
Abstract
Bumble bees (Bombus; Hymenoptera) are crucial pollinators for many agricultural crops, but their numbers have declined - with climate change as a possible driver. Different aspects of local weather conditions can have substantial effects on bumble bees’ ability to forage. Here we made 501 observations of bumblebee workers on bramble flowers (Rubus fruticosus) to explore how the characteristic sizes of foragers active and their individual rate of movement between flowers were correlated with aspects of weather. Firstly, we found that increased ambient temperature is correlated to reduced activity in larger workers. Moreover, while higher humidity was linked to an increase in smaller workers, an increase in cloud cover led their numbers to decrease. Visitation rate and temperature showed a significant negative relationship whereas no other weather aspect affected visitation rate significantly. Our results suggest that predicted rises in ambient temperatures will adversely affect bumble bee foraging, and particularly so for larger-bodied workers. We anticipate that this study can be used as a foundation for longer-term and more detailed studies on bumble bee foraging and colony performance. Such work is vital to identify management measures to mitigate pollinator declines and preserve food security.
Similar content being viewed by others


Introduction
Pollination is one of the key services that maintain ecosystems (Klein et al. 2007) and is crucial for the sexual reproduction of about 70% of tropical and 84% of European crops (Williams 1994; Roubik 1995). Thus, human food security is largely dependent on the diversity and efficiency of pollinating animals (of which most are flying insects). While the global land-cover used to grow pollination-dependent crops has increased (Aizen et al. 2009), the number of flying insects has substantially decreased in the last thirty years (Potts et al. 2010; Dirzo et al. 2014; Hallmann et al. 2017). Pollinators are threatened by a range of human-induced changes such as the increased use of pesticides, a reduction of hedgerows and fragmentation of habitat (Potts et al. 2010; Feltham et al. 2015; Goulson et al. 2015). Moreover, climate change is also thought to be a major cause of pollinator loss (Warren et al. 2001; Memmott et al. 2007); and so investigating the effects of weather on pollinators is vital.
Climate change is causing shifts in local weather conditions all over the world (Howe 2018). Average ambient temperature is predicted to rise by 1.5-2 °C by the end of the century (Collins et al. 2013), while wind speeds are expected to increase by 1 up to 5 m s− 1 within the next one hundred years (Robinson et al. 2017). Short-term weather effects are also becoming more extreme; with increasing frequencies of drought in summers while winters have had a rise in recorded rainfall (Jenkins et al. 2008). As mentioned, these changes can have detrimental effects on pollinating insects as, for example, high wind speeds can adversely affect their ability to fly and forage efficiently (Combes and Dudley 2009; Chang et al. 2016). On the other hand, higher temperatures can limit the ability of some large or well-insulated pollinators to forage, due to their limited ability to lose heat effectively (May 1979; Heinrich and Esch 1994; De Marco et al. Jr 2005). This may also have caused the observed range changes of pollinators towards higher latitudes (Chen et al. 2011).
One pollinator group that is vital for pollinating a large number of crops are bumble bees (Velthuis and Van Doorn 2006). Bumble bees are eusocial insects that make up the genus Bombus, which comprises ~ 255 species in the family of Hymenoptera (Goulson 2010) and are primarily found in the northern hemisphere (Williams et al. 2009). They are characteristically larger and hairier than other species of Hymenoptera and the resulting insulation due to the lower surface area to volume ratio decreases heat loss (Heinrich 2004). Furthermore, they are facultatively endothermic and can raise their internal body temperature to over 30 °C by contracting their flight muscles simultaneously to generate heat (Heinrich 1993) and by substrate cycling catalyzed by the enzyme fructose bisphosphatase (Newsholme et al. 1972; Clark et al. 1973). These adaptations allow them to forage in lower ambient temperatures than other pollinating insects (Ahn et al. 1988) and enables a successful provision of nectar and pollen to the colony even during colder spring months.
However, even with their ability to warm up their flight muscles, most bumble bee workers are restricted to foraging at ambient temperatures of about 12 °C (Goulson 2010). On the other hand, recent rises in average temperature especially during the summer (Jenkins et al. 2008) have increased their risk of overheating due to their larger, well-insulated bodies. A thoracic temperature of about 42 °C is lethal (Heinrich and Heinrich 1983) and despite having some methods of releasing heat (counter-current heat exchange through the petiole: Heinrich (2004); black fur on thorax: Williams (2007), the foraging activity of many workers may be limited in high temperatures as well. Moreover, as well as air temperature, they are affected by a range of other aspects of local weather. Crall et al. (2017) showed that wind and turbulence can significantly affect bumble bee body stability and wing kinematics which, in turn, affect their ability to land on flowers precisely. High wind speeds, especially when coming from the side, also cause the need for constant adjustment to prevent yaw and roll, and workers are more likely to have high-impact landings on flowers under these conditions (Chang et al. 2016). The effects of humidity are less well resolved, as studies indicate contradicting results (Sanderson et al. 2015; Yankit et al. 2020). Evidence does suggest, however, that air humidity affects which resource is collected by bumble bees and lower humidity seems to be preferable for pollen collection (Peat and Goulson 2005). Further, bumble bees are restricted by their low visual acuity, which decreases their ability to see in low light conditions (Kapustjanskij et al. 2007). Although some workers were observed to leave the nest during twilight (less than 10 lx), it was shown that their ability to successfully search for flowers is adversely affected by low light intensities (Hall et al. 2021).
