA in plants: an example from the Compositae. Molecular Phylogenetics and Evolution, 1, 3–16. https://doi.org/10.1016/10557903(92)90030-K.
Baldwin, B. G. (2005). Origin of the serpentine-endemic herb Layia discoidea from the widespread L. glandulosa (Compositae). Evolution, 59, 2473–2479. https://doi.org/10.1554/05-147.1.
Barraclough, T. G., & Vogler, A. P. (2000). Detecting the geographical pattern of speciation from species-level phylogenies. The American Naturalist, 155(4), 419–434. https://doi.org/10.1086/303332.
Barton, N. H., & Charlesworth, B. (1984). Genetic revolutions, founder effects, and speciation. Annual Review of Ecology, Evolution, and Systematics, 15, 133–164. https://doi.org/10.1146/annurev.es.15.110184.001025.
Baum, D. A. (2007). Concordance trees, concordance factors, and the exploration of reticulate genealogy. Taxon, 56, 417–426. https://doi.org/10.1002/tax.562013.
Bivand R, & Lewin-Koh, N. (2019). maptools: tools for handling spatial objects. R package version 0.9-5. https://CRAN.R-project.org/package=maptools. Accessed March 2019
Bivand, R. S., Pebesma, E., & Gomez-Rubio, V. (2013). Applied spatial data analysis with R (2nd ed.). New York: Springer.
Blisniuk, P. M., Stern, L. A., Chamberlain, C. P., Idleman, B., & Zeitler, P. K. (2005). Climatic and ecologic changes during Miocene surface uplift in the southern Patagonian Andes. Earth and Planetary Science Letters, 230, 125–142. https://doi.org/10.1016/j.Epsl.2004.11.015.
Blonder, B. (2018). Hypervolume concepts in niche-and trait-based ecology. Ecography, 41, 1441–1455. https://doi.org/10.1111/ecog.03187.
Blonder, B., & Harris, D. J. (2018). hypervolume: high dimensional geometry and set operations using kernel density estimation, support vector machines, and convex hulls. R package version 2.0.11. https://CRAN.R-project.org/package=hypervolume. Accessed March 2019
Blonder, B., Lamanna, C., Violle, C., & Enquist, B. J. (2014). The n-dimensional hypervolume. Global Ecology and Biogeography, 23, 595–609. https://doi.org/10.1111/geb.12146.
Boelcke, O. (1967). Cruciferae. In A. L. Cabrera (Ed.), Flora de la Provincia de Buenos Aires, Vol. 4, pt. 3 (pp. 281–371). Buenos Aires: Instituto Nacional de Tecnología Agropecuaria (INTA).
Boelcke, O., & Romanezuk, C. (1984). Cruciferae. In M. N. Correa (Ed.), Flora Patagonica (Vol. 4A, pp. 373–544). Buenos Aires: Instituto Nacional de Tecnología Agropecuaria (INTA).
Bowman, A. W. & Azzalini, A. (2018). R package ‘sm’: nonparametric smoothing methods (version 2.2-5.6). http://www.stats.gla.ac.uk/~adrian/sm. Accessed November 2019
Broennimann, O., Fitzpatrick, M. C., Pearman, P. B., Petitpierre, B., Pellissier, L., Yoccoz, N. G., Thuiller, W., Fortin, M., Randin, C., Zimmermann, N. E., Graham, C. H., & Guisan, A. (2012). Measuring ecological niche overlap from occurrence and spatial environmental data. Global Ecology and Biogeography, 21, 481–497. https://doi.org/10.1111/j.1466-8238.2011.00698.x.
Brown, J. L., & Carnaval, A. C. (2019). A tale of two niches: methods, concepts, and evolution. Frontiers of Biogeography, 11(4), e44158. https://doi.org/10.21425/F5FBG44158.
Cabrera, A. L. (1976). Regiones fitogeográficas Argentinas. Enciclopedia Argentina de Agricultura y Jardinería (Vol. II, Second ed.). Buenos Aires: Editorial ACME.
Cabrera, A., & Willink, A. (1973). Biogeografía de América Latina. Washington: Monografías OEA.
Calenge, C. (2006). The package adehabitat for the R software: a tool for the analysis of space and habitat use by animals. Ecological Modelling, 197, 516–519. https://doi.org/10.1016/j.ecolmodel.2006.03.017.
Carstens, B. C., Pelletier, T. A., Reid, N. M., & Satler, J. D. (2013). How to fail at species delimitation. Molecular Ecology, 22(17), 4369–4383. https://doi.org/10.1111/mec.12413.
Chan, K. O., & Grismer, L. L. (2019). To split or not to split? Multilocus phylogeny and molecular species delimitation of southeast Asian toads (family: Bufonidae). BMC Evolutionary Biology, 19(1), 95. https://doi.org/10.1186/s12862-019-1422-3.
Chesser, R. T., & Zink, R. M. (1994). Modes of speciation in birds: a test of Lynch’s method. Evolution, 48(2), 490–497. https://doi.org/10.1111/j.1558-5646.1994.tb01326.x.
Clapperton, C. M. (1993). Nature of environmental changes in South America at the Last Glacial Maximum. Palaeogeography, Palaeoclimatology, Palaeoecology, 101(3–4), 189–208. https://doi.org/10.1016/0031-0182(93)90012-8.
Clark, P. U., Dyke, A. S., Shakun, J. D., Carlson, A. E., Clark, J., Wohlfarth, B., Mitrovica, J. X., Hostetler, S. W., & McCabe, A. M. (2009). The Last Glacial Maximum. Science, 325, 710–714. https://doi.org/10.1126/science.1172873.
Collevatti, R. G., Lima, N. E., & Vitorino, L. C. (2020). The diversification of extant angiosperms in the South America dry diagonal. In V. Rull & A. C. Carnaval (Eds.), Neotropical Diversification: Patterns and Processes (pp. 547–568). Cham: Springer Nature.
Coyne, J. A. (1992). Genetics and speciation. Nature, 355, 511–515. https://doi.org/10.1038/355511a0.
Coyne, J. A., & Orr, A. H. (2004). Speciation. Massachusetts: Ed. Sunderland, Sinauer Associates, Inc..
Crandall, K. A., Bininda-Emonds, O. R., Mace, G. M., & Wayne, R. K. (2000). Considering evolutionary processes in conservation biology. Trends in Ecology & Evolution, 15(7), 290–295. https://doi.org/10.1016/S0169-5347(00)01876-0.
Darriba, D., Taboada, G. L., Doallo, R., & Posada, D. (2012). jModelTest2: more models, new heuristics and parallel computing. Nature Methods, 9, 772–772. https://doi.org/10.1038/nmeth.2109.
Dayrat, B. (2005). Towards integrative taxonomy. Biological Journal of the Linnean Society, 85(3), 407–417. https://doi.org/10.1111/j.1095-8312.2005.00503.x.
De Queiroz, K. (1998). The general lineage concept of species, species criteria, and the process of speciation and terminological recommendations. In D. J. Howard & H. S. Berlocher (Eds.), Endless Forms: Species and Speciation (pp. 57–75). Oxford: Oxford University Press.
De Queiroz, K. (1999). The general lineage concept of species and the defining properties of the species. In R. Wilson (Ed.), Species: new interdisciplinary essays (pp. 49–89). Cambridge, MA: MIT Press.
De Queiroz, K. (2005). Different species problems and their resolution. BioEssays, 27(12), 1263–1269. https://doi.org/10.1002/bies.20325.
De Queiroz, K. (2007). Species concepts and species delimitation. Systematic Biology, 56(6), 879–886. https://doi.org/10.1080/10635150701701083.
