
Abstract
Identifying early warning signs of sudden population changes and mechanisms leading to regime shifts are highly desirable in population biology. In this paper, a two-trophic ecosystem comprising of two species of predators, competing for their common prey, with explicit interference competition is considered. With proper rescaling, the model is portrayed as a singularly perturbed system with fast prey dynamics and slow dynamics of the predators. In a parameter regime near singular Hopf bifurcation, chaotic mixed-mode oscillations (MMOs), featuring concatenation of small and large amplitude oscillations are observed as long-lasting transients before the system approaches its asymptotic state. To analyze the dynamical cause that initiates a large amplitude oscillation in an MMO orbit, the model is reduced to a suitable normal form near the singular-Hopf point. The normal form possesses a separatrix surface that separates two different types of oscillations. A large amplitude oscillation is initiated if a trajectory moves from the “inner” to the “outer side” of this surface. A set of conditions on the normal form variables are obtained to determine whether a trajectory would exhibit another cycle of MMO dynamics before experiencing a regime shift (i.e. approaching its asymptotic state). These conditions serve as early warning signs for a sudden population shift as well as detect the onset of a regime shift in this ecological model.













Similar content being viewed by others
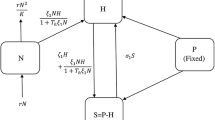
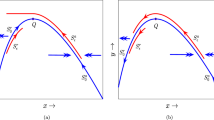
References
Ai S, Sadhu S (2020) The entry-exit theorem and relaxation oscillations in slow-fast planar systems. J Diff Equ 268:7220–7249
Albizuri JU, Desroches M, Krupa M, Rodrigues S (2020) Inflection, canards and folded singularities in excitable systems: application to a 3D FitzHugh - Nagumo model. J Nonlinear Sci 30:3265–3291
Baer SM, Erneux T (1986) Singular Hopf bifurcation to relaxation oscillations. SIAM J Appl Math 46:721–739
Baumgartner TR, Soutar A, Bartrina V (1992) Reconstruction of the history of Pacific sardine and Northern anchovy populations over the past two millennia from sediments of the Santa Barbara basin, California. CalCOFl Rep 33:24–40
Berglund N, Landon D (2012) Mixed-mode oscillations and interspike interval statistics in the stochastic FitzHugh - Nagumo model. Nonlinearity 25:2303–2335
Braaksma B (1998) Singular Hopf bifurcation in systems with fast and slow variables. J Nonlinear Sci 8(5):457–490
Brøns BM, Krupa M, Wechselberger M (2006) Mixed mode oscillations due to the generalized canard phenomenon, fields institute. Communications 49:39–63
Deng B (2001) Food chain chaos due to junction-fold point. Chaos 11:514–525
Deng B, Hines G (2002) Food chain chaos due to Shilnikov’s orbit. Chaos 12:533–538
Desroches M, Guckenheimer J, Krauskopf B, Kuehn C, Osinga HM, Wechselberger M (2012) Mixed-mode oscillations with multiple time scales. SIAM Rev 54:211–288
Desroches M, Krupa M, Rodrigues S (2013) Inflection canards and excitability threshold in neuronal models. J Math Biol 67:989–1017
Esper J, Bun̈tgen U, Frank DC, Nievergelt D, Liebhold A (2007) 1200 years of regular outbreaks in alpine insects. Proc R Soc B 274:671–679
Guckenheimer J (2008) Singular Hopf bifurcation in systems with two slow variables. SIAM J Appl Dyn Syst 7:1355–1377
Guckenheimer J, Holmes P (1983) Nonlinear oscillations, dynamical systems and bifurcations of vector fields. Springer-Verlag, Berlin
Guckenheimer J, Harris-Warrick R, Peck J, Willms AR (1997) Bifurcation, bursting, and spike frequency adaptation. J Comput Neurosci 4:257–277
Hastings A (2004) Transients: the key to long-term ecological understanding? Trends Ecol Evol 19(1):39–45
Hastings A, Abbott KC, Cuddington K, Francis T, Gellner G, Lai Y-C, Morozov A, Petrovskii S, Scranton K, Zeeman ML (2018) Transient phenomena in ecology. Science 361(6406):eaat6412
Krupa M, Popović N, Kopell N (2008) Mixed-mode oscillations in three time-scale systems: a prototypical example. SIAM J Appl Dyn Syst 7:361–420
Krupa M, Szmolyan P (2001) Relaxation oscillations and canard explosion. J Diff Equ 174:312–368
Krupa M, Szmolyan P (2001) Extending geometric singular perturbation theory to nonhyperbolic points - fold and canard points in two dimensions. SIAM J Math Anal 33(2):286–314