The extent to which bumble bee workers are affected by local weather conditions is also partly influenced by their individual size (Heinrich and Heinrich 1983; Heinrich 2004; Kapustjanskij et al. 2007; Goulson 2010; Willmer and Finlayson 2014). Bumble bees divide the work in the colony at least to some extent based on alloethism, meaning that tasks are divided based on worker size; rather than age as seen in, for example, honeybees (Wilson 1971). Individual body mass can vary up to ten-fold within a single nest (Alford 1975) where smaller workers conduct nest-based tasks whereas larger workers are primarily foragers (Goulson 2010). However, even within foragers there is large variation in sizes and some studies have recorded a more than two-fold difference in mass between the smallest and largest forager active at one time in the same colony (Goulson et al. 2002; Spaethe and Weidenmüller 2002). Some studies suggest that larger workers are better foragers, as they can carry more nectar and have a higher foraging efficiency in comparison to smaller ones (Spaethe and Weidenmüller 2002; Willmer and Finlayson 2014), and are better equipped to tolerate colder conditions due to their lower surface area to volume ratio (Heinrich 2004). Contrary to this, Peat and Goulson (2005) found no significant effect of increasingly poorer weather conditions on worker size. Nonetheless, Hall et al. (2021) and Kapustjanskij et al. (2007) found that larger bees were more successful foragers than smaller bees in lower light conditions due to their larger eye size (Taylor et al. 2019). Additionally, Crall et al. (2015) observed that larger workers fly faster in windy conditions but have greater need to perform corrective maneuvers if obstacles are in their flight path. Thus, further research is required to solve the discrepancies in previous studies and directly investigate the effects of weather conditions on the same population of bumble bees.
The aim of this study is to investigate the effects of weather conditions on the size distribution of actively foraging workers and their visitation rate in the field. Most studies investigating the effects of weather conditions on worker size did so by monitoring and measuring bees as they left and arrived at the nest (Goulson et al. 2002; Spaethe and Weidenmüller 2002; Peat and Goulson 2005; Hall et al. 2021). Although this allows an accurate measuring of time spent outside the nest and bee weight by using integrated scales in nest entrances and ID-tags, one cannot say for sure whether workers leaving the nest do so to forage, or for example for defecation or orientation flights (Cisarovsky 2013; Philippides et al. 2013). Hence, we aim to research bumble bee foraging behavior on flowers in the field to focus solely on foraging workers. We did this by recording weather conditions regularly throughout the day while observing bumble bee foragers on bramble flowers (Rubus fruticosus agg.) and documenting their species, size, and flower visits per minute. Based on previous work, we predict that larger workers would be more common at colder temperatures, lower light conditions and higher wind speeds. Moreover, we expect that the rate at which different flowers are visited would be negatively correlated with temperature as well as higher wind speeds.
Methods and Materials
Worker Size
During a one-week period from the 23 to the 30 May 2021, 102 bumble bees of a range of different species were caught opportunistically on flowers and their thorax width at its widest point measured to the nearest 0.1 mm (Goulson et al. 2002; Peat and Goulson 2005) using electronic Vernier calipers (RoHS, England). By determining peaks in a size-frequency histogram (Fig. 1) and using visual classification (Willmer and Finlayson 2014), the bees were categorized (using an ad hoc classification) as either small (≤ 5.5 mm thorax width), medium (> 5.5 to 7.0 mm), large (> 7.0 to 9.0 mm) and extra-large (> 9.0 mm). All but extra-large individuals were considered workers with extra-large bees being counted as gynes. Measured bees were larger on average than found in previous UK studies (Goulson et al. 2002; Del Castillo and Fairbairn 2012), but it has been suggested that bumble bees are larger in Scotland as it is colder than England (Peat et al. 2005), which was also supported by the findings of Willmer and Finlayson (2014). Another 100 bees were then caught, and size was classified based on visual inspection before measuring the individual, to determine accuracy of visual categorization of the observer. Over 90% of these bees had a thorax width within the range expected according to the visual classification On this basis, we assume that bees during the main observations were categorized reasonably accurately on the basis of visual inspection. This avoided the need to catch them during the main observation period which would have influenced their subsequent foraging behavior. Another 50 bees were caught after three weeks of data collection to ensure that accuracy remained high, and again more than 90% of individual had a thorax width within the range expected according to the visual classification.

Histogram showing the size distribution of a total of 102 bumble bees that were caught and the thorax width was measured at its widest point using Vernier calipers. Size categories were chosen based on peaks in the histogram as well as visual cues. Thus, workers were categorized into small (≤ 5.5 mm), medium (5.6 to 7.0 mm), large (7.1 to 9.0 mm) and extra-large (> 9.0 mm) bees were considered gynes and, hence, not considered during the main data collection period
Weather and Visitation Rate
Data collection occurred between the 10 June and 04 July 2021 around St Andrews, Scotland. Observations were made between 10am to 5pm, the peak foraging time for pollinators (Garbuzov et al. 2017), in all conditions except heavy rain that prevented any foraging activity. However, if no bumble bees were seen for 30 min the conditions were considered unsuitable and data collection ended for that day. Bumble bees were observed on bramble (Rubus fruticosus), a common plant locally that flowers between June and July and is frequently visited by bumble bee species (Goulson et al. 2005; Wignall et al. 2020). We surveyed at two sites (Site 1: Botanical Garden St Andrews, 56°20’09.8"N 2°48’17.0"W; Site 2: East Sands Coastal Path, St Andrews, 56°19’53.3"N 2°46’11.8"W) which were assigned a number based on the flowering start date of the present bramble. Bramble at the Botanical Garden bloomed first and was, thus, site 1. Data was collected there until another site was in peak bloom. As sites were added, they were surveyed on alternating days (Day 1: Site 1, Day 2: Site 2, etc.).
