
Abstract
Key message
Destruction of PAP phosphatase AHL suppresses atrzf1 phenotype in abiotic stress responses. AHL plays an intermediate role in the regulation of proline accumulation by PAP nucleotidase.
Abstract
Proline (Pro) metabolism is important for environmental responses, plant development, and growth. However, the role of Pro in abiotic stress process is unclear. Using atrzf1 (Arabidopsis thaliana ring zinc finger 1) mutant as a parental line for T-DNA tagging mutagenesis, we identified a suppressor mutant designated as proline content alterative 17 (pca17) that suppressed insensitivity of atrzf1 to abiotic stresses during early seedling growth. Pro content of pca17 was lower than that in both wild type (WT) and atrzf1 while complementary lines were less sensitive to abscisic acid (ABA) and abiotic stresses compared to WT. Thermal Asymmetric Interlaced (TAIL)-PCR of pca17 showed that T-DNA was inserted at site of At5g54390 (AHL for Arabidopsis Halotolerance 2-like) encoding 3′-phosphoadenosine-5′-phosphate (PAP) phosphatase. Under drought stress condition, products of sulfate metabolism such as PAP and adenosine monophosphate were significantly lower in pca17 than those in WT and atrzf1. Furthermore, pca17 showed significantly higher levels of several important drought parameters including malondialdehyde, ion leakage, and water loss than WT and atrzf1. Fluorescence signal of green fluorescent protein (GFP)-tagged AHL was quite strong in nuclei of the root and guard cells of transgenic seedlings. Additionally, AHL promoter-β-glucuronidase (GUS) construct revealed substantial gene expression in vasculature tissues and pollen. Collectively, these findings demonstrate that pca17 acts as a dominant suppressor mutant of atrzf1 in abiotic stress response by modulating proline and sulfate metabolism.








Similar content being viewed by others

Abbreviations
- ABA:
-
Abscisic acid
- GFP:
-
Green fluorescent protein
- PAP:
-
3′-Phosphoadenosine-5′-phosphate
- qPCR:
-
Quantitative real-time polymerase chain reaction
- RT-PCR:
-
Reverse transcription-PCR
- WT:
-
Wild-type
References
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39:205–207
Bechtold N, Pelletier G (1998) In planta Agrobacterium-mediated transformation of adult Arabidopsis thaliana plants by vacuum infiltration. Methods Mol Biol 82:259–266
Borsani O, Zhu J, Verslues PE, Sunkar R, Zhu JK (2005) Endogenous siRNAs derived from a pair of natural cis-antisense transcripts regulate salt tolerance in Arabidopsis. Cell 123:1279–1291
Chan KX, Wirtz M, Phua SY, Estavillo GM, Pogson BJ (2013) Balancing metabolites in drought: the sulfur assimilation conundrum. Trends Plant Sci 18:18–29
Chan KX, Mabbitt PD, Phua SY, Mueller JW, Nisar N, Gigolashvili T et al (2016) Sensing and signaling of oxidative stress in chloroplasts by inactivation of the SAL1 phosphoadenosine phosphatase. Proc Natl Acad Sci USA 113:E4567–E4576
Chen H, Zhang B, Hicks LM, Xiong L (2011) A nucleotide metabolite controls stress-responsive gene expression and plant development. PLoS ONE 6:e26661
Deuschle K, Funck D, Hellmann H, Däschner K, Binder S, Frommer WB (2001) A nuclear gene encoding mitochondrial Delta-pyrroline-5-carboxylate dehydrogenase and its potential role in protection from proline toxicity. Plant J 27:345–355
Dichtl B, Stevens A, Tollervey D (1997) Lithium toxicity in yeast is due to the inhibition of RNA processing enzymes. EMBO J 16:7184–7195
Earley KW, Haag JR, Pontes O, Opper K, Juehne T, Song K et al (2006) Gateway-compatible vectors for plant functional genomics and proteomics. Plant J 45:616–629
Estavillo GM, Crisp PA, Pornsiriwong W, Wirtz M, Collinge D, Carrie C et al (2011) Evidence for a SAL1-PAP chloroplast retrograde pathway that functions in drought and high light signaling in Arabidopsis. Plant Cell 23:3992–4012
Gigolashvili T, Geier M, Ashykhmina N, Frerigmann H, Wulfert S, Krueger S et al (2012) The Arabidopsis thylakoid ADP/ATP carrier TAAC has an additional role in supplying plastidic phosphoadenosine 5′-phosphosulfate to the cytosol. Plant Cell 24:4187–4204
Gil-Mascarell R, López-Coronado JM, Bellés JM, Serrano R, Rodríguez PL (1999) The Arabidopsis HAL2-like gene family includes a novel sodium-sensitive phosphatase. Plant J 17:373–383
Gy I, Gasciolli V, Lauressergues D, Morel JB, Gombert J, Proux F et al (2007) Arabidopsis FIERY1, XRN2, and XRN3 are endogenous RNA silencing suppressors. Plant Cell 19:3451–3461