Del Río, C. J., Griffin, M., McArthur, J. M., Martínez, S., & Thirlwall, M. F. (2013). Evidence for early Pliocene and late Miocene transgressions in southern Patagonia (Argentina): 87Sr/86Sr ages of the pectinid “Chlamys” actinodes (Sowerby). Journal of South American Earth Sciences, 47, 220–229. https://doi.org/10.1016/j.jsames.2013.08.004.
Doyle, J. J., & Doyle, J. L. (1987). A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin, 19, 11–15.
Drummond, A. J., Suchard, M. A., Xie, D., & Rambaut, A. (2012). Bayesian phylogenetics with BEAUti and the BEAST 1.7. Molecular Biology and Evolution, 29, 1969–1973. https://doi.org/10.1093/molbev/mss075.
Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research, 32, 1792–1797. https://doi.org/10.1093/nar/gkh340.
Encinas, A., Zambrano, P. A., Finger, K. L., Valencia, V., Buatois, L. A., & Duhart, P. (2013). Implications of deep-marine Miocene deposits on the evolution of the north Patagonian Andes. The Journal of Geology, 121, 215–238. https://doi.org/10.1086/66997.
Ezard, T., Fujisawa, T., & Barraclough, T. G. (2009). Splits: species’ limits by threshold statistics. R package version, 1.0-19 1(11), r29. http://r-forge.r-project.org/projects/splits/. Accessed January 2020
Federici, S., Galimberti, A., Bartolucci, F., Bruni, I., De Mattia, F., Cortis, P., & Labra, M. (2013). DNA barcoding to analyse taxonomically complex groups in plants: the case of Thymus (Lamiaceae). Botanical Journal of the Linnean Society, 171(4), 687–699. https://doi.org/10.1111/boj.12034.
Fitzpatrick, B. M., & Turelli, M. (2006). The geography of mammalian speciation: mixed signals from phylogenies and range maps. Evolution, 60(3), 601–615. https://doi.org/10.1554/05-453.1.
Flouri, T., Jiao, X., Rannala, B., & Yang, Z. (2018). Species tree inference with BPP using genomic sequences and the multispecies coalescent. Molecular Biology and Evolution, 35(10), 2585–2593. https://doi.org/10.1093/molbev/msy147.
Fujisawa, T., & Barraclough, T. G. (2013). Delimiting species using single-locus data and the Generalized Mixed Yule Coalescent approach: a revised method and evaluation on simulated data sets. Systematic Biology, 62(5), 707–724. https://doi.org/10.1093/sysbio/syt033.
Futuyma, D. J., & Kirkpatrick, M. (2017). Evolution (4th ed.). Sunderland, MA: Sinauer Associates.
Garzione, C. N., Hoke, G. D., Libarkin, J. C., Withers, S., MacFadden, B., Eiler, J., Ghosh, P., & Mulch, A. (2008). Rise of the Andes. Science, 320, 1304–1307. https://doi.org/10.1126/science.1148615.
Gaston, K. J., Rodrigues, A. S. L., Van Rensburg, B. J., Koleff, P., & Chown, S. L. (2001). Complementary representation and zones of ecological transition. Ecology Letter, 4, 4–9. https://doi.org/10.1046/j.1461-0248.2001.00196.x.
Givnish, T. J. (2010). Ecology of plant speciation. Taxon, 59(5), 1326–1366.
Godoy-Bürki, A. C., Ortega-Baes, P., Sajama, J. M., & Aagesen, L. (2014). Conservation priorities in the Southern Central Andes: mismatch between endemism and diversity hotspots in the regional flora. Biodiversity and conservation, 23(1), 81–107. https://doi.org/10.1007/s10531-013-0586-1.
Graham, A. (2009). The Andes: a geological overview from a biological perspective. Annals of the Missouri Botanical Garden, 96, 371–385. https://doi.org/10.3417/2007146.
Graham, A., Gregory-Wodzicki, K. M., & Wright, K. L. (2001). Studies in Neotropical paleobotany. XV. A Mio-Pliocene palynoflora from the Eastern Cordillera, Bolivia: Implications for the uplift history of the Central Andes. American Journal of Botany, 88, 1545–1557. https://doi.org/10.2307/3558398.
Gregory-Wodzicki, K. M. (2000). Uplift history of the Central and Northern Andes: a review. Geological Society of America Bulletin, 112, 1091–1105. https://doi.org/10.1130/00167606(2000)112%3C1091:UHOTCA%3E2.0.CO;2.
Grossenbacher, D. L., Veloz, S. D., & Sexton, J. P. (2014). Niche and range size patterns suggest that speciation begins in small, ecologically diverged populations in North American monkey-flowers (Mimulus spp.). Evolution, 68(5), 1270–1280. https://doi.org/10.1111/evo.12355.
Grummer, J. A., Bryson Jr., R. W., & Reeder, T. W. (2014). Species delimitation using Bayes factors: simulations and application to the Sceloporus scalaris species group (Squamata: Phrynosomatidae). Systematic Biology, 63(2), 119–133. https://doi.org/10.1093/sysbio/syt069.
Hall, T. A. (1999). BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series, 41, 95–98.
Hamilton, M. B. (1999). Four primer pairs for the amplification of chloroplast intergenic regions with intraspecific variation. Molecular Ecology, 8, 521–523.
Hazzi, N. A., Moreno, J. S., Ortiz-Movliav, C., & Palacio, R. D. (2018). Biogeographic regions and events of isolation and diversification of the endemic biota of the tropical Andes. Proceedings of the National Academy of Sciences of United States of America, 115(31), 7985–7990. https://doi.org/10.1073/pnas.1803908115.
Heled, J., & Drummond, A. J. (2010). Bayesian inference of species trees from multilocus data. Molecular Biology and Evolution, 27, 570–580. https://doi.org/10.1093/molbev/msp274.
Hewitt, G. M. (2004). Genetic consequences of climatic oscillations in the Quaternary. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 359(1442), 183–195. https://doi.org/10.1098/rstb.2003.1388.
Hijmans, R. J. (2019). raster: geographic data analysis and modeling. R package version 2.8-19. https://CRAN.R-project.org/package=raster. Accessed March 2019
Hoorn, C., Wesselingh, F. P., Steege, T., Bermudez, H., Mora, M. A., Sevink, A., Sanmartín, I., Sanchez-Meseguer, A., Anderson, C. L., Figueiredo, J. P., Jaramillo, C., Riff, D., Negri, F. R., Hooghiemstra, H., Lundberg, J., Stadler, T., Särkinen, T., & Antonelli, A. (2010). Amazonia through time: Andean uplift, climate change, landscape evolution, and biodiversity. Science, 330, 927–931. https://doi.org/10.1126/science.1194585.
Huang, X. C., German, D. A., & Koch, M. A. (2020). Temporal patterns of diversification in Brassicaceae demonstrate decoupling of rate shifts and mesopolyploidization events. Annals of botany, 125(1), 29–47. https://doi.org/10.1093/aob/mcz123.
Huson, D. H., & Bryant, D. (2006). Application of phylogenetic networks in evolutionary studies. Molecular Biology and Evolution, 23, 254–267. https://doi.org/10.1093/molbev/msj030.
Huson, D. H., Dezulian, T., Klopper, T., & Steel, M. A. (2004). Phylogenetic super-networks from partial trees. IEEE/ACM Transactions on Computational Biology and Bioinformatics, 1, 151–158.
Hutchinson, G. (1957). Concluding remarks. Cold Spring Harbor Symposia on Quantitative Biology, 22, 415–427. https://doi.org/10.1101/SQB.1957.022.01.039.
Isaac, N. J., Mallet, J., & Mace, G. M. (2004). Taxonomic inflation: its influence on macroecology and conservation. Trends in Ecology & Evolution, 19(9), 464–469. https://doi.org/10.1016/j.tree.2004.06.004.
Jackson, D. A. (1993). Stopping rules in principal components analysis: a comparison of heuristical and statistical approaches. Ecology, 74, 2204–2214.