Temperature, humidity, and wind speed were measured with hand-held instruments every 30 min; and light intensity every 15 min due to higher variability because of cloud cover changes. Wind speed and light intensity measurements occurred at 1 m above ground but the thermohydrometer was placed on the ground to ensure shade coverage and prevent overestimates due to the effects of direct sunlight on the thermometer. The thermohydrometer (Vaisala HM40, SW version 1.7, Finland) was left for about 10–15 min before recording the first data to give it time to adjust to conditions. Wind speed (Anemometer, RS AM-4201, United Kingdom) was measured by recording it for 60 s and the highest recorded value was documented as the current wind speed. Light intensity (ATP ETI-1301, United Kingdom) was measured while facing the sun but holding the device horizontal. Cloud cover was categorized in okta (number of eighth of the sky covered in clouds) and was assessed immediately after each foraging observation.
When a bumble bee was seen foraging on a flower, we started a stopwatch (Garmin, vivoactive 4s) and recorded the number of flowers visited during the foraging event. The maximum time of observation for each individual bee was 5 min. For each worker, we recorded date, time of start of observation, bumble bee species, size category, total number of flowers visited, duration of observation, presence or absence of pollen in pollen baskets, and the relevant weather data. Bee species were identified using Pinchen (2006), and Bombus terrestris and the morphologically very similar Bombus lucorum were both recorded as B. terrestris/lucorum due to the high risk of misidentification in the field (Wolf et al. 2010). We recorded the total number of flower visitations without consideration of whether a recorded sequence of visits involved different plants on the same patch or return visits to the same flower. Visitationrate was then calculated per minute by dividing the number of flowers visited by the duration in seconds and multiplying the result by 60.
On some occasions the workers visited flowers that were hidden from view by leaves or within the bushes so that accurate flower visitation could not have been recorded. In these cases, the stopwatch was paused, and we waited about 30 s for the bee to reappear into our field of view. If it did, the stopwatch was resumed, and the observation continued. If not, this individual observation was considered finished. If bees foraged for longer than 5 min, we tried to ensure not to record the same bee twice. Given typical values for the size of bumble bee colonies, the turnover of workers within colonies, typical visitation rate, and density of colonies, we except that the bulk of our observations will involve different individuals.
Statistical Analysis
All data was analysed and using R v. 4.1.0 (R Core Team 2021) and significance was taken as p < 0.05. Plots were created using the ggplot2 package (Wickham 2016).
To investigate whether the size of foraging worker bees was affected by local weather conditions a multinomial logistic regression with size of workers as the response variable and all recorded weather conditions and location as predictor variables was used (Model 1). This model was run using the nnet package in R (Venables and Ripley 2002).
The model was run a total of three times, each time with a different worker size as the intercept. Worker size levels were changed using the relevel function in R and allowed for pairwise comparison between the different response variable factors. All predictor variables were kept in the model, even if they had no significant effect on worker size as they were all considered important explanatory variables. The same was done for all other models run (see below).
To investigate the effects of local weather conditions on species composition Kruskal Wallis tests followed by pairwise Mann-Whitney U tests for pairwise comparisons were used. These were chosen as data distribution for all weather conditions was non-parametric (Shapiro-Wilk, p < 0.05 for all conditions). As species distribution differed with weather conditions (see Results), the above explained multinomial logistic regression was repeated but included the three most common species (B. hypnorum, B. terrestris/lucorum, B. pascuorum) as explanatory variables to investigate whether the changes seen in size were due to weather conditions rather than the species make-up of the actively foraging bee population.
We examined whether there were significant differences in visitation rate between worker sizes and species by using a Kruskal Wallis test, as data for visitation rate was non-parametric (Shapiro-Wilk, W = 0.927, p < 0.001). Moreover, to model the effects of local weather condition and location on the visitation rate a GLM with an inverse Gaussian family and identity link function was fitted. Hence, just like in Model 1 all weather conditions and location were included as factors, but also presence of pollen in pollen baskets and the number of open flowers were explanatory variables in the model to predict visitation rate (Model 2). Data for visitation rate was not normally distributed (Shapiro-Wilk, W = 0.927, p < 0.001) but right-skewed and log-transforming the data did not make it parametric (Shapiro-Wilk, W = 0.994, p = 0.038). Thus, the inverse Gaussian family was chosen as it deals with strictly positive, right-skewed data (Dunn and Smyth 2018) and the assumptions of the model for variance of residuals and a linear relationship between the response and the linear predictor were met.
Results
Effects of Weather on Worker Size
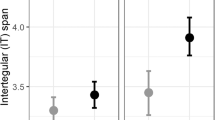
There are a total of 501 observations from 17 days of data collection, 394 of which were made at the Botanical Garden and 107 at the Coastal Path near East Sands. Medium workers were the most common (254 individuals), followed by large (176) and lastly small (71) individuals. Bombus hypnorum and Bombus terrestris/lucorum were the most common species, with Bombus lapidarius being the least common as it was only recorded 7 times. Bombus terrestris/lucorum and soroeensis showed the greatest proportion of large workers, whereas pratorum and lapidarius have the largest proportion of small workers (Fig. 2).