Halkier BA, Gershenzon J (2006) Biology and biochemistry of glucosinolates. Annu Rev Plant Biol 57:303–333
Hare PD, Cress WA, Staden JV (1999) Proline synthesis and degradation: a model system for elucidating stress-related signal transduction. J Exp Bot 50:413–434
Hayat S, Hayat Q, Alyemeni MN, Wani AS, Pichtel J, Ahmad A (2012) Role of proline under changing environments: a review. Plant Signal Behav 7:1456–1466
Jefferson RA, Klass M, Wolf N, Hirsh D (1987) Expression of chimeric genes in Caenorhabditis elegans. J Mol Biol 1:41–46
Ju HW, Min JH, Chung MS, Kim CS (2013) The atrzf1 mutation of the novel RING-type E3 ubiquitin ligase increases proline contents and enhances drought tolerance in Arabidopsis. Plant Sci 203–204:1–7
Karimi M, Inze D, Depicker A (2002) GATEWAYTM vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci 7:193–195
Kim AR, Min JH, Lee KH, Kim CS (2017) PCA22 acts as a suppressor of atrzf1 to mediate proline accumulation in response to abiotic stress in Arabidopsis. J Exp Bot 68:1797–1809
Klein M, Papenbrock J (2004) The multi-protein family of Arabidopsis sulphotransferases and their relatives in other plant species. J Exp Bot 55:1809–1820
Kopriva S (2006) Regulation of sulfate assimilation in Arabidopsis and beyond. Ann Bot 97:479–495
Kopriva S, Mugford SG, Matthewman C, Koprivova A (2009) Plant sulfate assimilation genes: redundancy versus specialization. Plant Cell Rep 28:1769–1780
Kopriva S, Mugford SG, Baraniecka P, Lee BR, Matthewman CA, Koprivova A (2012) Control of sulfur partitioning between primary and secondary metabolism in Arabidopsis. Front Plant Sci 3:163
Lee JH, Kim WT (2011) Regulation of abiotic stress signal transduction by E3 ubiquitin ligases in Arabidopsis. Mol Cells 31:201–208
Li X, Zhang H, Tian L, Huang L, Liu S, Li D et al (2015) Tomato SIRbohB, a member of the NADPH oxidase family, is required for disease resistance against Botrytis cinerea and tolerance to drought stress. Front Plant Sci 6:463
Liang X, Zhang L, Natarajan SK, Becker DF (2013) Proline mechanisms of stress survival. Antioxid Redox Signal 19:998–1011
Liu YG, Mitsukawa N, Oosumi T, Whittier RF (1995) Efficient isolation and mapping of Arabidopsis thaliana T-DNA insert junctions by thermal asymmetric interlaced PCR. Plant J 8:457–463
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25:402–408
Mugford SG, Yoshimoto N, Reichelt M, Wirtz M, Hill L, Mugford ST et al (2009) Disruption of adenosine-5′-phosphosulfate kinase in Arabidopsis reduces levels of sulfated secondary metabolites. Plant Cell 21:910–927
Pornsiriwong W, Estavillo GM, Chan KX, Tee EE, Ganguly D, Crisp PA et al (2017) A chloroplast retrograde signal, 3′-phosphoadenosine 5′-phosphate, acts as a secondary messenger in abscisic acid signaling in stomatal closure and germination. eLife 6:e23361
Qin F, Sakuma Y, Tran LSP, Maruyama K, Kidokoro S, Fujita Y et al (2008) Arabidopsis DREB2A-interacting proteins function as RING E3 ligase and negatively regulate plant drought stress-responsive gene expression. Plant Cell 20:1693–1707
Rajendrakumar CS, Suryanarayana T, Reddy AR (1997) DNA helix destabilization by proline and betaine: possible role in the salinity tolerance process. FEBS Lett 410:201–205
Ristic Z, Ashworth EN (1993) Changes in leaf ultrastructure and carbohydrates in Arabidopsis thaliana L. (Heyn) cv. Columbia during rapid cold acclimation. Protoplasma 172:111–123
Shinozaki K, Yamaguchi-Shinozaki K (2000) Molecular responses to dehydration and low temperature: differences and cross-talk between two stress signaling pathways. Plant Biol 3:217–223
Singh M, Kumar J, Singh VP, Prasad SM (2014) Proline and salinity tolerance in plants. Biochem Pharmacol 3:e170
Smart RE, Bingham GE (1974) Rapid estimates of relative water content. Plant Physiol 53:258–260
Szabados L, Savouré A (2010) Proline: a multifunctional amino acid. Trends Plant Sci 15:89–97
Takahashi S, Seki M, Ishida J, Satou M, Sakurai T, Narusaka M et al (2004) Monitoring the expression profiles of genes induced by hyperosmotic, high salinity, and oxidative stress and abscisic acid treatment in Arabidopsis cell culture using a full-length cDNA microarray. Plant Mol Biol 56:29–55
Verbruggen N, Hermans C (2008) Proline accumulation in plants: a review. Amino Acids 35:753–759
Weigel D, Ahn JH, Blázquez MA, Borevitz JO, Christensen SK, Fankhauser C et al (2000) Activation tagging in Arabidopsis. Plant Physiol 122:1003–1013