Jackson, N. D., Carstens, B. C., Morales, A. E., & O’Meara, B. C. (2017). Species delimitation with gene flow. Systematic Biology, 66(5), 799–812. https://doi.org/10.1093/sysbio/syw117.
Jezkova, T., & Wiens, J. J. (2018). Testing the role of climate in speciation: new methods and applications to squamate reptiles (lizards and snakes). Molecular Ecology, 27(12), 2754–2769. https://doi.org/10.1111/mec.14717.
Jordan, D. S. (1905). The origin of species through isolation. Science, 22(566), 545–562.
Jordan, T. E., Burns, W. M., Veiga, R., Pangaro, F., Copeland, P., Kelley, S., & Mpodozis, C. (2001). Extension and basin formation in the Southern Andes caused by increased convergence rate: a mid-Cenozoic trigger for the Andes. Tectonics, 20, 308–324. https://doi.org/10.1029/1999TC001181.
Karger, D. N., Conrad, O., Böhner, J., Kawohl, T., Kreft, H., Soria-Auza, R. W., Zimmermann, N. E., & Kessler, M. (2017a). Climatologies at high resolution for the earth’s land surface areas. Scientific data, 4, 170122. https://doi.org/10.1038/sdata.2017.122.
Karger, D. N., Conrad, O., Böhner, J., Kawohl, T., Kreft, H., Soria-Auza, R. W., Zimmermann, N. E., Linder, H. P., & Kessler, M. (2017b). Data from: climatologies at high resolution for the earth’s land surface areas. Dryad Digital Repository. https://doi.org/10.5061/dryad.kd1d4.
Kass, R. E., & Raftery, A. E. (1995). Bayes factors. Journal of the American Statistical Association, 90, 773–795.
Kindt, R., & Coe, R. (2005). In World Agroforestry Centre (ICRAF) (Ed.), Tree diversity analysis. A manual and software for common statistical methods for ecological and biodiversity studies. Nairobi: http://www.worldagroforestry.org/output/tree-diversity-analysis. Accessed January 2020
Kisel, Y., & Barraclough, T. G. (2010). Speciation has a spatial scale that depends on levels of gene flow. The American Naturalist, 175(3), 316–334. https://doi.org/10.1086/650369.
Koch, M. A. (2015). A new chromosome was born: comparative chromosome painting in Boechera. Trends in Plant Science, 20(9), 533–535. https://doi.org/10.1016/j.tplants.2015.07.001.
Koch, M. A., & Grosser, J. (2017). East Asian Arabis species (Brassicaceae) exemplify past hybridization and subsequent emergence of three main evolutionary lineages in East Asia, America and the amphi-Beringian region. Botanical Journal of the Linnean Society, 184(2), 224–237. https://doi.org/10.1093/botlinnean/box020.
Kolanowska, M., Grochocka, E., & Konowalik, K. (2017). Phylogenetic climatic niche conservatism and evolution of climatic suitability in Neotropical Angraecinae (Vandeae, Orchidaceae) and their closest African relatives. PeerJ, 5, e3328. https://doi.org/10.7717/peerj.3328.
Kolář, F., Dušková, E., & Sklenář, P. (2016). Niche shifts and range expansions along cordilleras drove diversification in a high-elevation endemic plant genus in the tropical Andes. Molecular ecology, 25(18), 4593–4610. https://doi.org/10.1111/mec.13788.
Kozak, K. H., & Wiens, J. J. (2006). Does niche conservatism promote speciation? A case study in North American salamanders. Evolution, 60(12), 2604–2621. https://doi.org/10.1554/06-334.1.
Larget, B. R., Kotha, S. K., Dewey, C. N., & Ané, C. (2010). BUCKy: Gene tree/species tree reconciliation with Bayesian concordance analysis. Bioinformatics, 26, 2910–2911. https://doi.org/10.1093/bioinformatics/btq539.
Lartillot, N., & Philippe, H. (2006). Computing Bayes factors using thermodynamic integration. Systematic Biology, 55, 195–207. https://doi.org/10.1080/10635150500433722.
Leaché, A. D., & Fujita, M. K. (2010). Bayesian species delimitation in West African forest geckos (Hemidactylus fasciatus). Proceedings of the Royal Society B: Biological Sciences, 277(1697), 3071–3077. https://doi.org/10.1098/rspb.2010.0662.
Leaché, A. D., Zhu, T., Rannala, B., & Yang, Z. (2019). The spectre of too many species. Systematic Biology, 68(1), 168–181. https://doi.org/10.1093/sysbio/syy051.
Leier, A., McQuarrie, N., Garzione, C., & Eiler, J. (2013). Stable isotope evidence for multiple pulses of rapid surface uplift in the Central Andes, Bolivia. Earth and Planetary Science Letters, 371, 49–58. https://doi.org/10.1016/j.epsl.2013.04.025.
Levin, D. A. (2003). Ecological speciation: lessons from invasive species. Systematic Botany, 28(4), 643–650. https://doi.org/10.1043/02-70.1.
Liu, C., Berry, P. M., Dawson, T. P., & Pearson, R. G. (2005). Selecting thresholds of occurrence in the prediction of species distributions. Ecography, 28, 385–393. https://doi.org/10.1111/j.0906-7590.2005.03957.x.
Liu, L., Zhao, B., Tan, D., & Wang, J. (2011). Phylogenetic relationships of Brassicaceae in China: insights from a non-coding chloroplast, mitochondrial, and nuclear DNA data set. Biochemical Systematics and Ecology, 39(4–6), 600–608. https://doi.org/10.1016/j.bse.2011.05.003.
Liu, C., White, M., & Newell, G. (2013). Selecting thresholds for the prediction of species occurrence with presence-only data. Journal of Biogeography, 40, 778–789. https://doi.org/10.1111/jbi.12058.
Liu, H., Ye, Q., & Wiens, J. J. (2020). Climatic-niche evolution follows similar rules in plants and animals. Nature Ecology & Evolution, 4(5), 753–763. https://doi.org/10.1038/s41559-020-1158-x.
Losos, J. B., & Glor, R. E. (2003). Phylogenetic comparative methods and the geography of speciation. Trends in Ecology & Evolution, 18(5), 220–227. https://doi.org/10.1016/S0169-5347(03)00037-5.
Luebert, F., & Weigend, M. (2014). Phylogenetic insights into Andean plant diversification. Frontiers in Ecology and Evolution, 2, 27. https://doi.org/10.3389/fevo.2014.00027.
Luo, A., Ling, C., Ho, S. Y., & Zhu, C. D. (2018). Comparison of methods for molecular species delimitation across a range of speciation scenarios. Systematic Biology, 67(5), 830–846. https://doi.org/10.1093/sysbio/syy011.
Lynch, J. D. (1989). The gauge of speciation: on the frequency of modes of speciation. In D. Otte & J. A. Endler (Eds.), Speciation and its consequences (pp. 527–553). Sunderland: Sinauer Associates.
Mallet, J. (2007). Hybrid speciation. Nature, 446(7133), 279–283. https://doi.org/10.1038/nature05706.
Mammola, S. (2019). Assessing similarity of n-dimensional hypervolumes: which metric to use? Journal of Biogeography, 46, 2012–2023. https://doi.org/10.1111/jbi.13618.
Mandakova, T., Li, Z., Barker, M. S., & Lysak, M. A. (2017). Diverse genome organization following 13 independent mesopolyploid events in Brassicaceae contrasts with convergent patterns of gene retention. The Plant Journal, 91(1), 3–21. https://doi.org/10.1111/tpj.13553.
