
Abstract
Purpose
Hyporheic biofilms are the central site for biogeochemical cycling in streams and rivers. In view of global warming and increasing human pressures, this study aimed to compare the response of hyporheic biofilm biomass and activities from an unpolluted reference stream reach surrounded by forest with those from a stream reach exposed to agricultural and urban land use using a mesocosm experiment in which the water temperature and dissolved organic carbon (DOC) contents were manipulated.
Methods
Hyporheic sediments collected in the field from the two study reaches (i.e. reference and impacted) were incubated in the laboratory at two different temperatures (10 °C, 14 °C) and wetted with three types of synthetic water (control [C] – 0 mg L−1; low DOC – 5 mg L−1; high DOC – 30 mg L−1) for four weeks. The responses of the hyporheic biofilms were measured weekly using structural (total protein content [TPC] as a proxy for biofilm biomass) and functional measures (electron transport system activity [ETSA] and community-level physiological profiling [CLPP]).
Results
The response of hyporheic biofilms to temperature changes and DOC enrichment was site-specific for all studied measures (TPC, ETSA and CLPP, including measured average well colour development [AWCD]). The addition of DOC to biofilms from the pristine stream reach significantly heightened the responses at 10 °C, a temperature within the normal environmental temperature ranges of the reference location, but not at 14 °C, which was here, a temperature outside normal environmental range. On the other hand, biofilms from the impacted stream reach exhibited increased responses following DOC enrichment under both temperature regimes, with a particularly pronounced response at 14 ºC, in this case, both experimental temperatures were within the normal environmental temperature ranges of the study locations.
Conclusion
Hyporheic biofilms were shown to be, like benthic biofilms, sensitive to temperature changes and organic enrichment, but their response to temperature changes and enrichment caused by climate change and/or other anthropogenic pressures (i.e. point and non-point pollution, removal of the riparian zone, hydromorphological modifications, etc.) was not simply linear but site-specific. The intensity of the response, characterized by increased activity and biomass production, appears to be constrained within the temperature ranges prevalent in the environment from which the biofilms originate. These findings emphasize the importance of site-specific considerations in predicting the impacts of climate change and anthropogenic pressures on these critical components of river and stream ecosystems.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Freshwater ecosystems, including rivers and streams, are exposed to a range of stressors resulting from intensive water and land use, affecting both ecosystem structure and function (Wagenhoff et al. 2011; Mori et al. 2019). In addition, freshwaters are highly vulnerable to climate change, which affects water temperatures and induces shifts in hydrological patterns (Woodward et al. 2010). Current climate models predict that mean air temperature will increase by up to 4.4 °C by the year 2100 (IPCC 2021), and a recent study from Switzerland (Michel et al. 2022) suggests that river water will warm significantly in the twenty-first century. Predictions from this study are that by the end of the century (2080–2090), the mean annual water temperature increase will range from + 0.9 °C for low-emission scenarios to + 3.5 °C for high-emission scenarios for lowland and alpine catchments. As temperature is an important environmental factor affecting every aspect of living organisms—from molecules to cells to ecosystem processes—these changes will inevitably affect river ecosystem structure and function, including microbial community composition and biogeochemical processes.
Aquatic biofilms are heterogeneous consortia of microorganisms (i.e. archaea, bacteria, fungi, algae and protozoans) embedded in a matrix of extracellular polymeric substances (Battin et al. 2016). Species-rich, surface-attached biofilms dominate microbial life in streams and rivers, where they contribute substantially to biogeochemical transformations and thus play an essential role in stream ecosystem functions (Besemer 2015; Battin et al. 2016). Most ecosystem processes in streams driven by microorganisms, including mineralisation of organic matter, degradation of pollutants, nutrient transformations and other redox-related processes, occur in the hyporheic zone (i.e. at the dynamic interface between the river channel and underlying sediments along the stream network sensu Orghidan (1959, 2010). However, most studies addressing the microbial communities in streams have investigated benthic or epilithic biofilms that include photoautotrophic microorganisms (e.g. Ylla et al. 2012, 2014; Oest et al. 2018; Romero et al. 2019; Dang et al. 2021; Thomas et al. 2021; Zhang et al. 2021), while fewer studies have focused on predominantly heterotrophic microbial communities from the hyporheic zones, i.e. hyporheic biofilms (but see Findlay and Sinsabaugh 2003; Nogaro et al. 2010; Wagner et al. 2014; Mori et al. 2019; Matjašič et al. 2021).
Both benthic and hyporheic biofilms are highly sensitive to changing environmental conditions (Dang et al. 2021). For example, the abundance, taxonomic composition and metabolic activity of aquatic biofilm communities have been shown to depend on environmental factors, such as hydrogeomorphology (Simčič and Mori 2007; Gerbersdorf et al. 2020), quality and quantity of organic matter (e.g. Findlay et al. 2003; Judd et al. 2006; Freixa et al. 2017), availability of inorganic nutrients and terminal electron acceptors (e.g. Baker et al. 2000; Ito et al. 2002), and temperature (e.g. Ylla et al. 2012, 2014; Mori et al. 2019). Temperature increases accelerates processes, such as microbial growth, activity and metabolic rates (Ylla et al. 2012; Mori et al. 2019). For example, Ylla et al. (2012) reported that the biofilm microbial community responded to increased water temperatures by increasing the bacterial cell number, respiratory activity, and extracellular enzyme activity. Alongside temperature, the availability of organic material and nutrients is thought to have the greatest influence on the abundance and activity of heterotrophic microbial communities in most aquatic ecosystems (Hedin 1990; den Heyer and Kalff 1998; de Vicente et al. 2010; Cibic et al. 2019). Dissolved organic matter (DOM), which is the primary substance for bacterial growth and respiration, is introduced into aquatic ecosystems, where organic carbon (DOC) is usually the most common form of DOM (Benner 2003). DOC contains a variety of compounds: from labile simple sugars readily available to microbes to lignin-based refractory molecules resistant to microbial assimilation (Aiken et al. 1985; Volk et al. 1997). The supply of bioavailable DOC has been cited as one of the most important factors controlling the abundance and metabolic activity of heterotrophic bacteria in many streams (e.g. Buffam et al. 2001; Findlay et al. 2003; Romaní et al. 2006; Foulquier et al. 2011; Weigelhofer et al. 2020). For example, Findlay et al. (2003) found significant differences in bacterial growth, oxygen consumption, and enzyme activities in hyporheic bacterial communities exposed to different DOC concentrations. In general, the addition of labile and recalcitrant organic materials increased oxygen consumption, production or both, depending on the background DOM. Moreover, concentrations of inorganic nutrients, particularly nitrogen and phosphorus, are most important in regulating the growth of autotrophic organisms in aquatic ecosystems, while the combined effect of high organic and nutrient loads may also directly affect the abundance of heterotrophic microbial organisms (Cibic et al. 2019).
Based on the occurrence of different microbial communities in sediments from different hyporheic environments (Lowell et al. 2009; Hall et al. 2012; Atashgahi et al. 2015), different structural and functional responses of hyporheic microbial biofilms to changing environmental conditions are expected. For example, bacterial production can only increase after a temperature change if DOM production, uptake, bacterial growth efficiency or a combination of all three factors increases (Kirchman et al. 2005). Furthermore, temperature acclimation adds another level of complexity to the physiological response to temperature by microorganisms in the environment (Hall et al. 2010). Understanding the individual and combined effects of increased temperatures and altered DOC supply as two of the major environmental drivers is therefore necessary to model ecological processes at different levels of biological organisation in aquatic ecosystems (Cross et al. 2015; Dang et al. 2021).