Xiong L, Lee H, Huang R, Zhu JK (2004) A single amino acid substitution in the Arabidopsis FIERY1/HOS2 protein confers cold signaling specificity and lithium tolerance. Plant J 40:536–545
Yang SL, Lan SS, Gong M (2009) Hydrogen peroxide-induced proline and metabolic pathway of its accumulation in maize seedlings. Plant Physiol 166:1694–1699
Zhang YY, Li Y, Gao T, Zhu H, Wang DJ, Zhang HW et al (2008) Arabidopsis SDIR1 enhances drought tolerance in crop plants. Biosci Biotechnol Biochem 72:2251–2254
Zhang X, Ivanova A, Vandepoele K, Radomiljac J, Van de Velde J, Berkowitz O et al (2017) The transcription factor MYB29 is a regulator of Alternative Oxidase 1. Plant Physiol 173:1824–1843
Acknowledgements
This work was supported in part by a grant to C.S.K. from the Next-Generation BioGreen21 program (SSAC, PJ013171) funded by the Rural Development Administration and by the Basic Science Research Program funded by the Ministry of Education, Science and Technology of Korea (2018R1D1A1B07045242).
Author information
Authors and Affiliations
Contributions
We thank Dr. Y-MK for technical assistance with HPLC analysis. CSK designed experiments and interpreted results. DJS, JHM, and TVN carried out experiments and interpreted the results.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest relevant to this study to disclose.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Table S1
Primers used for TAIL-PCR, qPCR, RT-PCR, and gene cloning (DOCX 18 KB)
Supplementary Fig. S1
Germination of pca17 mutant following abiotic stress. a Seeds of samples (WT, atrzf1, and pca17) were sown on MS medium and permitted to grow for 8 days. Photograph indicates that seedling growth and green cotyledons among WT, atrzf1, and pca17 plants were similar. b-d Seeds were sown on MS agar plates supplemented with 400 mM mannitol (b), 1 μM ABA (c), or 6 mM H2O2 (d). Complete emergence of the radicle indicates germination. It was scored on the indicated days. Data represent mean ± standard deviation values of three independent experiments (n = 50 of each). Error bars represent standard deviations. Different letters above bars indicate statistically significant difference (ANOVA, P < 0.05) (TIF 908 KB)
Supplementary Fig. S2
Cotyledon greening of pca17 mutant following abiotic stress and ABA treatment. Seeds were sown on MS agar plates supplemented with 400 mM mannitol (a), 1 μM ABA (b), or 6 mM H2O2 (c) and cultured for 6-8 days. Photographs were taken on day 6 for dehydration and ABA or on day 8 for oxidative stress. Decreased development and fewer green cotyledons were observed in pca17 compared with those in WT and atrzf1 under stress conditions (TIF 3074 KB)
Supplementary Fig. S3
Reduced tolerance of pca17 mutants to drought stress. a Drought sensitivity of pca17 mutants. Two-week-old WT, atrzf1, and pca17 plants were subjected to drought stress by withholding water for 10 days and then re-watering for 3 days. b Survival rates of WT, atrzf1 and pca17 plants were measured after re-watering. Error bars represent standard deviations. Different letters above bars indicate statistically significant difference (ANOVA, P < 0.05) (TIF 2683 KB)
Supplementary Fig. S4
Alignment of full-length deduced amino acid sequences of AHL and structural features of the AHL protein. a Sequences of Arabidopsis AHL (At5g54390), SAL1 (At5g63980), SAL2 (At5g64000), SAL3 (At5g63990), and SAL4 (At5g09290). Black and grey shading indicate identical and similar amino acids, respectively. Dashes indicate gaps introduced to optimize alignment. b Structure of the conserved motif region in AHL. Light gray, gray, and black boxes indicate active domain (41-310 aa), lithium-binding domain (144-310 aa), and substrate-binding domain (SBD) (257-310 aa), respectively (http://myhits.isb-sib.ch). c Phylogenetic tree depicting homology relationships among Arabidopsis thaliana AHL members (http://www.megasoftware.net). Branch length represents substitutions per site (TIF 1593 KB)
Supplementary Fig. S5