Mandáková, T., Zozomová-Lihová, J., Kudoh, H., Zhao, Y., Lysak, M. A., & Marhold, K. (2019). The story of promiscuous crucifers: origin and genome evolution of an invasive species, Cardamine occulta (Brassicaceae), and its relatives. Annals of Botany, 124(2), 209–220. https://doi.org/10.1093/aob/mcz019.
Mares, M. A., Morello, J., & Goldstein, G. (1985). The Monte Desert and other subtropical semi and arid biomes of Argentina, with comments on their relationships to north American arid areas. In M. Evenari, I. N. Meir, & D. Goodall (Eds.), Hot Deserts and Arid Shrublands (pp. 203–236). Amsterdam: Elsevier.
Marhold, K., & Lihová, J. (2006). Polyploidy, hybridization and reticulate evolution: lessons from the Brassicaceae. Plant Systematics and Evolution, 259(2–4), 143–174. https://doi.org/10.1007/s00606-006-0417-x.
Marhold, K., Šlenker, M., & Zozomová-Lihová, J. (2018). Polyploidy and hybridization in the Mediterranean and neighbouring northern areas: examples from the genus Cardamine (Brassicaceae). Biologia Serbica, 40, 47–59. https://doi.org/10.5281/zenodo.1424339.
Martínez, G. A., Arana, M. D., Oggero, A. J., & Natale, E. S. (2017). Biogeographical relationships and new regionalisation of high-altitude grasslands and woodlands of the central Pampean ranges (Argentina), based on vascular plants and vertebrates. Australian Systematic Botany, 29(6), 473–488. https://doi.org/10.1071/SB16046.
Mayr, E. (1954). Change of genetic environment and evolution. In J. Huxley, A. C. Hardy, & E. B. Ford (Eds.), Evolution as a process (pp. 157–180). New York: Collier Books.
Mayr, E. (1959). Isolation as an evolutionary factor. Proceedings of the American Philosophical Society, 103(2), 221–230.
Mayr, E. (1982). Processes of speciation in animals. In C. Barigozzi (Ed.), Mechanisms of speciation (pp. 1–19). New York: Alan R. Liss.
Miller, M. A., Pfeiffer, W., & Schwartz, T. (2010). Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE) (pp. 1–8). New Orleans: IEEE.
Moritz, C. (1994). Defining ‘evolutionarily significant units’ for conservation. Trends in Ecology & Evolution, 9, 373–375. https://doi.org/10.1016/0169-5347(94)90057-4.
Morrone, J. J. (2015). Biogeographical regionalisation of the Andean región. Zootaxa, 3936, 207–236. https://doi.org/10.11646/zootaxa.3936.2.3.
Morrone, J. J. (2017). Neotropical biogeography: regionalization and evolution, CRC biogeography series. Boca Raton: CRC Press.
Morrone, J. J. (2018). Evolutionary biogeography of the Andean region, CRC biogeography series. Boca Raton: CRC Press.
Naciri, Y., & Linder, H. P. (2015). Species delimitation and relationships: the dance of the seven veils. Taxon, 64(1), 3–16. https://doi.org/10.12705/641.24.
Nieto-Montes de Oca, A., Barley, A. J., Meza-Lázaro, R. N., García-Vázquez, U. O., Zamora-Abrego, J. G., Thomson, R. C., & Leaché, A. D. (2017). Phylogenomics and species delimitation in the knob-scaled lizards of the genus Xenosaurus (Squamata: Xenosauridae) using ddRADseq data reveal a substantial underestimation of diversity. Molecular Phylogenetics and Evolution, 106, 241–253. https://doi.org/10.1016/j.ympev.2016.09.001.
Nosil, P. (2012). Ecological speciation. New York: Oxford University Press.
Nosil, P., & Harmon, L. (2009). Niche dimensionality and ecological speciation. In R. Butlin, J. Bridle, & D. Schluter (Eds.), Speciation and patterns of diversity (pp. 127–154). New York: Cambridge University Press.
Nürk, N. M., Uribe-Convers, S., Gehrke, B., Tank, D. C., & Blattner, F. R. (2015). Oligocene niche shift, Miocene diversification–cold tolerance and accelerated speciation rates in the St. John’s Worts (Hypericum, Hypericaceae). BMC Evolutionary Biology, 15(1), 80. https://doi.org/10.1186/s12862-015-0359-4.
Nürk, N. M., Michling, F., & Linder, H. P. (2018). Are the radiations of temperate lineages in tropical alpine ecosystems pre-adapted? Global Ecology and Biogeography, 27(3), 334–345. https://doi.org/10.1111/geb.12699.
O’Kane Jr., S. L. (2012). Evolution and classification of Physaria (Brassicaceae). In Principles of Crucifer Evolution, Botany 2012 Symposium. Columbus: Botanical Society of America.
O’Kane Jr., S. L., & Al-Shehbaz, I. A. (2004). The genus Physaria (Brassicaceae) in South America. Novon, 196–205.
Oksanen J., Blanchet F. G., Friendly M., Kindt R., Legendre P., McGlinn D., Minchin P. R., O’Hara R. B., Simpson G. L., Solymos P., Stevens M. H., Szoecs E., Wagner H. (2019). vegan: community ecology package. R package version 2.5-6. https://CRAN.R-project.org/package=vega. Accessed December 2019
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C., D’amico, J. A., Itoua, I., Strand, H. E., Morrison, J. C., Loucks, C. J., Allnutt, T. F., Ricketts, T. H., Kura, Y., Lamoreux, J. F., Wettengel, W. W., Hedao, P., & Kassem, K. R. (2001). Terrestrial ecoregions of the world: a new map of life on Earth. BioScience, 51, 933–938. https://doi.org/10.1641/0006-3568(2001)051[0933:TEOTWA]2.0.CO;2.
Ortiz-Jaureguizar, E., & Cladera, G. A. (2006). Paleoenvironmental evolution of southern South America during the Cenozoic. Journal of Arid Environments, 66(3), 498–532. https://doi.org/10.1016/j.jaridenv.2006.01.007.
Paradis, E. (2010). Pegas: an R package for population genetics with an integrated–modular approach. Bioinformatics, 26, 419–420. https://doi.org/10.1093/bioinformatics/btp696.
Paradis, E., & Schliep, K. (2018). ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics, 35, 526–528. https://doi.org/10.1093/bioinformatics/bty633.
Pennington, R. T., Lavin, M., Prado, D. E., Pendry, C. A., Pell, S. K., & Butterworth, C. A. (2004). Historical climate change and speciation: Neotropical seasonally dry forest plants show patterns of both Tertiary and Quaternary diversification. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 359(1443), 515–538. https://doi.org/10.1098/rstb.2003.1435.
Peterson, A. T. (2011). Ecological niche conservatism: a time-structured review of evidence. Journal of Biogeography, 38(5), 817–827. https://doi.org/10.1111/j.1365-2699.2010.02456.x.
Peterson, A. T., & Lieberman, B. S. (2012). Species’ geographic distributions through time: playing catch-up with changing climates. Evolution: Education and Outreach, 5(4), 569. https://doi.org/10.1007/s12052-012-0385-2.
Peterson, A. T., Soberón, J., & Sánchez-Cordero, V. (1999). Conservatism of ecological niches in evolutionary time. Science, 285, 1265–1267. https://doi.org/10.1126/science.285.5431.1265.
Phillimore, A. (2014). Geography, range evolution, and speciation. In J. Losos, D. Baum, D. Futuyma, H. Hoekstra, R. Lenski, A. Moore, C. L. Peichel, D. Schluter, & M. Whitlock (Eds.), The Princeton guide to evolution (pp. 50–511). New Jersey: Princeton University Press.
Phillips, S. J., Anderson, R. P., Dudík, M., Schapire, R. E., & Blair, M. E. (2017). Opening the black box: an open-source release of Maxent. Ecography, 40, 887–893. https://doi.org/10.1111/ecog.03049.