This study aims to quantify the effects and relative importance of temperature changes and elevated DOC on hyporheic biofilms in streams that are exposed to different intensity of human pressures in the catchment. We investigated the response of hyporheic biofilms originating from the two river reaches with distinct trophic status and temperature regimes (i.e., the unpolluted, colder upstream river reach (pristine, reference location, temperature range 6-10 ºC) and agriculturally impacted, warmer downstream river reach (impacted location, temperature range 6-16 ºC)). Since the biofilms at the reference location have a more constrained ecological niche by means of temperature and nutrient availability ranges compared to the biofilms colonizing sediments at the impacted location, we hypothesised that the structure (i.e. microbial biomass – total protein content (TPC)) and functioning (i.e. respiratory electron transport system activity (ETSA) and community-level physiological profiling (CLPP)) of hyporheic biofilms in sediments from the reference stream reach would respond differently by means of intensity to temperatures changes and increased DOC quantities from those collected from the impacted reach due to above mentioned initial inherent differences in biofilm structural characteristics. We hypothesised that the addition of DOC to biofilms in sediments will increase the biomass production and microbial activity and shift the physiological profile only in a case when exposed to temperature within their natural thermal regimes. To test these hypotheses, a mesocosm experiment was carried out in the laboratory under controlled conditions using hyporheic sediments collected in the field from the two study reaches (i.e. reference and impacted). This study contributes to the understanding of how the combined effects of climate change, by means of changes in water temperature regimes (warming, extreme thermal and hydrological events) and increasing human-driven organic pollution, may further affect the metabolism of streams already exposed to varying degrees of anthropogenic impacts in the catchments.
2 Materials and methods
2.1 Sediment collection
Sediments needed for the mesocosm experiment were collected in October 2018 from the gravel bed pre-alpine river Kamniška Bistrica (Slovenia). Two 50 m long river reaches, approximately 20 km apart, were selected for sediment collection: the unpolluted upstream (i.e. reference) reach near Stahovica (Ref; 46°16′ 08,63'' lat, 14°36′18,33'' lon) and the agriculturally impacted (i.e. impacted) downstream reach near Videm (Imp; 46°04′34,04'' lat, 14°38′11,27'' lon). Reported nitrate concentrations in the river water were 1.94 and 8.97 mg NO3 L−1 for the reference and impacted river reach, respectively, accompanied by a low total phosphorus concentration at the reference reach (Ptot, 0.003 mg L−1) and by a higher concentration at the impacted reach (Ptot, 0.05 mg L−1) (EARS) 2018). The reported water temperatures during the year ranged from 5.6 °C to 10.2 °C at reference and from 6.1 °C to 15.7 °C at impacted reaches (Debeljak 2018). Mean annual DOC were 0.96 mg L−1 (± 0.19 mg L−1) and 2.47 mg L−1 (± 0.93 mg L−1) for the reference and impacted study reaches, respectively (unpublished data). More information on the characteristics of the study river is available in Debeljak (2018), Debeljak et al. (2017) and Mori et al. (2019).
At both river reaches, a plastic tube (diameter 30 cm, height 50 cm) was inserted into the riverbed at three sites to obtain spatial replicates within the study reach. First, the larger stones and the upper layer of the sediments (cca. 20 cm) were removed from the tube to access the hyporheic sediments. Extracted hyporheic sediment samples were gently sieved through an 8 mm mesh and then through a 4 mm mesh to obtain sediments with a grain size < 4 mm. Approximately 15 L of homogenised sediment from each study reach was transported to the laboratory where it was used for incubation experiments under controlled conditions.
2.2 Experimental design
Prior to the experimental setup, the grain size distribution was determined for the sediments from both study reaches using a shaker (MINOR, Endecotts, UK) consisting of three sieves (mesh size 0.063 mm, 0.2 mm and 2 mm). The grain size distribution was expressed as a percentage of the total dry mass of the sample for each grain size fraction.
After grain size characterisation, the sieved sediments from the two study reaches were placed in 12 round Plexiglas containers (diameter 21 cm, height 10 cm) and incubated in the dark in temperature-controlled chambers for four weeks (W1, W2, W3, W4) under two temperatures (10 °C and 14 °C) and wetted with three types of synthetic water (control – C; low DOC – lDOC; high DOC – hDOC; N = 2 loc × 2 temp × 3 water types × 1 replicate). 1 L of synthetic water (i.e. 2940 mg of CaCl × 2 H2O, 1230 mg of MgSO4 × 7 H2O, 650 mg of NaHCO3 and 60 mg of KCl in 10 L of bidistilled water) (ISO standard 6341, 1996) was added to four control containers (C). 1 L of synthetic water was added with 5 mg C (2.5 mg C as cellobiose (Sigma-Aldrich) and 2.5 mg C as dipeptide proline-leucine (Sigma-Aldrich)) into four "low DOC" containers (lDOC), whereas 1 L of synthetic water was added to four "high DOC" containers (hDOC) with 30 mg C (15 mg C cellobiose and 15 mg C proline-leucine). An air stone was placed in each container to enable continuous aeration, and the container was covered with dark foil during incubation to prevent evaporation. At the bottom, the containers had two diametrically positioned openings (diameter 0.5 cm) into which a plastic tube with a valve at the end was inserted to regularly collect the water samples for further chemical analyses collected on W1, W2, W3 and W4. Inside the container, the openings were covered with a net to prevent the tubes from clogging when interstitial water was sampled.
2.3 Water sampling and analyses
During the incubation, 100 mL of interstitial water was collected weekly (W1, W2, W3 and W4) for the measurements of physical and chemical characteristics using plastic tubes placed at the bottom of the containers. A temperature, dissolved oxygen concentration, and conductivity were measured using WTW probes (Multi 3430 Set, WTW, Germany). After the measurements, the collected water was poured back into the respective containers. After the first and fourth weeks of incubation, water samples were collected to determine dissolved organic carbon (DOC) and total nitrogen (TN). DOC and TN in water were analysed according to SIST ISO 8245:2000 (filtration through 0.45 μm filter) and SIST EN 12260:2003, respectively.
2.4 Biofilm analyses
Biofilms were analysed prior to the experimental setup and after one, two and four weeks of incubation, when 20 cm3 of sediment was collected from each container in triplicate. After collecting, part of the sediment was immediately used for community-level physiological profile (CLPP) analysis and particulate organic matter (POM) determination, while the other part of the sediment was stored in a plastic bag in the freezer at -80 °C until ETSA and TPC analyses were performed. POM was also determined for sediments prior to the incubation. The amount of POM in the sediments was determined by the loss of ignition. Dried samples (24 h, 105 °C) were ignited at 520 °C for 2 h, and the POM content was determined as loss on ignition in g per dry mass of sediment.
TPC (i.e. microorganisms and extracellular polymeric protein substances) in the sediment samples was determined using a commercially available kit according to the manufacturer's recommendations (Pierce™ BCA Protein Assay Kit, Thermo Scientific). First, 25 µL of the standard solutions or supernatant of samples were pipetted in duplicate into each well of a microtiter plate, and then 200 µL of the working reagent was added to each well. After incubation, absorbance was measured at 562 nm within 10 min using a microplate reader (Synergy Mx, BioTek). The protein content of the sample was calculated using a standard curve and expressed as µg g−1 dry mass of sediment.