Abiotic stress response in complementation transgenic plants. aAHL expression levels in WT, atrzf1, pca17, three independent complementation transgenic lines (Com4, Com6, and Com9), three independent ahl RNAi (ri2-3, ri5-2, and ri6-7), and three independent AHL-overexpressing lines (OE2-2, OE5-1, and OE7-7) were confirmed by qPCR using RNA extracted from 8-day-old seedlings. Actin 1 (ACT1) was used as an internal qPCR control. Error bars indicate standard deviations from three independent experiments (n = 7 of each). Different letters above bars indicate statistically significant difference (ANOVA, P < 0.05). b-e Seeds were sown on MS agar medium without additive (b) or with 400 mM mannitol (c), 1 µM ABA (d), or 6 mM H2O2 (e) and grown for 8 days after germination. Photographs show that atrzf1, three independent complementation, and three independent AHL-overexpressing lines exhibit better development and green cotyledons than WT, pca17 and three independent ahl RNAi plants under dehydration, ABA, and oxidative stress conditions (TIF 2065 KB)
Supplementary Fig. S6
AHL expression in Arabidopsis plants under abiotic stress conditions. qPCR analyses showing expression levels of AHL, RAB18, and APX1 in response to abiotic stress. All quantifications were made in three independent RNA samples obtained from plants treated with 100 µM ABA (a), 400 mM mannitol (b), or 6 mM H2O2 (c) for the indicated times. Error bars indicate standard deviations from three independent experiments conducted. Each experiment was performed with total RNA obtained from ten pooled seedlings of each line (WT, atrzf1 and pca17). Arabidopsis Actin 1 was used as an internal control. Different letters above bars indicate statistically significant difference (ANOVA, P < 0.05) (TIF 302 KB)
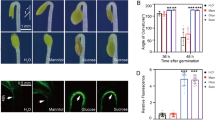
Supplementary Fig. S7
Nuclear localization of AHL in transgenic Arabidopsis plants. a Five-day-old transgenic plants grown on MS agar medium were analyzed for GFP expression by confocal microscopy. AHL-GFP green fluorescent signal was observed in the nuclei of root cells. b Ten-day-old transgenic plants grown on MS agar medium were analyzed for GFP expression in guard cells by confocal microscopy. AHL-GFP green fluorescent signal was relatively strong in the nuclei of guard cells. GFP, green fluorescent protein; B/W, black and white. Scale bars = 200 µm. This experiment was performed in duplicate. Five individual seedlings were analyzed per experiment and similar results were obtained. c Immunoblot analysis of transgenic plants producing AHL-GFP fusions. Plant extracts were prepared from 10-day-old seedlings, 20 µL of cell lysate was loaded in each lane, and AHL-GFP (arrow) was detected with an anti-GFP monoclonal antibody. WT, wild-type plant; AHL-GFP, Arabidopsis expressing AHL-GFP. Molecular masses (kDa) of protein standards are shown on the left. Total proteins were separated by SDS-PAGE and electrophoretically transferred onto nylon membranes followed by Ponceau S staining (TIF 1595 KB)
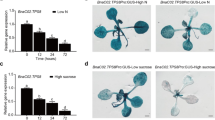
Supplementary Fig. S8
AHL promoter-GUS expression pattern in transgenic Arabidopsis plants. a GUS staining for a 7-day-old seedling plant. b GUS staining results are shown in a flower of a 4-week-old transgenic plant. GUS expression was observed in the vascular system of leaves or hypocotyl, the anther, the filament, and the vein of the petal. c-f Ten-day-old seedlings of AHL promoter-GUS transgenic plants were subjected to H2O (c), 400 mM mannitol (d), 50 µM ABA (e), or 6 mM H2O2 (f). After 12 h of abiotic stress exposure, seedlings were stained for GUS expression. H2O-treated seedlings were used as the control (c). This experiment was repeated twice. Each GUS staining assay was analyzed from five individual seedlings per experiment and similar results were obtained (TIF 1116 KB)
Supplementary Fig. S9
Proline contents in WT, atrzf1, pca17 and AHL transgenic plants under drought stress. Light-grown 5-week-old plants were grown for 7 days without watering. Proline contents were measured from drought-treated leaves. Values of Pro content represent the average of three independent experiments. Each assay was performed with extracts obtained from ten seedlings of each plant. Error bars represent standard deviations. Different letters above bars indicate statistically significant difference (ANOVA, P < 0.05) (TIF 122 KB)
Rights and permissions
About this article

Cite this article
Shin, DJ., Min, JH., Van Nguyen, T. et al. Loss of Arabidopsis Halotolerance 2-like (AHL), a 3′-phosphoadenosine-5′-phosphate phosphatase, suppresses insensitive response of Arabidopsis thaliana ring zinc finger 1 (atrzf1) mutant to abiotic stress. Plant Mol Biol 99, 363–377 (2019). https://doi.org/10.1007/s11103-019-00822-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11103-019-00822-0