Ponce, J. F., Rabassa, J., Coronato, A., & Borromei, A. M. (2011). Palaeogeographical evolution of the Atlantic coast of Pampa and Patagonia from the last glacial maximum to the Middle Holocene. Biological Journal of the Linnean Society, 103(2), 363–379. https://doi.org/10.1111/j.1095-8312.2011.01653.x.
Pons, J., Barraclough, T. G., Gomez-Zurita, J., Cardoso, A., Duran, D. P., Hazell, S., Kamoun, S., Sumlin, W. D., & Vogler, A. P. (2006). Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Systematic Biology, 55(4), 595–609. https://doi.org/10.1080/10635150600852011.
Pyron, R. A., Costa, G. C., Patten, M. A., & Burbrink, F. T. (2015). Phylogenetic niche conservatism and the evolutionary basis of ecological speciation. Biological Reviews, 90(4), 1248–1262. https://doi.org/10.1111/brv.12154.
Quantum GIS Development Team. (2016). QGIS Geographic Information System. Open Source Geospatial Foundation. http://qgis.org. Accessed December 2017
R Core Team. (2018). R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing https://www.R-project.org. Accessed Jan 2019
Rabassa, J. (2008). Late Cenozoic glaciations in Patagonia and Tierra del Fuego. Developments in Quaternary Sciences, 11, 151–204. https://doi.org/10.1016/S1571-0866(07)10008-7.
Rabassa, J., & Ponce, J. F. (2013). The Heinrich and Dansgaard-Oeschger climatic events during Marine Isotopic Stage 3: searching for appropriate times for human colonization of the Americas. Quaternary International, 299, 94–105.
Rabassa, J., Coronato, A., & Martinez, O. (2011). Late Cenozoic glaciations in Patagonia and Tierra del Fuego: an updated review. Biological Journal of the Linnean Society, 103(2), 316–335. https://doi.org/10.1111/j.1095-8312.2011.01681.x.
Rambaut, A., Drummond, A. J., Xie, D., Baele, G., & Suchard, M. A. (2018). Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Systematic Biology, 67, 901–904. https://doi.org/10.1093/sysbio/syy032.
Ramsey, J. (2011). Polyploidy and ecological adaptation in wild yarrow. Proceedings of the National Academy of Sciences of United States of America, 108(17), 7096–7101. https://doi.org/10.1073/pnas.1016631108.
Rannala, B., & Yang, Z. (2017). Efficient Bayesian species tree inference under the multispecies coalescent. Systematic Biology, 66(5), 823–842. https://doi.org/10.1093/sysbio/syw119.
Renner, M. A., Heslewood, M. M., Patzak, S. D., Schäfer-Verwimp, A., & Heinrichs, J. (2017). By how much do we underestimate species diversity of liverworts using morphological evidence? An example from Australasian Plagiochila (Plagiochilaceae: Jungermanniopsida). Molecular Phylogenetics and Evolution, 107, 576–593. https://doi.org/10.1016/j.ympev.2016.12.018.
Revell, L. J. (2012). phytools: an R package for phylogenetic comparative biology (and other things). Methods in Ecology and Evolution, 3, 217–223. https://doi.org/10.1111/j.2041-210X.2011.00169.x.
Ricklefs, R. E. (1987). Community diversity: relative roles of local and regional processes. Science, 235(4785), 167–171. https://doi.org/10.1126/science.235.4785.167.
Riddle, B. R., & Hafner, D. J. (1999). Species as units of analysis in ecology and biogeography: time to take the blinders off. Global Ecology and Biogeography, 8(6), 433–441. https://doi.org/10.1046/j.1365-2699.1999.00170.x.
Rieseberg, L. H., & Willis, J. H. (2007). Plant speciation. Science, 317(5840), 910–914. https://doi.org/10.1126/science.1137729.
Roig, F. A., Roig-Juñent, S., & Corbalán, V. (2009). Biogeography of the Monte desert. Journal of Arid Environments, 73(2), 164–172. https://doi.org/10.1016/j.jaridenv.2008.07.016.
Rollins, R. C. (1939). The cruciferous genus Physaria. Contributions from the Gray Herbarium of Harvard University, 126, 392–415.
Romano, G. M. (2017). A high resolution shapefile of the Andean biogeographical region. Data in Brief, 13, 230–232. https://doi.org/10.1016/j.dib.2017.05.039.
Ronquist, F., Teslenko, M., van der Mark, P., Ayres, D. L., Darling, A., Höhna, S., Larget, B., Liu, L., Suchard, M. A., & Huelsenbeck, J. P. (2012). MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a Large Model Space. Systematic Biology, 61, 539–542. https://doi.org/10.1093/sysbio/sys029.
Rull, V. (2011). Neotropical biodiversity: timing and potential drivers. Trends in Ecology & Evolution, 26(10), 508–513. https://doi.org/10.1016/j.tree.2011.05.011.
Rundle, H. D., & Nosil, P. (2005). Ecological speciation. Ecology Letters, 8(3), 336–352. https://doi.org/10.1111/j.1461-0248.2004.00715.x.
Rutter, N., Coronato, A., Helmens, K., Rabassa, J., & Zárate, M. (2012). Glaciations in North and South America from the Miocene to the Last Glacial Maximum. Comparisons, linkages and uncertainties. New York: Springer. https://doi.org/10.1007/978-94-007-4399-1.
Ryder, O. A. (1986). Species conservation and systematics: the dilemma of subspecies. Trends in Ecology & Evolution, 1, 9–10.
Salariato, D. L., & Zuloaga, F. O. (2017). Climatic niche evolution in the Andean genus Menonvillea (Cremolobeae: Brassicaceae). Organisms Diversity & Evolution, 17, 11–28. https://doi.org/10.1007/s13127-016-0291-5.
Salariato, D. L., Zuloaga, F. O., & Al-Shehbaz, I. A. (2013). Revision and tribal placement of the Argentinean genus Parodiodoxa (Brassicaceae). Plant Systematics and Evolution, 299, 305–316. https://doi.org/10.1007/s00606012-0722-5.
Salariato, D. L., Zuloaga, F. O., Cano, A., & Al-Shehbaz, I. A. (2015). Molecular phylogenetics of tribe Eudemeae (Brassicaceae) and implications for its morphology and distribution. Molecular Phylogenetics and Evolution, 82, 43–59. https://doi.org/10.1016/j.ympev.2014.09.030.
Salariato, D. L., Zuloaga, F. O., Franzke, A., Mummenhoff, K., & Al-Shehbaz, I. A. (2016). Diversification patterns in the CES clade (Brassicaceae tribes Cremolobeae, Eudemeae, Schizopetaleae) in Andean South America. Botanical Journal of the Linnean Society, 181, 543–566. https://doi.org/10.1111/boj.12430.
Salariato, D. L., Al-Shehbaz, I. A., & Zuloaga, F. O. (2018). Reinstatement of the southern Andean genus Stenodraba (Brassicaceae) based on molecular data and insights from its environmental and geographic distribution. Systematic Botany, 43, 35–52. https://doi.org/10.1600/036364418X697085.
Salariato, D. L., Cano, A., Zuloaga, F. O., & Al-Shehbaz, I. A. (2020). Molecular phylogeny of Cremolobus (Brassicaceae) supports the recognition of the new genus Yunkia and demonstrates the high habitat diversity of tribe Cremolobeae. Systematics and Biodiversity, 18, 295–314. https://doi.org/10.1080/14772000.2020.1739777.
Schemske, D. W. (2010). Adaptation and the origin of species. The American Naturalist, 176(S1), S4–S25. https://doi.org/10.1086/657060.
Schluter, D. (2000). The Ecology of Adaptive Radiation. Oxford: Oxford University Press.
Schluter, D. (2001). Ecology and the origin of species. Trends in Ecology & Evolution, 16(7), 372–380. https://doi.org/10.1016/S0169-5347(01)02198-X.