Respiratory ETSA was measured using an assay originally proposed by Packard (1971) and modified by Broberg (1985). It estimates potential metabolic activity by determining the activities of dehydrogenases and cytochromes within the respiratory electron transport system. 10 g of sediment was weighed into centrifuge tubes that were stored in ice. To each tube, 3 mL of homogenisation buffer was added, and the sample was ultrasonically homogenised for 2 min (ultrasonic homogeniser, Cole-Palmer). The samples were centrifuged in a refrigerated centrifuge (Sigma) at 10,000 rpm for 4 min. From each sample, 30 µL of the supernatant was pipetted into three wells of a microtiter plate, while the fourth well was left empty and used as a blank. 150 µL of the substrate solution (NADPH, NADH) and 50 µL of the reagent solution (2-(p-iodophenyl)-3-(p-nitrophenyl)-5-phenyltetrazolium chloride, INT) were added to all four wells. The entire procedure was repeated for all samples and then incubated in the dark at 20 °C for 30 min. The reaction was stopped by adding 50 μL of the stop solution (H3PO4 conc.: formalin) to all wells. Finally, another 30 μL of the sample was added to the blank well, and absorbance was measured at 490 nm within 10 min. We calculated the concentration of oxygen consumed, which corresponds to the amount of formazan formed by the reduction of INT. We used the following formula:
ETSA = electron transport system activity (μL O2 S−1 h−1).
Abs490nm = sample absorption (the absorbance of the blank subtracted from the absorbance of the sample).
Vr = final volume of the reaction mixture (0.28 mL).
Vh = volume of homogenate (3 mL).
Va = volume of homogenate in the sample (0.03 mL).
S = sample size (dry mass (DW), g).
t = incubation time (30 min).
1.30 = path length correction factor (Lampinen et al. 2012).
1.42 = factor for conversion of formazan concentration to oxygen volume (Kenner and Ahmed 1975).
CLPP was performed using the Biolog Ecoplates™ assay (Biolog, California, USA), which provides information on community functioning. The Biolog Ecoplate™ consists of 96 wells containing 31 different carbon sources and a control well in three replicates (Weber and Legge 2009). For each well, the redox dye tetrazolium violet was used as a colour indicator to show whether the added microorganisms utilised the substrates (Insam and Goberna 2004). First, one gram of fresh sediment was weighed into a sterilised glass beaker (100 mL) using an analytical balance (Sartorius, Germany), and 10 mL of cold Ringer's solution (Ringer's solution ¼ strength tablets, Sigma-Aldrich) was added. The beakers were placed in an ultrasonic bath (Elma, Germany) for 1 min (37 kHz, 30%). The solution from each beaker was poured into a centrifuge tube and centrifuged at 800 rpm for 5 min at 4 °C. Then, 150 μL of the supernatant of the first replicate was transferred from the vial to each well of one-third of the Biolog Ecoplates™ using a multichannel pipette. The supernatants of the second and third replicates from the same container were transferred to the second and third parts of Biolog Ecoplates™, respectively. The absorbance of each well was measured on the same day and after 24, 48 and 72 h at 590 nm using a SynergyMX spectrofluorometer (BioTek Instruments, USA). Between measurements, the Biolog Ecoplates™ were incubated in the dark at 20 °C. The raw absorbance values measured for the substrate wells on each third of the plate were corrected for the absorbance of the blank well (inoculated but without a carbon source). Negative values were coded as zeroes for subsequent data analysis. The average well colour development (AWCD), which represents the average metabolic activity over all wells, was calculated as
where Ai represents the absorbance reading of well i and A0 is the absorbance reading of the blank well. In accordance with certain recommendations (Weber and Legge 2010), substrate utilisation measured after 48 h of incubation with Biolog Ecoplates™ was used for CLPP.
The figures for the results of the biofilm parameters analysed at the first and second week of incubation are attached as supplementary material.
2.5 Statistical analysis
We used Shapiro–Wilk's test and Levene's test to examine the normality of the data distribution and homoscedasticity. Consequently, all values (TPC, ETSA, AWCD and substrate utilisation absorbances) were log10(x + 1) transformed and proportions for carbon substrate groups were arcsin transformed to ensure normality and homoscedasticity. First, a t-test was used to test for the differences in measured variables between sediments from reference and impacted study reaches or sediments exposed to two temperatures, and a one-way ANOVA was applied to test for the differences in the three DOC regimes. To examine the effects of the study reach (Reference, Impacted), the two temperatures (10 °C and 14 °C) and three DOC regimes (C, lDOC and hDOC) on TPC, ETSA and AWCD, the utilisation of carbon substrate groups, and three-way ANOVAs were performed on data obtained after one week, two weeks, and four weeks of incubation, respectively, with location (Loc), temperature (Temp), and quality (DOC) as factors. Tukey's honest significant difference post hoc test was used to determine the significance of the factors and interactions.
The CLPP data were investigated using the non-metric multidimensional scaling (NMDS) analysis method based on the Bray–Curtis similarity index to visualise and test for differences in overall metabolic function between samples. Input data for NMDS included absorbance data from all treatments (in triplicate), separately for each week (W1, W2 and W4). One-way (between sediments from the reference and impacted study reaches) and two-way analyses of similarity (ANOSIM) were performed to test for the significance of differences in substrate utilisation separately between sediments from the two study reaches (Reference, Impacted) and incubation temperatures (10 °C and 14 °C), the two locations and water quality (C, lDOC, hDOC), and incubated temperatures and water quality as factors. Similarity percentage analysis (SIMPER) was used to examine the substrate utilisation that contributed most to differences between locations, incubation temperatures, and water quality separately for each week (W1, W2 and W4). Statistical analyses (three-way ANOVA, t-test, Shapiro–Wilk's test and Levene's test) were performed using SPSS Statistics version 20.0 for Windows (SPSS Inc., Chicago, IL, USA), while NMDS, ANOSIM and SIMPER were performed using PAST 3.22 (Hammer et al. 2001).
3 Results
3.1 Physical and chemical characteristics of water and sediments prior and during incubation
The grain size distribution of the sediments from both sampling locations prior to the experimental setup was: 8% and 9% of grain size fraction < 0.063 mm, 63% and 61% of grain size fraction 0.2–2 mm, and 20% and 22% of grain size fraction 2–4 mm, respectively, for the sediments from the reference and impacted study reaches. Prior the incubation, the sediments from the reference study reach had a significantly higher content of initial POM (9.1 ± 1.1 g kg−1) than those from the impacted stream reach (7.1 ± 0.7 g kg−1) (N = 6, t-test, p ≤ 0.05).
During incubation, however, no clear pattern in POM related to controlled parameters (location, temperature, DOC) was observed (Table 1). The lowest average oxygen concentrations in the interstitial water of both stream sediments were measured at 14 °C and high DOC enrichment indicating intensive oxygen consumption due to higher temperature and the high availability of food resources. Generally, higher temperature and DOC enrichment let to lower oxygen concentrations in the sediments from both study reaches, and significant differences between the two study reaches were observed for the incubation at 10 °C but not at 14 °C (N = 24, t-test, p ≤ 0.05) (Table 1). Substantially increased conductivity was observed in the sediments from both locations after incubation at 14 °C and under treatment with hDOC water (Table 1). In contrast, concentrations of TN in the interstitial water showed no clear pattern related to tested factors (e.g. location, temperature and DOC), most probably due to the low number of samples.
3.2 Total protein content (TPC) and electron transport system activity (ETSA)
The biomass of the biofilms, measured as TPC in sediments from the two locations prior the incubation significantly differed and was lower (13.4 ± 1.3 µg g−1) for the reference location than that from the impacted location (24.9 ± 2.7 µg g−1) (t-test, p < 0.01). At the end of the experiment (i.e., after four weeks), the average TPC (N = 9) was 46.9 µg g−1for the sediments from reference and 51.2 µg g−1for the sediments from impacted location incubated at 10 °C (Fig. 1a, b). At 10 °C, hDOC enrichment resulted in increased TPC in sediments from reference, but not in those from impacted location. The average TPC at 14 °C was 19.2 and 84.0 µg g−1 for the sediments from the reference and impacted locations, respectively (Fig. 1a, b). Relatively low TPC was measured at 14 °C after DOC enrichment in sediments from reference location, while in biofilms from impacted location TPC increased with DOC enrichment. According to the three-way ANOVA (factors: location, temperature, DOC), significantly higher TPC values were found in sediments from the impacted location after four weeks of incubation, and also after one and two weeks (Figs. S1-S2; Table 2). Higher TPC values were found in sediments with high DOC than in those with low DOC after two and four weeks of incubation (Table 2). After four weeks, the high DOC treatment showed also significantly higher TPC values than the control (Tukey's post hoc test, p < 0.05). Surprisingly, temperature did not prove to be a significant factor for differences in TPC, but there was a significant interaction between location and temperature on TPC after one and four weeks of incubation (Table 2).