(A) Number of observations of each bumble bee species and respective size distribution within the species. The most commonly recorded species was Bombus hypnorum (n = 193), followed by B. terrestris/lucorum (n = 122), B. pascuorum (n = 75), B. pratorum (n = 54), B. soroeensis (n = 36), B. hortotum (n = 14), and lastly B. lapidarius (n = 7). (B) Percentage distribution of size categories within each species. The species with the highest proportion of large workers were B. terrestris/lucorum (67.2%), B. soroeensis (66.7%), and B. hortotum (50.0%) whereas B. soroeensis (5.5%) and B. terrestris/lucorum (5.8%) had the lowest percentage of small workers
Weather conditions varied substantially over the course of data collection. Air temperatures ranged from 12.8 to 27.5 °C with a median of 16.7 °C, with humidity fluctuating between 42.2% and 92.9% (median = 73.2%). The maximum wind speed record was 6.8 m s− 1 and the minimum was 0 m s− 1, with median wind speed being 1.3 m s− 1. Light intensity varied approximately 10-fold and ranged from 12,600 to 129,700 lx and a median of 52,900 lx. Cloud cover ranged between 1 and 8 oktas with a median of 6.
The effects of weather conditions on the presence of differently sized workers were modelled using a multinomial logistic regression (Table 1) and showed that temperature significantly affected the size of workers present at the bramble patches. With rising temperatures there were significantly more small workers active in comparison to both medium (Multinomial logistic regression, estimate ± SE: -0.081 ± 0.039, p = 0.03) and large worker bees (estimate ± SE: -0.169 ± 0.041p < 0.001). Moreover, there were also significantly fewer large than medium workers observed (estimate ± SE: -0.087 ± 0.029, p < 0.01) as temperature increased. Humidity also significantly influenced observed worker size and small workers were only substantially more common than large (estimate ± SE: -0.029 ± 0.012, p = 0.01), but not medium individuals (estimate ± SE: -0.016 ± 0.011, p = 0.15) as humidity rose. There was no significant difference between frequencies of observation of medium and large workers (estimate ± SE: -0.014 ± 0.008, p = 0.08) with changes of humidity. A similar pattern can be seen for cloud cover. Medium workers were not significantly more common than small ones during higher cloud scores (estimate ± SE: 0.089 ± 0.086, p = 0.297), but large worker bees were (estimate ± SE: 0.204 ± 0.095, p = 0.031). Although larger bees were more common than medium-sized individuals at higher cloud cover, this difference was not significant (estimate ± SE: 0.115 ± 0.070, p = 0.1). Medium sized workers were more common relative to small and large ones with increasing wind speed, but there was no significant difference between any bee sizes as a function of wind speed (p > 0.05 for all estimates). Moreover, light intensity and location did not significantly affect the size of workers foraging on bramble (p > 0.05 for all estimates, Table 1).
Relative species prevalence was also influenced by a variety of weather conditions. More specifically, after conducting pairwise comparisons using Mann-Whitney U tests after initial Kruskal Wallis tests, we found that workers of the characteristically medium bodied, brown-black-white colored species, Bombus hypnorum, differed most from others. They were relatively more common than half of the other observed species (Pairwise Mann-Whitney U, B. pratorum, B. terrestris/lucorum and B. soroeensis: p < 0.001) at higher temperatures. Furthermore, they were significantly more frequent at higher wind speeds in comparison to all species except the small to medium sized red-tailed species of B. lapidarius (for B. hortotum, B. pascuorum and B. terrestris/lucorum: p < 0.001; B. pratorum: p = 0.03; B. soroeensis: p = 0.04). On the other hand, they became significantly less common than four out of the six other species (B. lapidarius: p = 0.01; B. pascuorum, B. soroeensis and B. terrestris/lucorum: p < 0.001) as humidity rose; and less likely to forage in higher cloud cover than the two characteristically large-bodied white-tailed species B. terrestris/lucorum (p = 0.002) and B. soroeensis (p = 0.01). The usually medium sized red-tailed workers of B. pratorum became significantly rarer than B. terrestris/lucorum (p < 0.001) and B. soroeensis (p = 0.003) with increasing temperature. Moreover, workers of this species (p = 0.002), as well as worker of two other species (for B. hypnorum and B. pascuorum: p < 0.001), were also significantly less likely to forage in more humid conditions than the three-banded workers of B. hortotum. Light intensity was the only weather condition that did not affect the presence of any specific species during observations (Kruskal Wallis, H = 236.42, df = 218, p = 0.187).
As species distribution differed significantly with certain weather conditions, the multinomial logistic regression to predict worker size category was repeated but included only data from the three most common species. Species was then also added as a explanatory factor to the model, to investigate whether the differences were in fact due to changes in local weather conditions rather than bumble bee species. In comparison to B. hypnorum, B. terrestris/lucorum’s were significantly more likely to be large rather than small in size (estimate ± SE: 2.99 ± 0.42, p < 0.001), but there was not a significantly higher chance of being medium than small (estimate ± SE: 0.123 ± 0.39, p = 0.76). No other species was a significant predictor of size (p > 0.05 for all estimates) so it seems we can conclude that results from the original multinomial logistic regression were due to changes in weather conditions affecting bumble bee on the basis of size more than on the basis of other species-related traits.