Schluter, D. (2009). Evidence for ecological speciation and its alternative. Science, 323(5915), 737–741. https://doi.org/10.1126/science.1160006.
Simpson, G. G. (1951). The species concept. Evolution, 5, 285–298. https://doi.org/10.1111/j.1558-5646.1951.tb02788.x.
Simpson, G. G. (1953). The major features of evolution. New York: Columbia University Press.
Soberón, J. (2007). Grinnellian and Eltonian niches and geographic distributions of species. Ecology Letters, 10(12), 1115–1123. https://doi.org/10.1111/j.1461-0248.2007.01107.x.
Solbrig, O. T., Blair, W. F., Enders, F. A., Hulse, A. C., Hunt, J. H., Mares, M. A., Neff, J., Otte, D., Simpson, B. B., & Tomoff, C. S. (1977). The biota, the dependent variable. In G. H. Orians & O. T. Solbrig (Eds.), Convergent Evolution in Warm Deserts, an Examination of Strategies and Patterns in Deserts of Argentina and the United States (pp. 50–66). Pennsylvannia: Dowden, Hutchinson & Ross Inc..
Spalink, D., Drew, B. T., Pace, M. C., Zaborsky, J. G., Li, P., Cameron, K. M., Givnish, T. J., & Sytsma, K. J. (2016). Evolution of geographical place and niche space: Patterns of diversification in the North American sedge (Cyperaceae) flora. Molecular Phylogenetics and Evolution, 95, 183–195. https://doi.org/10.1016/j.ympev.2015.09.028.
Stamatakis, A. (2014). RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics, 30, 1312–1313. https://doi.org/10.1093/bioinformatics/btu033.
Stamatakis, A., Hoover, P., & Rougemont, J. (2008). A rapid bootstrap algorithm for the RAxML web-servers. Systematic Biology, 57, 758–771. https://doi.org/10.1080/10635150802429642.
Sudhaus, W. (2004). Radiation within the framework of evolutionary ecology. Organisms Diversity & Evolution, 4(3), 127–134. https://doi.org/10.1016/j.ode.2004.04.001.
Sukumaran, J., & Knowles, L. L. (2017). Multispecies coalescent delimits structure, not species. Proceedings of the National Academy of Sciences, 114(7), 1607–1612. https://doi.org/10.1073/pnas.1607921114.
Taberlet, P., Gielly, L., Pautou, G., & Bouvet, J. (1991). Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Molecular Biology, 17, 1105–1109. https://doi.org/10.1007/BF00037152.
Tewes, L. J., Michling, F., Koch, M. A., & Müller, C. (2018). Intracontinental plant invader shows matching genetic and chemical profiles and might benefit from high defence variation within populations. Journal of Ecology, 106(2), 714–726. https://doi.org/10.1111/1365-2745.12869.
Thiers B. (2020). [continuously updated]. Index Herbariorum: a global directory of public herbaria and associated staff. New York Botanical Garden’s Virtual Herbarium. http://sweetgum.nybg.org/science/ih/. Accessed March 2020.
Trabucco, A., & Zomer, R. (2019). Global Aridity Index and Potential Evapotranspiration (ET0) Climate Database v2. figshare. Dataset. https://doi.org/10.6084/m9.figshare.7504448.v3. Accessed July 2019
Uba, C. E., Hasler, C. A., Buatois, L. A., Schmitt, A. K., & Plessen, B. (2009). Isotopic, paleontologic, and ichnologic evidence for late Miocene pulses of marine incursions in the central Andes. Geology, 37, 827–830. https://doi.org/10.1130/G30014A.1.
Warren, D. L., Glor, R. E., & Turelli, M. (2008). Environmental niche equivalency versus conservatism: quantitative approaches to niche evolution. Evolution, 62, 2868–2883. https://doi.org/10.1111/j.1558-5646.2008.00482.x.
Warwick, S. I., & Al-Shehbaz, I. A. (2006). Brassicaceae: chromosome number index and database on CD-Rom. Plant Systematics and Evolution, 259, 237–248. https://doi.org/10.1007/s00606-006-0421-17.
Wiens, J. J. (2004a). Speciation and ecology revisited: phylogenetic niche conservatism and the origin of species. Evolution, 58(1), 193–197. https://doi.org/10.1554/03-447.
Wiens, J. J. (2004b). What is speciation and how should we study it? The American Naturalist, 163(6), 914–923. https://doi.org/10.1086/386552.
Wiens, J. J. (2012). Perspective: why biogeography matters: historical biogeography vs. phylogeography and community phylogenetics for inferring ecological and evolutionary processes. Frontiers of Biogeography, 4(3), 128–135.
Wiens, J. J., & Graham, C. H. (2005). Niche conservatism: integrating evolution, ecology, and conservation biology. Annual Review of Ecology, Evolution, and Systematics, 36, 519–539. https://doi.org/10.1146/annurev.ecolsys.36.102803.095431.
Wiens, J. J., Ackerly, D. D., Allen, A. P., Anacker, B. L., Buckley, L. B., Cornell, H. V., Ellen, I., Damschen, E. I., Davies, T. J., Grytnes, J. A., Harrison, S. P., Hawkins, B. A., Holt, R. D., McCain, C. M., & Stephens, P. R. (2010). Niche conservatism as an emerging principle in ecology and conservation biology. Ecology Letters, 13(10), 1310–1324. https://doi.org/10.1111/j.1461-0248.2010.01515.x.
Wiley, E. O. (1978). The evolutionary species concept reconsidered. Systematic Zoology, 27, 17–26.
Xie, W., Lewis, P. O., Fan, Y., Kuo, L., & Chen, M. H. (2011). Improving marginal likelihood estimation for Bayesian phylogenetic model selection. Systematic Biology, 60, 150–160. https://doi.org/10.1093/sysbio/syq085.
Yang, Z. (2015). The BPP program for species tree estimation and species delimitation. Current Zoology, 61(5), 854–865. https://doi.org/10.1093/czoolo/61.5.854.
Yang, Z., & Rannala, B. (2014). Unguided species delimitation using DNA sequence data from multiple loci. Molecular Biology and Evolution, 31(12), 3125–3135. https://doi.org/10.1093/molbev/msu279.
Young, K. R., Ulloa Ulloa, C., Luteyn, J. L., & Knapp, S. (2002). Plant evolution and endemism in Andean South America: an introduction. The Botanical Review, 68, 4–21. https://doi.org/10.1663/0006-8101(2002)068[0004:PEAEIA]2.0.CO;2.
Zachos, J., Pagani, M., Sloan, L., Thomas, E., & Billups, K. (2001). Trends, rhythms, and aberrations in global climate 65 Ma to present. Science, 292, 686–693. https://doi.org/10.1126/science.1059412.
Acknowledgements
Our deep gratitude goes to Dr. Ihsan A. Al-Shehbaz for the critical review of this work and his valuable support, guidance, and suggestions in the study of South American Brassicaceae over the years. We thank the directors, curators, and collection managers of the herbaria listed. Special thanks are to Lone Aagesen, Juan M. Acosta, and Ariel Lliully Aguilar for their help in the fieldtrips. We especially appreciate the help of editors and two anonymous reviewers who provided useful suggestions to improve the early version of this paper. Botanical collections associated to this project were conducted under the permits: APN No. 1103 and 1546 (Argentina) and VMABCC No. 026/09 (Bolivia).