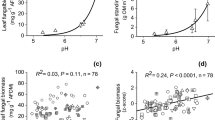
Total protein content – TPC (a, b), electron transport system activity – ETSA (c, d), average well colour development – AWCD (e, f) in the sediments from reference and impacted locations incubated at 10 or 14 °C with no addition of DOC (control – C), addition of 5 mg of DOC L−1 (lDOC) or 30 mg DOC L.−1 (hDOC) measured after four weeks of incubation. Bars represent means ± SD (n = 3). Different letters in the column indicate significant differences between treatments (p < 0.05)
ETSA differed between the sediments from the reference and impacted locations prior the incubation. Significantly lower ETSA (0.50 ± 0.03 µLO2 g−1DW h−1) was determined in the sediments from reference location than in those from the impacted location (1.16 ± 0.07 µLO2 g−1DW h−1) (t-test, p < 0.01). At the end of the incubation at 10 °C, the average ETSA was 0.49 and 1.01 µLO2 g−1DW h−1 for the sediments from the reference and impacted locations, respectively (Fig. 1c, d), indicating no effects of temperature on ETSA. The addition of DOC at 10 °C did not lead to an increase in ETSA in the sediments from reference locations, but it increased slightly in impacted sediments. The average ETSA after four weeks of incubation at 14 °C was 0.55 and 1.39 µLO2 g−1DW h−1 for the sediments from the reference and impacted locations, respectively (Fig. 1c, d). Results showed that increased ETSA was observed at 14 °C and for hDOC enrichment. A three-way analysis of variance (ANOVA) indicated location, temperature and DOC as significant factors for differences in ETSA but not for all sampling dates (Table 2). For example, location was a significant factor on all sampling occasions after four weeks, while temperature was significant after two and four weeks and DOC was significant after one week of incubation. Interactions (Loc × Temp; Loc × DOC) were observed only for data obtained after one week of incubation (Table 2).
3.3 Community-level physiological profiling
At the beginning of the incubation, differences in microbial functioning were observed between the sediments collected at reference and impacted locations. Significantly lower AWCD values were determined in the sediments from the reference (0.42 ± 0.01) than in those from the impacted location (0.89 ± 0.11) (t-test, p < 0.05). Utilisation of alfa-D-lactose, alfa-cyclodextrin, D-cellobiose and glucose-1-phosphate contributed most to dissimilarity between reference and impacted locations, but differences were not statistically significant (ANOSIM, p > 0.05). The utilisation of carbon substrate groups—carbohydrates, polymers, carboxylic and ketonic acids, phenolic compounds, amino acids and amines/amides—did not differ significantly between sediments from reference and impacted locations prior the start of the incubation (t-test, p > 0.05). Among the groups, the microbes efficiently utilised the carbohydrates, while a low percentage of utilisation was observed for the phenolic compounds.
At the end of the experiment (i.e., after four weeks), the AWCD values were still lower in the sediments of the reference location than those of the impacted location (Fig. 1e, f). The average AWCD after four weeks of incubation at 10 °C was 0.70 for the sediments from the reference locations and 0.81 for the impacted sediments. No pattern was observed in the responses of AWCD to DOC addition in sediments from reference or impacted location at 10 °C. At 14 °C, average AWCD was 0.74 and 0.87 for the biofilms in reference and impacted sediments, respectively, where DOC enrichment resulted in increased AWCD. A three-way ANOVA indicated a significant impact of location, temperature and DOC after four weeks, whereas earlier during incubation (at the first and second week; Fig. S1-S2), temperature have no impact on AWCD (Table 2). A significant interaction between location and DOC was obtained after four weeks and between temperature and DOC after two and four weeks (Table 2).
Comparison of microbial community functioning using NMDS showed differential substrate utilisation between sediments from reference and impacted locations at after four weeks (Fig. 2; stress = 0.1808), and the similar results were obtained for the first week (Fig. S3; stress = 0.1056), and second week (Fig. S4; stress = 0.1501). Moreover, after four weeks, greater variability under different treatments was observed for hyporheic biofilms from the reference study reach, particularly due to the pronounced effect of DOC addition on substrate utilisation. Substrate utilisation of hyporheic biofilms from the impacted location was less variable after incubation in different temperature and DOC regimes. ANOSIM showed the importance of the location and addition of DOC for the differences in substrate utilisation available on Biolog Ecoplates™, but no significant effect of temperature was detected after four weeks, and also not after first and second week (Table 3). The SIMPER analysis showed that the substrates that contributed most to the differences between locations, temperature and the various DOC additions after four weeks were alfa-D-lactose, alfa-cyclodextrin, glucose-1-phosphate, and 4-hydroxybenzoic acid (Fig. 3). In addition to the above substrates, L-arginine contributed more than 5% to the differences at first week Fig. S5) and 2-hydroxy benzoic acid and phenylethyl-amine contributed more than 5% to the differences at second week (Fig. S6).

Ordination diagrams of NMDS based on data on substrate utilisation (Biolog Ecoplates™) after four weeks of incubation (stress = 0.1808) in the sediments from reference (blue colour) and impacted (orange colour) locations incubated at 10 °C (empty marks) and 14 °C (full marks) with no addition of DOC (control—C; ◊), low DOC (lDOC; ∆) and high DOC (hDOC; □)

Heatmap indicating the intensity of substrate utilisation on Biolog Ecoplates™ in the sediments from reference and impacted locations incubated at 10 or 14 °C with no addition of DOC (control – C), addition of 5 mg DOC L−1 (lDOC) or 30 mg DOC L−1 (hDOC) at the beginning of incubation (Initial) and after four weeks of incubation. The green–yellow–red scale was used; the darkest green colour indicates the log maximal substrate utilisation (absmax = 0.6) and the darkest red indicates the minimal substrate utilisation (absmin = 0), expressed as absorbance (abs) and measured spectrophotometrically
At the end of the experiment (i.e., after four weeks) the average percentage for carbohydrates at 10 °C was 39.0% and 36.6% for the sediments from the reference and impacted locations, respectively (Fig. 4). Addition of hDOC increased percentage of carbohydrate utilisation in sediments from reference locations, but not impacted ones. The average percentage for carbohydrates utilisation at 14 °C was 36.6% and 36.0% for the sediments from the reference and impacted locations, respectively. DOC enrichment did not affect percentage of carbohydrate utilisation in hyporheic biofilms of impacted sediments. According to three-way ANOVA (Table 4), location as a factor had a significant effect on the proportion of carbohydrate, polymer, phenolic compound and amine/amide utilisation, being higher in the sediments from reference locations for the first two groups and in the sediments from impacted locations for the last two groups at the fourth week, and also at the first and second week (Fig. S7-S8). The proportion of utilised carbohydrates was higher at 10 °C than at 14 °C at the end of the experiment. Moreover, the utilisation percentages of carbohydrates were significantly affected by different DOC additions (lower for lDOC than hDOC and control) and temperature, where a higher proportion was obtained at 10 °C. At the end of the experiment, interaction between location and DOC was observed for carbohydrates.