Effects of Weather, Size, and Species on Visitation Rate
There was no significant difference in the visitation rates between different species (Kruskal Wallis, H = 346.9, df = 323, p = 0.173) and sizes (Kruskal Wallis, H = 325.9, df = 323, p = 0.44). However, the GLM used to investigate the effect of local weather conditions on visitation rates (Fig. 3) showed that higher temperatures were significantly correlated to a decrease in flower visits per minute (GLM, estimate ± SE: -0.149 ± 0.056, p = 0.008). During increased humidity there was a rise in visitation rate, but this was not significant (GLM, estimate ± SE: 0.012 ± 0.013, p = 0.5). Neither wind speed, light intensity or cloud cover significantly affected visitation rate (GLM, p > 0.05 for all coefficients, Table 2). However, flower visits per minute were significantly affected by location and were higher at site 2 (East Sands) than at site 1 (Botanical Garden, GLM, estimate ± SE: 3.619 ± 0.570, p < 0.001, Fig. 3). Additionally, the presence of pollen in pollen baskets resulted in a significantly lower visitation rate in workers (GLM, estimate ± SE: -0.610 ± 0.237, p = 0.01) whereas the number of open flowers was significantly positively correlated to visitation rate (GLM, estimate ± SE: 0.0006 ± 0.0002, p < 0.001).

The relationships between different weather conditions and location on the visitation rate of bumble bee workers on bramble. Data was collected over 17 days in June to July 2021 in St Andrews, Scotland from a total of 501 worker bees. Individual plots present the effect of (A) Temperature, (B) Humidity, (C) Wind speed, (D) Light intensity, (E) Number of open flowers, (F) Cloud cover, (G) Location, and (H) Presence of pollen on flower visits per minute. Lines in (A) to (E) present the lines of best fit using a linear regression with standard errors being represented by shaded areas
Discussion
Plant-pollinator interactions have co-evolved for a long time (Johnson and Anderson 2010) but these fine-tuned mutualistic relationships may now be threatened by rapid shifts in local weather conditions as a result of climate change (Memmott et al. 2007). Thus, it is crucial to investigate the effects of these changes on the ability of pollinators such as bumble bees to pollinate plants, reproduce and survive. In this study, by observing bumble bee workers on bramble flowers, we investigated whether changes in the recorded weather conditions affected worker size and visitation rate.
Worker Size
We found that the size of foraging workers decreases as temperature increases. This supports the hypothesis of Heinrich (2004) that larger workers have a lower surface area to volume ratio and are thus less able to lose heat efficiently, causing them to overheat in high temperatures. Interestingly, although our study showed a similar ambient temperature range as the one done by Peat and Goulson (2005), we did find a significant correlation between worker size and air temperature when they did not. Their use of commercial bumble bees allowed them to measure all workers to the nearest 0.1 mm, which provides more accurate size measurements than our three size categories. Regardless, our results indicate that larger workers may be crucial foragers during low temperature periods and also due to their increased capacity to carry pollen and nectar (Goulson et al. 2002). However, although they offer benefits, individually larger workers are more costly to rear (Goulson 2010) as they require more food during larval stages and a higher incubation temperature (Couvillon and Dornhaus 2009). This is specifically difficult to maintain when queens rear the first batch of workers as she regularly must leave the nest to forage, causing the eggs to cool down while she is gone (Goulson 2010). Hence, many colonies may not be able to rear high quantities of larger foragers at the start of the colony cycle when they are needed which can in turn lead to less overall workers in the summer and a decreased chance of successful queen rearing.
Climate change is used in some studies to refer to temperature changes only (Gérard et al. 2020), although a multitude of other aspects of weather are also affected (IPCC 2021). Thus, we investigated the effects of a range of other weather conditions and found, for example, no correlation between wind speed and forager size. This may be as body size is directly correlated to the size of other body parts such as wing length (Medler 1962) and, thus, all bumble bees, regardless of size, should show a similar body stability. Contrary to this, Crall et al. (2015) found that larger bumble bees have to adjust more when flying through obstacles in windy condition. However, although larger workers had to expend more energy in those situations, this may not stop them from foraging entirely. Furthermore, Riley et al. (1999) found that bumble bees have adapted to higher wind speeds by flying lower to the ground in windier conditions. Therefore, we may not have seen a size difference as workers show similar body stability and have developed behavioral adaptations during high wind speeds which allow all size spectra to forage in such situations.
Increasing cloud cover was linked to a higher number of large in comparison to small workers which may be due to the correlation between cloud cover and temperature. It is often warmer when there are no or little clouds, both of which were associated with an increased number of small foragers on bramble. Light intensity, on the other hand, had no effect on observed forager size. Other studies found that larger bumble bee workers were more likely to leave the nest in lower light intensities, due to their increased visual acuity as a result of bigger eyes (Kapustjanskij et al. 2007). The same pattern was also found in a study of stingless bees where larger species had longer foraging days due to their ability to fly earlier in the morning and later in the evening than smaller species (Streinzer et al. 2016). The lack of a significant association in our study was probably caused by the decreased light intensity range in comparison to these other studies. Kapustjanskij et al. (2007) recorded that the minimum light intensity for foraging is less than 10 lx, whereas our lowest documented light intensity was 12,600 lx. Thus, all size categories were presumably able to see and, hence, based on our results one should not consider light intensity an unimportant factor in predicting bumble bee worker size, as this may have been due to limitations in methodology rather than the lack of an existing relationship.
Lastly, there were significantly more active smaller workers in higher humidity in comparison to large ones. This was unexpected as humidity is often negatively correlated to temperature (Chambó et al. 2017), meaning that lower ambient temperatures are usually associated with higher air humidity. Moreover, smaller foragers are most commonly pollen foragers (Goulson et al. 2002) and as pollen is primarily collected in low humidity (Peat and Goulson 2005; Chambó et al. 2017) small workers should be less active with increasing humidity. Although the relationship was not as strong as the one seen for temperature, the trend was in the opposite direction, and we would have predicted to see more smaller workers when humidity was low. Nevertheless, due to the proximity to the coast we can expect higher than average humidity measurements at our sites, regardless of temperature (Pepler 2011). Thus, small workers may have adapted to high humidity conditions, but it may also the case that worker size is not solely affected by changes in weather conditions but also by other factors such as plant development, human interference, or differences in foraging efficiency of differently sized workers.