Funding
This work was funded by ANPCyT (Agencia Nacional de Promoción Científica y Tecnológica) grant PICT-2016-0096, CONICET (Consejo Nacional de Investigaciones Científicas y Técnicas) grant PIP-112-201301-00124CO, and the National Geographic Society grant #9841-16, for which we are profoundly grateful.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical approval
This article does not use animals and does not contain any studies with human participants performed by any of the authors.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Fig. S1
Maximum likelihood tree generated with nrITS sequences and RAxML 8.2.10 showing the monophyly of South American Physaria. Values on branches correspond to Bootstrap support ≥ 50%. Units of branch length are proportional to nucleotide substitutions per site. (PDF 256 kb)
Fig. S2
Distribution map of South American Physaria and delimited area corresponding to the 50 km1423 buffered minimum convex polygon (study area) used in the niche analyses and geographic range comparisons. (PDF 1266 kb)
Fig. S3
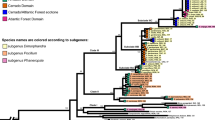
Maximum likelihood trees generated with RAxML 8.2.10 showing the phylogenetic placement of sampled specimens of South American Physaria. a, nrITS dataset. b, cpDNA dataset (trnL-F/trnH-psbA/trnG intron/trnS-trnG). c, concatenated nrITS+cpDNA dataset. Values on branches correspond to Bootstrap support ≥ 50%. Units of branch length are proportional to nucleotide substitutions per site. Boxes to the rigth show delimited lineages. For tip labels: CRA, P. crassistigma, LAT, P. lateralis; MEN, P. mendocina; OKA, P. okanensis; PYG, P. pygmaea; URB: P. urbaniana. Image: Flower and fruit of P. mendocina (PDF 793 kb)
Fig. S4
Maximum clade credibility trees obtained from the analyses with MrBayes 3.2.6 showing the phylogenetic placement of sampled specimens of South American Physaria. a, nrITS dataset. b, cpDNA dataset (trnL-F/trnH-psbA/trnG intron/trnS-trnG). c, concatenated n rITS+cpDNA dataset. Circles on branches correspond to posterior probabilities (PP) ≥ 50%. Units of branch length are proportional to nucleotide substitutions per site. Boxes to the rigth show delimited lineages. For tip labels: CRA, P. crassistigma, LAT, P. lateralis; MEN, P. mendocina; OKA, P. okanensis; PYG, P. pygmaea; URB: P. urbaniana. Image: flower and fruit of P. crassistigma. (PDF 937 kb)
Fig. S5
Phylogenetic placement of sampled specimens of South American Physaria. a, Primary concordance tree from the BUCKy analyses using the 6002 trees from the nrITS and the cpDNA datasets obtained in the MrBayes analyses and under α = 1. Concordance factor values (CF) ≥ 0.5 are shown on branches. Units of branch length are proportional to the concordance factor. Results from concordance analyses varying the discordance prior (α) had no effect on topology or concordance. Stars on nodes indicate delimited lineages of South American Physaria. b, filtered supernetwork generated in SplitsTree 4.14.8 using 1000 Bayesian posterior trees per each nrITS and cpDNA dataset with the Z-closure algorithm, and filtering the splits to show only those present in a minimum of 30% input trees. Shades groups represent delimited lineages. (PDF 537 kb)
Fig. S6
Results from the climatic niche analyses in the environmental space obtained using the PCA env method. a–c, Correlation circles showing the contribution of environmental variables on the first three axes of the PCA-env and the percentage of inertia explained. a, PC1 vs PC2. b, PC1 vs PC3. c, PC2 vs PC3. d, Bar plot from the broken-stick analysis. PCA-env axes with larger percentages of accumulated inertia than the broken-stick variances are significant (three in this case). (PDF 31 kb)
Fig. S7
Univariate climatic niche comparisons in the environmental space among delimited lineages of South American Physaria. a, density plots computed for each independently evolving lineage using the PCenv1, PCenv2, PCenv3, elevation, annual mean temperature (BIO1), and annual mean precipitation (BIO12). b, p-values from Kruskal-Wallis test together with the Bonferroni correction for multiple comparisons to assess for differences between Physaria lineages for the three first PCs, elevation, BIO1, and BIO12. Values marked with an asterisk (red cells) are rejected at α=0.05. Labels: CRA, crassistigma lineage; NLA, northern lateralis lineage; SLA, southern lateralis lineage; MEN, mendocina lineage; URB, urbaniana lineage; YUN, yungas lineage. (PDF 645 kb)
Fig. S8
Geographic range overlap of delimited lineages of South American Physaria for the present time. Binary (presence/absence) distributions maps for delimited lineages derived from the species distribution modelling (SDM) in MaxEnt using the maximum training sensitivity plus specificity as threshold. Blue and green cells indicate presence of only one lineage, while red cells indicate presence of both species (sympatric cells). (PDF 1635 kb)
Fig. S9
Geographic range overlap of delimited lineages of South American Physaria for the Last Glacial Maximum (LGM) (~ 21 kya). Binary (presence/absence) distributions maps for delimited lineages derived from the species distribution modelling (SDM) in MaxEnt using the maximum training sensitivity plus specificity as threshold. Blue and green cells indicate presence of only one lineage, while red cells indicate presence of both species (sympatric cells). (PDF 2255 kb)
Table S1
(DOCX 13 kb)
Table S2
(DOCX 5 kb)
Table S3
(DOCX 10 kb)
Table S4
(DOCX 11 kb)
ESM 14
ITS alignment for the South American Physaria dataset used for phylogenetic analyses in this study. (NEX 32 kb)
ESM 15
trnL-F alignment for the South American Physaria dataset used for phylogenetic analyses in this study. (NEX 39 kb)
ESM 16
trnH-psbA alignment for the South American Physaria dataset used for phylogenetic analyses in this study. (NEX 15 kb)
ESM 17
trnG intron alignment for the South American Physaria dataset used for phylogenetic analyses in this study. (NEX 31 kb)
ESM 18
trnS-trnG alignment for the South American Physaria dataset used for phylogenetic analyses in this study. (NEX 26 kb)
Appendix 1
Appendix 1
Taxa and GenBank accession numbers for the ITS, trnH-psbA , trnL-F, trnG intron, and trnS-trnG sequences used in the molecular analyses.