The utilisation of carbon substrate groups in different treatments at the beginning of incubation (Initial) and after four weeks of incubation in the sediments from reference and impacted locations. Abbreviation represents incubation temperature (10 °C or 14 °C), and nutrient level (no addition of DOC (control – C), addition of 5 mg DOC L−1 (lDOC) or addition of 30 mg DOC L.−1 (hDOC))
4 Discussion
In general, changes in environmental factors such as water temperature and inputs of nutrients and organic substrates affect aquatic microbial communities through both changes in diversity and metabolic activity, and thus through structural and functional responses (Besemer 2015). However, the intensity and the mode of the response of hyporheic biofilms under specific environmental conditions is pre-determined by existing microbial community composition, abundances, biomass and age of biofilm (e.g. Ylla et al. 2012, 2014; Phillips et al. 2017; Mori et al. 2019). This was confirmed also by our study which showed that the microbial communities in the hyporheic sediments from colder, unpolluted (i.e. reference) locations responded more strongly to DOC enrichment at a lower temperature of 10 °C and very little under 14 ºC, which is a temperature beyond the range occurring in the environment from where the sediments used in the study originated. On the other hand, the hyporheic biofilms already acclimated to both experimental temperatures (10 and 14 ºC) and eutrophication responded very intensively to DOC enrichments at both temperatures, but with more accelerated response at higher temperature,
Higher initial TPC (Fig. 1b) measured prior to the experiment in sediments from polluted locations was in accordance with the findings of Mori et al. (2019), who reported that higher TPC is related to a higher proportion of urban and agricultural land use in the catchment and of Simčič et al. (2022) reporting increased TPC in the environments with increased nutrients and organic matter contents. A similar response was observed for ETSA (Fig. 1c,d). ETSA has been frequently used as a functional measure of the intensity of organic matter decomposition and mineralisation in aquatic sediments (e.g. Trevors 1984; Fleituch and Leichtfried 2007; Simčič and Mori 2007; Ylla et al. 2012; Mori et al. 2017, 2019; Simčič et al. 2022). It depends on the trophic conditions of the aquatic ecosystem and increases with increasing trophic status (Debeljak et al. 2017).
It is well known that increasing temperature accelerates biological processes, including microbial growth rate (Phillips et al. 2017) and metabolic activity (Mori et al. 2019). However, the results of the present study show that temperature has no significant effect on hyporheic biofilm biomass, but a slight decrease in biomass was observed during temperature change from 10 ºC to 14 ºC for biofilms from the reference location. It seems that incubation at 14 °C represented stress for hyporheic biofilms from the reference location, even when enriched with DOC, as they are adapted to lower temperatures and without the ability to adjust temperature changes (Hall et al. 2010). Furthermore, Mori et al. (2019) found that water temperature is irrelevant for predicting the response of structural indicators in models that consider the effects of multiple stressors. This means that other stressors can override the effects of temperature. In contrast to TPC, ETSA was significantly higher in sediments incubated at higher temperatures, as reported by Ylla et al. (2012). Similarly, ETSA was shown to be significantly related with temperature when conducting a regional study of hyporheic biofilms' response to multiple stressors (Mori et al. 2019). Furthermore, elevated temperatures accelerated the degradation of recalcitrant (mainly allochthonous) compounds more than the degradation of labile compounds (Ylla et al. 2012). Thus, the extremely high DOC concentrations observed in treatments with high DOC enrichment at high temperatures at the end of incubation are likely related to the high ETSA and microbial biofilm decomposition activity of recalcitrant organic matter (i.e. POM) in the sediments. In addition, the higher DOC in sediments from reference locations than in sediments from impacted locations, both incubated at 14 °C with hDOC at fourth week (393 mg L−1 and 287 mg L−1 for reference and impacted locations, respectively) could be related to the higher amount of POM in the sediments from the reference location compared with the second one with impacted sediments. One of the reasons for the higher POM in the reference sediments, which are intensively degraded during incubation at 14 °C, is probably the lower water temperature in the upper reaches of the river where the reference location is located. This result is in accord with the findings of Freixa et al. (2017), who reported that small temperature changes can have a strong impact on carbon metabolism by increasing heterotrophic activity and accelerating carbon cycling, as well as altering sediment biofilm communities, ultimately affecting the composition of the food web in rivers.
The abundance and metabolic activity of heterotrophic bacteria are controlled by the supply of bioavailable DOC (e.g. Weigelhofer et al. 2020). Accordingly, the addition of DOC resulted in an increase in TPC relative to the amount of DOC added, as the highest TPC values were obtained in sediments with the highest DOC addition, which is in agreement with the results of the study by Ylla et al. (2012), where the addition of labile DOC increased the heterotrophic biomass of the biofilm. Furthermore, the higher TPC values in the sediments from the impacted location are consistent with the results of Cibic et al. (2019), who reported that the abundance of heterotrophic microbial organisms was stimulated by the combined effect of high organic and nutrient loads. The microbial community background was also reflected in the higher ETSA responses of the biofilm in the sediments from the impacted locations compared to those from the reference locations. In addition, the biofilm response also depended on the amount of DOC added. The reason for the higher ETSA in sediments incubated with high DOC enrichment than in sediments with low DOC enrichment is likely the greater availability of labile DOC for the metabolic processes of heterotrophic microbial biofilms that require organic carbon as an energy and food source (Bechtold et al. 2012), which was limited under natural conditions (Romaní et al. 2006). In the present study, high conductivity was observed for all treatments with biofilms from impacted locations as well as for treatments with biofilms from reference locations with hDOC enrichment at 14 °C. In addition to the high conductivity, a high ETSA of the biofilms was present. This is consistent with the results of Debeljak et al. (2017), in which conductivity and DOC were identified as the main drivers of ETSA in five pre-Alpine rivers in Slovenia over two seasons (summer and winter) and at two depths (5–15 and 20–40 cm).
A more complex measure describing microbial functioning is community-level physiological profiling (CLPP) using Biolog Ecoplates™ (Biolog, California, USA), which has been used in several studies to examine the effects of sediment origin (e.g. Oest et al. 2018; Thomas et al. 2021), temperature (e.g. Biggs et al. 2011; Ylla et al. 2014; Dadi et al. 2017), or nutrients (e.g. Kiersztyn et al. 2019; Lacoste et al. 2019) on the utilisation of different substrates to determine the functional diversity of microbial communities. AWCD, which represents the average metabolic activity over 31 wells with carbon substrates on Biolog Ecoplates™, was influenced by sediment origin, revealing higher AWCD in sediments from impacted locations than at reference locations at all time points (i.e., after one, two and four weeks). The reason for higher AWCD for biofilm in impacted sediments is higher microbial abundance since Christian and Lind (2006) found a significant positive correlation between bacterial inoculum density and AWCD. Similarly, Liao et al. (2018) reported that anthropogenic activities promoted bacterial production, influenced the distribution of dominant species and accelerated the metabolic rate of organic matter by microbes. In addition to sediment origin, DOC enrichment and temperature as factors significantly affect AWCD after four weeks, with high DOC enrichment and elevated temperature leading to higher AWCD, as also reported by Dadi et al. (2017). However, the addition of DOC resulted in different responses in the AWCD with respect to the locations, with lower values after the addition of DOC to the biofilms from the reference location compared to the control without DOC addition, while the AWCD after the addition of hDOC was higher in the sediments of the impacted location than in the control. Moreover, the significant interaction between temperature and DOC treatment showed a higher AWCD in the control at 10 °C but a higher value in the hDOC treatment at 14 °C, indicating the importance of the synergistic interaction between the studied factors for carbon substrate utilisation.