Visitation Rate
Temperature was a strong negative predictor of visitation rate which may be the result of an increased need to lose heat as temperatures increase. This is crucial to prevent overheating and may cause bumble bees to pause between visiting flowers, thereby decreasing the number of flowers visited per unit time. No other weather conditions significantly affected visitation rate but there was a negative correlation between the presence of pollen in the pollen baskets and their foraging speed. This is in line with results of previous studies which showed that foragers rarely forage solely for pollen (Goulson et al. 2002), meaning that they collect both nectar and pollen which increases the amount of time spent on each individual flower. The number of open flowers increased visitation rate which is expected as it decreases the need to fly between inflorescences when they are in crawling distance. This relationship may also explain the large difference in visitation rate between the two observation sites. Most bramble flowers at East Sands (site 2) started blooming within a day or two of each other whereas at the Botanical Garden (site 1) blooming onset occurred more steadily with only about 50 flowers blooming when observations started. Hence, inflorescences were further apart, and workers always had to take off to visit another flower which negatively affected visitation rate.
Although we did not expect cloud cover and light intensity to have major effects on the number of flower visits per minute, we were surprised about the lack of a significant relationship with wind speed. Based on observations during data collection, workers seemed to struggle to fly and land as wind speeds increased and some individuals were even blown off flowers or unable to reach an inflorescence at all. Moreover, previous research found that bumble bee workers were negatively affected by increasing wind speeds and it took them longer to approach flowers (Chang et al. 2016). The lack of a relationship may be due to the large number of open flowers, which, as explained above, decreases the need to fly between inflorescences which may have been linked to high visitation rates even during strong wind. Hence, wind may still negatively affect bumble bee foragers, specifically on plants where inflorescences are not in close proximity and studies looking into flight ability in the field rather than wind tunnels could help us to better understand the exact effects of wind on pollinators.
Individual Bumble Bee Species
While not a main focus of this study, we found that species prevalence was differently affected by certain aspects of weather. Interestingly, our results indicate that the invasive tree bumble bee Bombus hypnorum is more resistant to higher wind speeds and temperatures than native species. Bombus hypnorum was first documented in the UK in 2001 (Goulson and Williams 2001) and has since then rapidly spread across many parts of mainland England, Scotland and Wales (Crowther 2017). This rapid expansion of this species may be due their better adaptations to urbanization as they nest in above-ground cavities which are abundantly found in towns (Crowther et al. 2014) whereas ground-nesting sites are lacking (Fitch et al. 2019). Additionally, some hypotheses suggest that certain species are not as negatively affected by climate change, although the reasons for that are unclear (Crowther et al. 2014). Hence, further research into the adaptations of individual species to changes in local weather conditions may allow us to understand which physiological and behavioral traits are crucial to survival in a changing world.
Limitations and Further Studies
In this study, data was only collected at two sites within the same town over a short period of time. Thus, one should take care in generalizing the results to other species and habitats. Moreover, it would have been beneficial if the study had run throughout the same time as the colony cycle of most species. This would have given us an idea as to whether worker size changes throughout the colony life cycle which could have been incorporated as an explanatory factor in the models. However, such study would also need to consider a broader range of flowering species and aspects of flower choice, as there may be an interaction between local weather conditions, worker size and plant species.
References
Ahn C, Choe J, Um Y, Cho I, Yu I, Park J (1988) Effects of bee pollination and growth regulators treatment on preventing the malformation and accelerating the growth of strawberry fruit. Res Rep Rural Dev Adm Hortic 30(3):22–30
Aizen MA, Garibaldi LA, Cunningham SA, Klein AM (2009) How much does agriculture depend on pollinators? Lessons from long-term trends in crop production. Ann Bot 103:1579–1588
Alford DV (1975) Bumblebees. Davis-Poynter
Chambó ED, de Oliveira NTE, Garcia RC, Ruvolo-Takasusuki MCC, de Toledo VAA (2017) Statistical modeling of insect behavioral response to changes in weather conditions in Brassica napus L. Arthropod Plant Interact 11:613–621
Chang JJ, Crall JD, Combes SA (2016) Wind alters landing dynamics in bumblebees. J Exp Biol 219:2819–2822
Chen I-C, Hill JK, Ohlemüller R, Roy DB, Thomas CD (2011) Rapid range shifts of species associated with high levels of climate warming. Science 333:1024–1026