Physaria fendleri (A. Gray) O'Kane & Al-Shehbaz (outgroup) (AF055198, -, AF055266, -). Physaria crassistigma O'Kane & Al-Shehbaz. ARGENTINA. Mendoza. Las Heras: Above Villavicencio on road to Paramillo de Uspallata, roadside, open areas among shrubs, ca. 44 km before Uspallata, Al-Shehbaz et al. 808 (SI) (CRA6) (MW366047, MW366135, MW366090, MW366180, MW366223). Luján de Cuyo: Centro de Ski Vallecitos, ladera derecha de cerros subiendo por la pista de ski, Salariato et al. 252 (SI) (CRA3) (MW366023, MW366110, MW366068, MW366157, MW366201). Tunuyán: RP94, Camino al Portillo Argentino, 0.5 km de las Llaretas desde el puesto de gendarmeria hacia el portillo, Salariato et al. 207 (SI) (CRA1) (MW366021, MW366108, MW366066, MW366155, MW366199); camino del Portillo Argentino, desde Las Llaretas hacia el hotel abandonado, ca. 3 km del desvío a Valle de Manantiales, Salariato et al. 233 (SI) (CRA2) (MW366022, MW366109, MW366067, MW366156, MW366200); Ruta Provincial 94 de Manzano Histórico a Portillo Argentino, Zuloaga et al. 5292 (SI) (CRA5) (MW366040, MW366128, MW366083, MW366173, MW366216). Tupungato: Ruta provincial n° 94: entre Los Manantiales y Real de Piedras Coloradas, Cocucci & Sérsic 2253 (CORD, SI) (CRA4) (MW366037, MW366125, MW366080, MW366170, MW366214). Physaria lateralis O'Kane & Al-Shehbaz. ARGENTINA. Mendoza. Las Heras: Ruta Provincial 13, alrededor de Monolito, Giussani et al. 602 (SI) (LAT8) (MW366048, MW366137, MW366092, MW366182, MW366225). Malargüe: Base del Cerro Nevado por Ruta Prov. 180, Prina et al. 2630 (SI) (LAT6) (MW366038, MW366126, MW366081, MW366171, -). San Carlos: Refugio militar Gral. Alvarado, entrada a la Reserva Laguna del Diamante, Al-Shehbaz et al. 813 (SI) (LAT10) (MW366050, MW366139, -, MW366184, MW366226). San Rafael: Cerro Nevado, Prina, et al. 2917 (SRFA, SI) (LAT5) (MW366036, MW366124, MW366079, MW366169, -); de Termas del Sosneado a Laguna El Sosneado, Zuloaga et al. 12408 (SI) (LAT9) (MW366049, MW366138, MW366093, MW366183, -); Ruta Nacional 144, cuesta de los Terneros hacia 25 de Mayo, Zuloaga et al. 15277 (SI) (LAT11) (MW366042, MW366130, MW366085, MW366175, MW366218). San Luis. Coronel Pringles: Ruta Provincial 9, entre Trapiche y La Carolina, Aagensen 26 (SI) (LAT1) (MW366015, MW366102, MW366060, MW366149, MW366193); Ruta provincial 9, Monumento y Museo Lafinur, pasando La Carolina, Biganzoli 2034 (SI) (LAT7) (MW366039, MW366127, MW366082, MW366172, MW366215); Alrededores de la Gruta de Intihuasi, Cerana 1266 (CORD) (LAT4) (MW366033, MW366121, MW366076, MW366166, MW366211); Camino a San Francisco 10-15 km de La Carolina, Cerana 1330 (CORD) (LAT3) (MW366028, MW366115, -, -, -); Cañada Honda, Cerana 1360 (CORD) (LAT2) (MW366027, MW366114, -, MW366161, MW366205). Physaria mendocina (Phil.) O'Kane & Al-Shehbaz. ARGENTINA. Córdoba. Calamuchita: San Agustin (Cerro La Cruz), Cantero 6107 (CORD) (MEN4) (MW366020, MW366107, MW366065, MW366154, MW366198). Colón: Candonga, Cantero 5897 (CORD) (MEN8) (MW366029, MW366116, MW366072, MW366162, MW366206). Punilla: Quilpo (Canteras), Cantero 5928 (CORD) (MEN7) (MW366026, MW366113, MW366071, MW366160, MW366204); Valle Hermoso, Cantero 6039 (CORD) (MEN6) (MW366025, MW366112, MW366070, MW366159, MW366203). Rio Cuarto: Alta Gracia, Cantero 5834 (CORD) (MEN9) (MW366030, MW366117, MW366073, MW366163, MW366207); Achiras (Monte Guazú), Cantero 6291 (CORD) (MEN3) (MW366019, MW366106, MW366064, MW366153, MW366197); Cuchi Yaco (Cerca Salsacate), Cantero 5656 (CORD) (MEN10) (MW366031, MW366118, MW366074, MW366164, MW366208); El Saucesito, Cantero 5565 (CORD) (MEN11) (MW366032, MW366119, MW366075, MW366165, MW366209); Iguazu, Cantero 5550 (CORD) (MEN2) (MW366018, MW366105, MW366063, MW366152, MW366196); Mesa de la Argentina, Cantero 5636 (CORD) (MEN5) (MW366024, MW366111, MW366069, MW366158, MW366202). La Pampa. Lihué Calel: Parque Nacional Lihue Calel, camino interior hacia cerros más bajos y pinturas rupestres, Zuloaga & Salariato 16073 (SI) (MEN1) (MW366016, MW366103, MW366061, MW366150, MW366194). Mendoza. Las Heras: de la RN 14 hacia los Berros, RN 153, Zuloaga et al. 15741 (SI) (MEN14) (-, MW366136, MW366091, MW366181, MW366224). Tunuyán: Manzano Histórico, Deginani et al. 2173 (SI) (MEN16) (MW366043, MW366131, MW366086, MW366176, MW366219). Tupungato: Bajando del Cerro Cristo Rey, Deginani et al. 2295 (SI) (MEN15) (MW366052, MW366141, MW366095, MW366186, MW366228). Río Negro. Valcheta: En km 50, cerros basálticos, Kiesling 9923 (SI) (MEN13) (MW366035, MW366123, MW366078, MW366168, MW366213). San Juan. Angaco: Pie de Palo, Kiesling et al. 9384 (SI) (MEN12) (MW366034, MW366122, MW366077, MW366167, MW366212). Physaria okanensis Al-Shehbaz & Prina. ARGENTINA. Catamarca. Ambato: al norte de cumbres de cerro el manchao, Halloy 809 (LIL) (OKA2) (-, MW366120, -, -, MW366210); subiendo hacia el manchao desde el rodeo, en el sitio Falda del Momo, Cantero et al. 7254 (CORD) (OKA1) (MW366017, MW366104, MW366062, MW366151, MW366195). Physaria pygmaea O'Kane & Al-Shehbaz. ARGENTINA. Catamarca. Andalgalá: Capillitas, alrededores del globo meteorológico, Acosta & von Mering 624 (SI) (PYG1) (MW366055, MW366144, MW366098, MW366189, MW366231); cerca de Mina Capillitas. subida al globo meteorológico, Salomón et al. 234 (SI) (PYG3) (MW366044, MW366132, MW366087, MW366177, MW366220). Jujuy. Humahuaca: Mina Aguilar, Espinazo del Diablo, Zuloaga et al. 13572 (SI) (PYG4) (MW366046, MW366134, MW366089, MW366179, MW366222). La Rioja. Famatina: Pampa de Tambería, Barboza et al. 3359 (CORD) (PYG2) (MW366058, MW366147, -, MW366191, MW366234). Physaria urbaniana (Muschl.) O'Kane & Al-Shehbaz. ARGENTINA. La Rioja. Famatina: Entre la bifurcación hacia Tres Piedras y Los Tambitos, Barboza et al. 2875 (CORD) (URB4) (MW366057, MW366146, MW366100, MW366190, MW366233); Pampa de Achavil, pasando el Río Achavil rumbo a Tres Piedras, en la pampa del "Ajenjo", Barboza et al. 2854 (CORD) (URB3) (MW366056, MW366145, MW366099, -, MW366232). Salta. Cachi: Cuesta de obispo, 200 mts pasando piedra del Molino, Zuloaga et al. 12956 (SI) (URB6) (MW366041, MW366129, MW366084, MW366174, MW366217). Cachi: Parque Nacional Los Cardones, pasando el Cerro Negro o Malacanta. en curso de agua, Zanotti et al. 787 (SI) (URB2) (MW366053, MW366142, MW366096, MW366187, MW366229); RP 33, Cuesta del Obispo, Zanotti et al. 800 (SI) (URB1) (MW366051, MW366140, MW366094, MW366185, MW366227); RP 33, Cuesta del Obispo, Piedra del Molino, Acosta & von Mering 607 (SI) (URB7) (MW366054, MW366143, MW366097, MW366188, MW366230). Tucumán. Tafí: RP 307, Abra el Infiernillo, Zanotti et al. 860 (SI) (URB8) (MW366045, MW366133, MW366088, MW366178, MW366221). BOLIVIA. Tarija. Yunchara: al S. de Copacabana, Beck et al. 23756 (LPB) (URB5) (MW366059, MW366148, MW366101, MW366192, MW366235).
Rights and permissions
About this article

Cite this article
Salariato, D.L., Zuloaga, F.O. Ecological and spatial patterns associated with diversification of South American Physaria (Brassicaceae) through the general concept of species. Org Divers Evol 21, 161–188 (2021). https://doi.org/10.1007/s13127-021-00486-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13127-021-00486-z