The response of biofilms from the reference and impacted locations indicates the importance of the origin of microbial communities for the utilisation of organic compounds. Because environmental factors vary across stream regions, it seems plausible that the general composition of organic matter and consequently the microbial community and its metabolic function varies across the studied stream regions (Findlay and Sinsabaugh 2006). The response of biofilm to multiple stressors depends on the ability of its organisms to respond to each stressor as well as potential interactions (Romero et al. 2019). When 31 carbon substrates were considered separately, the results of our study showed that the profiles of carbon substrate utilisation were influenced by location and DOC enrichment but not by temperature (Table 3). The utilisation of four carbon substrate groups, namely carbohydrates, polymers, phenolic compounds and amines/amides were all affected by sediment origin, while the utilisation of carbohydrates was also significantly affected by DOC enrichment and temperature after four weeks (Table 4). It is known that carbohydrates are the preferred metabolic pathway of many aerobic and facultative heterotrophs involving the oxidation of a simple or complex carbohydrate and oxygen as the terminal electron acceptor (Oest et al. 2018). These carbohydrates are utilised to a greater extent at lower temperatures because more bioavailable organic matter is rapidly utilised regardless of higher temperatures, while the degradation of recalcitrant substances is enhanced by warming (Ylla et al. 2012). In addition, higher proportions of carbohydrates and polymers were utilised in the sediments from the reference location, while higher utilisation of phenolic compounds and amines/amides was observed in the impacted sediments (Table 4), indicating a different microbial community structure in the sediments from the two study reaches (i.e. reference and impacted). For example, 4-hydroxybenzoic acid, one of the phenolic compounds on Biolog Ecoplates™ used as an intermediate in the synthesis of agrochemicals (pesticides) (OECD SIDS 1999), is extensively utilised by biofilms in sediments from agriculturally impacted locations. Consistent with this, Langenheder et al. (2006) reported that the history and distribution of taxa within the source community (i.e. inoculum) were important factors in the composition of the selected communities, which differed in terms of specific functions, such as enzyme activities, whereas environmental conditions (i.e. medium) appeared to be less important when bacteria from distinctly different aquatic habitats were regrown under identical conditions. To gain a deeper understanding of the structural–functional linkage of hyporheic biofilms, further investigation of the taxonomic composition of the community using next-generation sequencing (NGS) would be a next step in studies such as ours. Several studies have already reported distinct microbial community composition along anthropogenic pressure gradients (Liu et al. 2018; Liao et al. 2019), and some have also provided insights into structural–functional linkages (Li et al. 2021).
5 Conclusion
This study provides new information on the response of hyporheic biofilms from different environments to temperature changes and enrichment with DOC. As shown, site-specific differences in biofilm features are reflected in the different responses of structural (i.e. TPC) and functional (ETSA, CLPP) measures. In general, higher values for all study measures were observed in biofilms from urban and agriculturally polluted locations, which increased more intensely at higher temperatures and DOC enrichment than those from unpolluted reference locations. Lower microbial biomass was observed in the sediments from the reference location when exposed to higher temperatures, most likely due to a stress response to high temperatures, as the environment from which the biofilm originated has lower water temperatures ranges (6 -10 ºC). After four weeks of incubation, the CLPP exhibited the high diversity among location, temperature and DOC regimes for carbohydrate utilisation and less diversity in other substrates utilization (e.g. amino acids). Our study demonstrates that site-specific environmental conditions, such as trophic status and/or existing temperature regime, strongly underline the response of the hyporheic biofilm structure and its functioning to additional stressors.
Data availability
The data that support the findings of this study are available from the corresponding author on reasonable request.
References
Aiken GR, McKnight DM, Wershaw RL, MacCarthy P (eds) (1985) Humic Substances in Soil, Sediment, and Water: Geochemistry, Isolation, and Characterization. Wiley-Interscience, New York
Atashgahi S, Aydin R, Dimitrov M et al (2015) Impact of a wastewater treatment plant on microbial community composition and function in a hyporheic zone of a eutrophic river. Sci Rep 5:17284. https://doi.org/10.1038/srep17284
Baker MA, Dahm CN, Valett HM (2000) Organic carbon supply and metabolism in a shallow groundwater ecosystem. Ecology 81:3133–3148. https://doi.org/10.1890/0012-9658(2000)081[3133:OCSAMI]2.0.CO;2
Battin TJ, Besemer K, Bengtsson MM, Romani AM, Packmann AI (2016) The ecology and biogeochemistry of stream biofilms. Nat Rev Microbiol 14:251–263. https://doi.org/10.1038/nrmicro.2016.15
Bechtold HA, Marcarelli AM, Baxter CV, Inouye RS (2012) Effects of N, P, and organic carbon on stream biofilm nutrient limitation and uptake in a semi-arid watershed. Limnol Oceanogr 57:1544–1554. https://doi.org/10.4319/lo.2012.57.5.1544
Benner R (2003) Molecular indicators of the bioavailability of dissolved organic matter. In: Findlay S, Sinsabaugh RL (eds) Aquatic ecosystems: Interactivity of dissolved organic matter. Academic Press, San Diego, California, USA, pp 121–137
Besemer K (2015) Biodiversity, community structure and function of biofilms in stream ecosystems. Res Microbiol 166:774–781. https://doi.org/10.1016/j.resmic.2015.05.006
Biggs CA, Olaleye OI, Jeanmeure LF, Deines P, Jensen HS, Tait SJ, Wright PC (2011) Effect of temperature on the substrate utilization profiles of microbial communities in different sewer sediments. Environ Technol 32:133–144. https://doi.org/10.1080/09593330.2010.490852
Broberg A (1985) A modified method for studies of electron transport system activity in freshwater sediments. Hydrobiologia 120:181–187. https://doi.org/10.1007/BF00032140
Buffam I, Galloway JN, Blum LK, McGlathery KJ (2001) A stormflow/baseflow comparison of dissolved organic matter concentrations and bioavailability in an Appalachian stream. Biogeochemistry 53:269–306. https://doi.org/10.1023/A:1010643432253
Christian BW, Lind OT (2006) Key issues concerning Biolog use for aerobic and anaerobic freshwater bacterial community-level physiological profiling. Int Rev Hydrobiol 91:257–268. https://doi.org/10.1002/iroh.200510838
Cibic T, Fazi S, Nasi F, Pin L, Alvisi F, Berto D, Viganòe L, Zoppinib A, Del Negro P (2019) Natural and anthropogenic disturbances shape benthic phototrophic and heterotrophic microbial communities in the Po River Delta system. Estuar Coast Shelf Sci 222:168–182. https://doi.org/10.1016/j.ecss.2019.04.009
Cross W, Hood JM, Benstead JP, Huryn AD, Nelson D (2015) Interactions between temperature and nutrients across levels of ecological organization. Glob Chang Biol 21:1025–1040. https://doi.org/10.1111/gcb.12809