Cisarovsky GO (2013) Factors shaping the infection success of a bumblebee parasite. ETH Zurich
Clark MG, Bloxham DP, Holland PC, Lardy HA (1973) Estimation of the fructose diphosphatase–phosphofructokinase substrate cycle in the flight muscle of Bombus affinis Biochem J 134:589–597
Collins M, Knutti R, Arblaster J, Dufresne J-L, Fichefet T, Friedlingstein P, Gao X, Gutowski WJ, Johns T, Krinner G, Tebaldi MSC, Weaver AJ, Wehner M (2013) Long-term Climate Change: Projections, Commitments and Irreversibility: Climate Change 2013: The Physical Science Basis Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge
Combes SA, Dudley R (2009) Turbulence-driven instabilities limit insect flight performance. Proc Natl Acad Sci 106:9105–9108
Couvillon MJ, Dornhaus A (2009) Location, location, location: larvae position inside the nest is correlated with adult body size in worker bumble-bees (Bombus impatiens). Proc R Soc B Biol Sci 276:2411–2418
Crall J, Ravi S, Mountcastle AM, Combes SA (2015) Bumblebee flight performance in cluttered environments: effects of obstacle orientation, body size and acceleration. J Exp Biol 218:2728–2737
Crall J, Chang J, Oppenheimer R, Combes S (2017) Foraging in an unsteady world: bumblebee flight performance in field-realistic turbulence. Interface Focus 7:20160086
Crowther L (2017) The tree bumblebee, Bombus hypnorum: ecology and genetics of a naturally colonising pollinator. University of East Anglia
Crowther LP, Hein P-L, Bourke AF (2014) Habitat and forage associations of a naturally colonising insect pollinator, the tree bumblebee Bombus hypnorum. PloS One 9(9)
De Marco P Jr, Latini AO, Resende DC (2005) Thermoregulatory constraints on behavior: patterns in a Neotropical dragonfly assemblage. Neotrop Entomol 34:155–162
Del Castillo RC, Fairbairn DJ (2012) Macroevolutionary patterns of bumblebee body size: detecting the interplay between natural and sexual selection. Ecol Evol 2:46–57
Dirzo R, Young HS, Galetti M, Ceballos G, Isaac NJ, Collen B (2014) Defaunation in the anthropocene. Science 345:401–406
Dunn PK, Smyth GK (2018) Positive continuous data: gamma and inverse gaussian GLMs: Generalized linear models with examples in R. Springer, Berlin, pp 425–456
Feltham H, Park K, Minderman J, Goulson D (2015) Experimental evidence that wildflower strips increase pollinator visits to crops. Ecol Evol 5:3523–3530
Fitch G, Glaum P, Simao M-C, Vaidya C, Matthijs J, Iuliano B, Perfecto I (2019) Changes in adult sex ratio in wild bee communities are linked to urbanization. Sci Rep 9:1–10
Garbuzov M, Alton K, Ratnieks FL (2017) Most ornamental plants on sale in garden centres are unattractive to flower-visiting insects. PeerJ 5:e3066
Gérard M, Martinet B, Maebe K, Marshall L, Smagghe G, Vereecken NJ, Vray S, Rasmont P, Michez D (2020) Shift in size of bumblebee queens over the last century. Glob Change Biol 26:1185–1195
Goulson D, Williams P (2001) Bombus hypnorum (L.)(Hymenoptera: Apidae), a new British bumblebee? Brit J Entomol Nat Hist 14:129–131
Goulson D, Peat J, Stout JC, Tucker J, Darvill B, Derwent LC, Hughes WO (2002) Can alloethism in workers of the bumblebee, Bombus terrestris, be explained in terms of foraging efficiency? Anim Behav 64:123–130
Goulson D, Hanley ME, Darvill B, Ellis J, Knight ME (2005) Causes of rarity in bumblebees. Biol Conserv 122:1–8
Goulson D (2010) Bumblebees: behaviour, ecology, and conservation. Oxford University Press on Demand
Goulson D, Nicholls E, Botías C, Rotheray EL (2015) Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 347(6229):1255957
Hall K, Robert T, Gaston KJ, Hempel de Ibarra N (2021) Onset of morning activity in bumblebee foragers under natural low light conditions. Ecol Evol 11:6536–6545
Hallmann CA, Sorg M, Jongejans E, Siepel H, Hofland N, Schwan H, Stenmans W, Müller A, Sumser H, Hörren T (2017) More than 75% decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 12:e0185809
Heinrich B, Heinrich MJ (1983) Size and caste in temperature regulation by bumblebees. Physiol Zool 56:552–562
Heinrich B (1993) The hot-blooded insects. Springer-Verlag, Berlin
Heinrich B, Esch H (1994) Thermoregulation in bees. Am Sci 82:164–170
Heinrich B (2004) Bumblebee economics. Harvard University Press, Cambridge
Howe PD (2018) Perceptions of seasonal weather are linked to beliefs about global climate change: Evidence from Norway. Clim Change 148:467–480
IPCC (2021) Summary for policymakers: climate change 2021: The Physical Science Basis Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Masson-Delmotte, V, P Zhai, A Pirani, S L Connors, C Péan, S Berger, N Caud, Y Chen, L Goldfarb, M I Gomis, M Huang, K Leitzell, E Lonnoy, JBR Matthews, T K Maycock, T Waterfield, O Yelekçi, R Yu and B Zhou (eds)]. Cambride University Press, Cambride
Jenkins GJ, Perry MC, Prior MJ (2008) The climate of the United Kingdom and recent trends. Met Office Hadley Centre
Johnson SD, Anderson B (2010) Coevolution between food-rewarding flowers and their pollinators. Evol Educ Outreach 3:32–39
Kapustjanskij A, Streinzer M, Paulus H, Spaethe J (2007) Bigger is better: implications of body size for flight ability under different light conditions and the evolution of alloethism in bumblebees. Funct Ecol 21:1130–1136
Klein A-M, Vaissiere BE, Cane JH, Steffan-Dewenter I, Cunningham SA, Kremen C, Tscharntke T (2007) Importance of pollinators in changing landscapes for world crops. Proc R Soc B Biol Sci 274:303–313