Dadi T, Wendt-Potthoff K, Koschorreck M (2017) Sediment resuspension effects on dissolved organic carbon fluxes and microbial metabolic potentials in reservoirs. Aquat Sci 79:749–764. https://doi.org/10.1007/s00027-017-0533-4
Dang C, Kellner E, Martin G, Freedman ZB, Hubbart J, Stephan K, Kelly CN, Morrissey EM (2021) Land use intensification destabilizes stream microbial biodiversity and decreases metabolic efficiency. Sci Total Environ 767:145440. https://doi.org/10.1016/j.scitotenv.2021.145440
de Vicente I, Amores V, Guerrero F, Cruz-Pizarro L (2010) Contrasting factors controlling microbial respiratory activity in the sediment of two adjacent Mediterranean wetlands. Naturwissenschaften 97:627–635. https://doi.org/10.1007/s00114-010-0678-7
Debeljak B, Simčič T, Ciglič R, Pflieger M, Mori N (2017) Spatio-temporal variation in microbial respiration in the shallow hyporheic zone of pre-alpine rivers related to catchment land use. Fundam Appl Limnol 190:265–277. https://doi.org/10.1127/fal/2017/0962
Debeljak B (2018) Structure and functioning of the hyporheic zones in the gravel-beds of five rivers in relation to catchment land use. Dissertation, University of Nova Gorica. https://repozitorij.ung.si/Dokument.php?id=17203&lang=slv. Accessed 12 Jan 2024
den Heyer C, Kalff J (1998) Organic matter mineralization rates in sediments: a within- and among-lake study. Limnol Oceanogr 43:695–705. https://doi.org/10.4319/lo.1998.43.4.0695
EARS (2018) Rezultati monitoringa ekološkega stanja vodotokov v letu 2017 (Results of ecological monitoring of watercourses in 2017) https://www.arso.gov.si/vode/reke/ocena%20stanja/Ekolosko_stanje_reke_2017.pdf. Accessed 22 Dec 2022
SIST EN 12260:2003 Water quality - Determination of nitrogen - Determination of bound nitrogen after oxidation to nitrogen oxides. European Committee for Standardization, Brussels
Findlay S, Sinsabaugh RL (2003) Response of hyporheic biofilm metabolism and community structure to nitrogen amendments. Aquat Microb Ecol 33:127–136. https://doi.org/10.3354/ame033127
Findlay SEG, Sinsabaugh RL, Sobczak WV, Hoostal M (2003) Metabolic and structural response of hyporheic microbial communities to variations in supply of dissolved organic matter. Limnol Oceanog 48:1608–1617. https://doi.org/10.4319/lo.2003.48.4.1608
Findlay S, Sinsabaugh RL (2006) Large-scale variation in subsurface stream biofilms: A cross-regional comparison of metabolic function and community similarity. Microb Ecol 52:491–500 (http://www.jstor.org/stable/25153402)
Fleituch T, Leichtfried M (2007) Electron transport system (ETS) activity in alder leaf litter in two contrasting headwater streams. Int Rev Hydrobiol 92:378–391. https://doi.org/10.1002/iroh.200610986
Foulquier A, Mermillod-Blondin F, Malard F, Gibert J (2011) Response of sediment biofilm to increased dissolved organic carbon supply in groundwater artificially recharged with stormwater. J Soils Sediments 11:382–393. https://doi.org/10.1007/s11368-010-0323-2
Freixa A, Acuña V, Casellas M, Pecheva S, Romaní AM (2017) Warmer night-time temperature promotes microbial heterotrophic activity and modifies stream sediment community. Glob Chang Biol 23:3825–3837. https://doi.org/10.1111/gcb.13664
Gerbersdorf SU, Koca K, de Beer D, Chennu A, Noss C, Risse-Buhl U, Weitere M, Eiff O, Wagner M, Aberle J, Schweikert M, Terheiden K (2020) Exploring flow-biofilm-sediment interactions: Assessment of current status and future challenges. Water Res 185:116182
Hall EK, Singer GA, Kainz MJ, Lennon JT (2010) Evidence for a temperature acclimation mechanism in bacteria: an empirical test of a membrane-mediated trade-off. Funct Ecol 24:898–908. https://doi.org/10.1111/j.1365-2435.2010.01707.x
Hall EK, Besemer K, Kohl L, Preiler C, Riedel K, Schneider T, Wanek W, Battin TJ (2012) Effects of resource chemistry on the composition and function of stream hyporheic biofilms. Front Microbiol 3:35. https://doi.org/10.3389/fmicb.2012.00035
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: paleontological Statistics software package for education and data analysis, 3.22 ed. Palaentologia Electronica 4(1): 9p. http://palaeo-electronica.org/2001_1/past/issue1_01.htm
Hedin LO (1990) Factors controlling sediment community respiration in woodland stream ecosystem. Oikos 57:94–105. https://doi.org/10.2307/3565742
Insam H, Goberna M (2004) Use of Biolog® for Community Level Physiological Profiling (CLPP) of Environmental Samples. In Molecular Microbial Ecology Manual, 2nd edn. Kluwer Academic, Dordrecht, pp 853–860
SIST ISO 8245:2000 Water quality - Guidelines for the determination of total organic carbon (TOC) and dissolved organic carbon (DOC). International Organization for Standardization, Geneva
ISO-standard 6341 (1996) Determination of the inhibition of the mobility of Daphnia magna Straus (Cladocera, Crustacea)-Acute toxicity test, 3rd edn. International Organization for Standardization, Geneva
Ito T, Okabe S, Satoh H, Watanabe Y (2002) Successional development of sulfate-reducing bacterial populations and their activities in a wastewater biofilm growing under microaerophilic conditions. Appl Environ Microbiol 68:1392–1402. https://doi.org/10.1128/AEM.68.3.1392-1402.2002
Judd KE, Crump BC, Kling GW (2006) Variation in dissolved organic matter controls bacterial production and community composition. Ecology 87:2068–2079. https://doi.org/10.1890/0012-9658(2006)87[2068:VIDOMC]2.0.CO;2
Kenner RA, Ahmed SI (1975) Measurements of electron transport activities in marine phytoplankton. Mar Biol 33:119–127. https://doi.org/10.1007/BF00390716
Kiersztyn B, Chróst R, Kaliński T, Siuda W, Bukowska A, Kowalczyk G, Grabowska K (2019) Structural and functional microbial diversity along a eutrophication gradient of interconnected lakes undergoing anthropopressure. Sci Rep 9:11144. https://doi.org/10.1038/s41598-019-47577-8
Kirchman DL, Malmstrom RR, Cottrell MT (2005) Control of bacterial growth by temperature and organic matter in the Western Arctic. Deep Sea Res Part II Top Stud Oceanogr 52:3386–3395. https://doi.org/10.1016/j.dsr2.2005.09.005
Lacoste É, Weise AM, Lavoie MF, Archambault P, McKindsey CW (2019) Changes in infaunal assemblage structure influence nutrient fluxes in sediment enriched by mussel biodeposition. Sci Total Environ 692:39–48. https://doi.org/10.1016/j.scitotenv.2019.07.235
Lampinen J, Raitio M, Perälä A, Oranen H, Harinen RR (2012) Microplate based pathlength correction method for photometric DNA quantification assay. Vantaa: SP&A Application Laboratory Thermo Fischer Scientific. Retrieved from: https://tools.thermofisher.com/ontent/sfs/brochures/AN-SkanIT-Microplate-Based-Pathlength-Correction-Technical-Note-EN.pdf
Langenheder S, Lindström ES, Tranvik LJ (2006) Structure and function of bacterial communities emerging from different sources under identical conditions. Appl Environ Microbiol 72:212–220. https://doi.org/10.1128/AEM.72.1.212-220.2006
Li C, Wang L, Ji S, Chang M, Wang L, Gan Y, Liu J (2021) The ecology of the plastisphere: microbial composition, function, assembly, and network in the freshwater and seawater ecosystems. Water Res 202:117428. https://doi.org/10.1016/j.watres.2021.117428
Liao K, Bai Y, Huo Y, Jian Z, Hu W, Zhao C, Qu J (2018) Integrating microbial biomass, composition and function to discern the level of anthropogenic activity in a river ecosystem. Environ Int 116:147–155. https://doi.org/10.1016/j.envint.2018.04.003