May ML (1979) Insect thermoregulation. Annu Rev Entomol 24:313–349
Medler JT (1962) Morphometric studies on bumble bees. Ann Entomol Soc Am 55:212–218
Memmott J, Craze PG, Waser NM, Price MV (2007) Global warming and the disruption of plant–pollinator interactions. Ecol Lett 10:710–717
Newsholme E, Crabtree B, Higgins S, Thornton S, Start C (1972) The activities of fructose diphosphatase in flight muscles from the bumble-bee and role of this enzyme in heat generation. Biochem J 128:89–97
Peat J, Darvill B, Ellis J, Goulson D (2005) Effects of climate on intra-and interspecific size variation in bumble-bees. Funct Ecol 19:145–151
Peat J, Goulson D (2005) Effects of experience and weather on foraging rate and pollen versus nectar collection in the bumblebee, Bombus terrestris Behav Ecol Sociobiol 58:152–156
Pepler A (2011) Heat, humidity, and the El Niño—Southern Oscillation in Sydney. Australia. Aust Meteorol Oceanogr J 61:231
Philippides A, de Ibarra NH, Riabinina O, Collett TS (2013) Bumblebee calligraphy: the design and control of flight motifs in the learning and return flights of Bombus terrestris J Exp Biol 216:1093–1104
Pinchen BJ (2006) A pocket guide to the bumblebees of Ireland and Britain, 2 edn. Forficula Books
Potts SG, Biesmeijer JC, Kremen C, Neumann P, Schweiger O, Kunin WE (2010) Global pollinator declines: trends, impacts and drivers. Trend Ecol Evol 25:345–353
R Core Team (2021) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing
Riley J, Reynolds D, Smith A, Edwards A, Osborne J, Williams I, McCartney H (1999) Compensation for wind drift by bumble-bees. Nature 400:126–126
Robinson E, Cipullo M, Sousounis P, Kafali C, Latchman S, Higgs S, Maisey P, Mitchell L (2017) UK windstorms and climate change, an update to ABI research paper No. 19, 2009. Met Office
Roubik DW (1995) Pollination of cultivated plants in the tropics. Food & Agriculture Organization of the United Nations
Sanderson RA, Goffe LA, Leifert C (2015) Time-series models to quantify short‐term effects of meteorological conditions on bumblebee forager activity in agricultural landscapes. Agric For Entomol 17:270–276
Spaethe J, Weidenmüller A (2002) Size variation and foraging rate in bumblebees (Bombus terrestris). Insect Soc 49:142–146
Streinzer M, Huber W, Spaethe J (2016) Body size limits dim-light foraging activity in stingless bees (Apidae: Meliponini). J Comp Physiol A 202:643–655
Taylor GJ, Tichit P, Schmidt MD, Bodey AJ, Rau C, Baird E (2019) Bumblebee visual allometry results in locally improved resolution and globally improved sensitivity. Elife 8:e40613
Velthuis HH, Van Doorn A (2006) A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie 37:421–451
Venables WN, Ripley BD (2002) Modern Applied Statistics with S. Springer, Vol. Fourth
Warren M, Hill J, Thomas J, Asher J, Fox R, Huntley B, Roy D, Telfer M, Jeffcoate S, Harding P (2001) Rapid responses of British butterflies to opposing forces of climate and habitat change. Nature 414:65–69
Wickham H (2016) ggplot2: Elegant Graphics for Data Analysis. Springer-Verlag, New York
Wignall VR, Arscott NA, Nudds HE, Squire A, Green TO, Ratnieks FL (2020) Thug life: bramble (Rubus fruticosus L. agg.) is a valuable foraging resource for honeybees and diverse flower-visiting insects. Insect Conserv Diver 13:543–557
Williams I (1994) The dependence of crop production within the European Union on pollination by honey bees. Agric Zool Rev 6:229–257
Williams P (2007) The distribution of bumblebee colour patterns worldwide: possible significance for thermoregulation, crypsis, and warning mimicry. Biol J Linn Soc 92:97–118
Williams P, Colla S, Xie Z (2009) Bumblebee vulnerability: common correlates of winners and losers across three continents. Conserv Biol 23:931–940
Willmer PG, Finlayson K (2014) Big bees do a better job: intraspecific size variation influences pollination effectiveness. J Pollinat Ecol 14:244–254
Wilson EO (1971) The insect societies. Belknap Press, Cambridge
Wolf S, Rohde M, Moritz RF (2010) The reliability of morphological traits in the differentiation of Bombus terrestris and B. lucorum (Hymenoptera: Apidae). Apidologie 41:45–53
Yankit P, Rana K, Sharma HK, Kumar R, Nayak GMB, Negi N (2020) Foraging activities of bumble bee (Bombus haemorrhoidalis Smith) on Tomato (Solanum lycopersicum Mill.) under protected conditions and its correlation with weather parameters. J Entomol Zool Stud 8(4):2264–2268
Acknowledgements
We would like to thank the St Andrews Botanical Garden for providing access to their bramble patches.
Funding
Open Access funding enabled and organized by Projekt DEAL. This research was funded by the University of St Andrews, Scotland.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by Cassandra Uthoff. The first draft of the manuscript was written by Cassandra Uthoff and Graeme Ruxton commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of Interest
The authors have no competing interests to declare that are relevant to the content of this article.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Uthoff, C., Ruxton, G. Local Weather Conditions Affect Forager Size and Visitation Rate on Bramble Flowers (Rubus fruticosus) in Bumble Bees (Bombus spp). J Insect Behav 35, 17–30 (2022). https://doi.org/10.1007/s10905-022-09797-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10905-022-09797-1