Liao H, Yu K, Duan Y, Ning Z, Li B, He L, Liu C (2019) Profiling microbial communities in a watershed undergoing intensive anthropogenic activities. Sci Total Environ 647:1137–1147. https://doi.org/10.1016/j.scitotenv.2018.08.103
Liu J, Chen X, Shu HY, Lin XR, Zhou QX, Bramryd T, Shu W, Huang LN (2018) Microbial community structure and function in sediments from e-waste contaminated rivers at Guiyu area of China. Environ Pollut 235:171–179. https://doi.org/10.1016/j.envpol.2017.12.008
Lowell JL, Gordon N, Engstrom D, Stanford JA, Holben WE, Gannon JE (2009) Habitat heterogeneity and associated microbial community structure in a small-scale floodplain hyporheic flow path. Microb Ecol 58:611–620. https://doi.org/10.1007/s00248-009-9525-9
Matjašič T, Simčič T, Kanduč T, Samardžija Z, Mori N (2021) Presence of polyethylene terephthalate (PET) fibers in hyporheic zone alters colonization patterns and seasonal dynamics of biofilm metabolic functioning. Wat Res 203:117455. https://doi.org/10.1016/j.watres.2021.117455
Michel A, Schaefli B, Wever N, Zekollari H, Lehning M, Huwald H (2022) Future water temperature of rivers in Switzerland under climate change investigated with physics-based models. Hydrol Earth Syst Sci 26:1063–1087. https://doi.org/10.5194/hess-26-1063-2022
Mori N, Simčič T, Brancelj A, Robinson CT, Doering M (2017) Spatio-temporal heterogeneity of actual and potential respiration in two contrasting floodplains. Hydrol Process 31:2622–2636. https://doi.org/10.1002/hyp.11211
Mori N, Debeljak B, Škerjanec M, Simčič T, Kanduč T, Brancelj A (2019) Modelling the effects of multiple stressors on respiration and microbial biomass in the hyporheic zone using decision trees. Water Res 149:9–20. https://doi.org/10.1016/j.watres.2018.10.093
Nogaro G, Datry T, Mermillod-Blondin F, Descloux S, Montuelle B (2010) Influence of streambed sediment clogging on microbial processes in the hyporheic zone. Freshw Biol 2010:1288–1302. https://doi.org/10.1111/j.1365-2427.2009.02352.x
Oest A, Alsaffar A, Fenner M, Azzopardi D, Tiquia-Arashiro SM (2018) Patterns of change in metabolic capabilities of sediment microbial communities in river and lake ecosystems. Int J Microbiol 2018:6234931. https://doi.org/10.1155/2018/6234931
Orghidan T (1959) Ein neuer Lebensraum des Unterirdischen Wassers der hyporheischen Biotope. Arch Hydrobiol 55:392–414
Orghidan T (2010) New habitat of subsurface waters: the hyporheic biotope. Fundam Appl Limnol / Arch Hydrobiol 176:291–302. https://doi.org/10.1127/1863-9135/2010/0176-0291
Packard TT (1971) The measurement of respiratory electron-transport activity in marine phytoplankton. J Mar Res 29:235–244
Phillips KN, Godwin CM, Cotner JB (2017) The effects of nutrient imbalances and temperature on the biomass stoichiometry of freshwater bacteria. Front Microbiol 8:1692. https://doi.org/10.3389/fmicb.2017.01692
Romaní AM, Vázquez E, Butturini A (2006) Microbial availability and size fractionation of dissolved organic carbon after drought in an intermittent stream: biogeochemical link across the stream–riparian interface. Microb Ecol 52:501–512. https://doi.org/10.1007/s00248-006-9112-2
Romero F, Acuña V, Font C, Freixa A, Sabater S (2019) Effects of multiple stressors on river biofilms depend on the time scale. Sci Rep 9(1):15810. https://doi.org/10.1038/s41598-019-52320-4
OECD SIDS (1999). 4-Hydroxybenzoic acid. http://www.inchem.org/documents/sids/sids/99967.pdf. Accessed 26 Jun 2023
Simčič T, Mori N (2007) Intensity of mineralization in the hyporheic zone of the prealpine river Bača (West Slovenia). Hydrobiologia 586:221–234. https://doi.org/10.1007/s10750-007-0621-x
Simčič T, Pengal P, Eleršek T, Cokan B, Mori N (2022) Ecological processes in the hydropower plant reservoir under multiple anthropogenic pressures. Fundam Appl Limnol 195:257–273. https://doi.org/10.1127/fal/2021/1408
Thomas FA (2021) Mohan M Krishnan KP (2021) Bacterial diversity and their metabolic profiles in the sedimentary environments of Ny-Ålesund. Arctic Antonie Van Leeuwenhoek 114:1339–1360. https://doi.org/10.1007/s10482-021-01604-9
Trevors J (1984) The measurement of electron transport system (ETS) activity in freshwater sediment. Water Res 18:581–584. https://doi.org/10.1016/0043-1354(84)90207-0
IPCC (2021) Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Masson-Delmotte, V., P. Zhai, A. Pirani, S.L. Connors, C. Péan, S. Berger, N. Caud, Y. Chen, L. Goldfarb, M.I. Gomis, M. Huang, K. Leitzell, E. Lonnoy, J.B.R. Matthews, T.K. Maycock, T. Waterfield, O. Yelekçi, R. Yu, and B. Zhou (eds.)]. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, 2391 pp. https://doi.org/10.1017/9781009157896
Volk CJ, Volk CB, Kaplan LA (1997) Chemical composition of biodegradable dissolved organic matter in streamwater. Limnol Oceanogr 42:39–44. https://doi.org/10.4319/lo.1997.42.1.0039
Wagenhoff A, Townsend CR, Phillips N, Matthaei CD (2011) Subsidy-stress and multiple-stressor effects along gradients of deposited fine sediment and dissolved nutrients in a regional set of streams and rivers. Freshw Biol 56:1916–1936. https://doi.org/10.1111/j.1365-2427.2011.02619.x
Wagner K, Bengtsson MM, Besemer K, Sieczko A, Burns NR, Herberg ER, Battin TJ (2014) Functional and structural responses of hyporheic biofilms to varying sources of dissolved organic matter. Appl Environ Microbiol 80:6004–6012. https://doi.org/10.1128/AEM.01128-14
Weber KP, Legge RL (2009) One-dimensional metric for tracking bacterial community divergence using sole carbon source utilization patterns. J Microbiol Methods 79:55–61. https://doi.org/10.1016/j.mimet.2009.07.020
Weber KP, Legge RL (2010) Community-Level Physiological Profiling. In: Cummings SP (ed) Bioremediation: Methods and Protocols. Humana Press, Totowa, NJ, pp 263–281
Weigelhofer G, Jirón TS, Yeh TC, Steniczka G, Pucher M (2020) Dissolved organic matter quality and biofilm composition affect microbial organic matter uptake in stream flumes. Water 12:3246. https://doi.org/10.3390/w12113246
Woodward G, Perkins DM, Brown LE (2010) Climate change and freshwater ecosystems: impacts across multiple levels of organization. Philos Trans R Soc Lond B Biol Sci 365:2093–2106. https://doi.org/10.1098/rstb.2010.0055
Ylla I, Romaní AM, Sabater S (2012) Labile and recalcitrant organic matter utilization by river biofilm under increasing water temperature. Microb Ecol 64:593–604. https://doi.org/10.1007/s00248-012-0062-6
Ylla I, Canhoto C, Romaní AM (2014) Effects of warming on stream biofilm organic matter use capabilities. Microb Ecol 68:132–145. https://doi.org/10.1007/s00248-014-0406-5
Zhang L, Delgado-Baquerizo M, Shi Y et al (2021) Co-existing water and sediment bacteria are driven by contrasting environmental factors across glacier-fed aquatic systems. Water Res 198:117139. https://doi.org/10.1016/j.watres.2021.117139. (PMID: 33895591)
Acknowledgements
This study was financially supported by the Slovenian Research and Innovation Agency (Research Program P1-0255). We thank Tjaša Matjašič for help during laboratory work and anonymous reviewers for the constructive comments and suggestions.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Responsible editor: Thomas Reid
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Simčič, T., Poklukar, M. & Mori, N. Response of hyporheic biofilms to temperature changes and dissolved organic carbon enrichment: a mesocosm study. J Soils Sediments (2024). https://doi.org/10.1007/s11368-024-03845-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11368-024-03845-3