Krupa M, Wechselberger M (2010) Local analysis near a folded saddle- node singularity. J Diff Equ 248(2):2841–2888
Kuehn C (2015) Multiple time scale dynamics. Springer, NewYork
Kuwamura M, Chiba H (2009) Mixed-mode oscillations and chaos in a prey-predator system with dormancy of predators. Chaos 19:1–10
Kuznetsov YA (1998) Elements of applied bifurcation theory. Springer, NewYork
Liu W, Xiao D, Yi Y (2003) Relaxation oscillations in a class of predator-prey systems. J Diff Equ 188:30–331
Mujica J, Krauskopf B, Osinga HM (2018) Tangencies between global invariant manifolds and slow manifolds near a singular Hopf bifurcation. SIAM J Appl Dyn Syst 17(2):1395–1431
Muratori S, Rinaldi S (1989) Remarks on competitive coexistence. SIAM J Appl Math 49:1462–1472
Peet AB, Deutsch PA, Peacock - Lopez E (2005) Complex dynamics in a three-level trophic system with intraspecies interaction. J Theor Biol 232:491–503
Prescott SA, De Koninck Y, Sejnowski TJ (2008) Biophysical basis for three distinct dynamical mechanisms of action potential initiation. PLoS Comput Biol 4(10):e1000198
Rinaldi S, Muratori S (1992) Slow-fast limit cycles in predator-prey models. Ecol Modell 61:287–308
Sadhu S (2017) Stochasticity induced mixed-mode oscillations and distribution of recurrent outbreaks in an ecosystem. Chaos 27(3):033108
Sadhu S, Chakraborty Thakur S (2017) Uncertainty and predictability in population dynamics of a two-trophic ecological model: mixed-mode oscillations, bistability and sensitivity to parameters. Ecol Complex 32:196–208
Sadhu S, Kuehn C (2018) Stochastic mixed-mode oscillations in a three-species predator-prey model. Chaos 28(3):033606
Sadhu S (2021) Complex oscillatory patterns near singular Hopf bifurcation in a two-timescale ecosystem. Discret Contin Dyn Syst B 26(10):5251–5279
Scheffer M, Carpenter SR (2003) Catastrophic regime shifts in ecosystems: linking theory to observation. Trends Ecol Evol 18:648–656
Scheffer M (2009) Critical transitions in nature and society, vol 16. Princeton University Press, New Jersey
Szmolyan P, Wechselberger M (2001) Canards in \(\mathbb{R} ^3\). J Diff Equ 177:419–453
Acknowledgements
I would like to thank the two anonymous reviewers for their suggestions and Dr. Saikat Chakraborty Thakur for the numerous inspiring discussions. I am also grateful for having fruitful discussions at the workshops on “Human-Environment Systems: Feedback and Management” at the Fields Institute in March 2018 and “New Mathematical Methods for Complex Systems in Ecology” at BIRS, Canada in July 2019.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix
Appendix
1.1 Center manifold reduction
The center manifold can be expressed as a graph
where O(3) represents cubic and higher-order terms in u and v. The function \(\kappa \) can be determined by solving the equation \(\frac{d \kappa }{d \tau } = \delta (H_3 \kappa +\frac{1}{2}H_{11} u^2)+O(\delta ^2)\). Using the equation for w and the above equation and equating the coefficients of like terms, (see Braaksma 1998 for details) one obtains that
The corresponding equations in the center manifold up to higher order terms are
It is clear from the governing equations of the two-dimensional center manifold that the equilibrium (0, 0, 0) (up to \(O(\delta )\)) is asymptotically stable if and only if \(\alpha <0\), where \(\alpha \) is given by (6).
1.2 Special Cases of the normal form
The FSN II point \((\bar{x}, \bar{y}, \bar{z}, \bar{h})\) can be explicitly computed for the following special cases:
Equal predation efficiencies: When \(\beta _1=\beta _2\), one can solve for \(\bar{h}\) in terms of the other parameters and can compute \((\bar{x}, \bar{y},\bar{z})\). In this case,
where \(\alpha _{12}, \alpha _{21}, c, d\) are free parameters that satisfy (33)-(34) below:
Furthermore,
For biological significance, in addition to (33)- (34) we will choose \(\alpha _{12}, \alpha _{21}, c, d\) so that that the expression in the numerator of \(\bar{h}\) is positive, i.e.
No exclusive competition: Assuming that the predators do not exhibit interference competition, i.e. \(\alpha _{12}=\alpha _{21}=0\), one can explicitly solve for \((\bar{x}, \bar{y},\bar{z}, \bar{h})\), namely,
with
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Sadhu, S. Analysis of the onset of a regime shift and detecting early warning signs of major population changes in a two-trophic three-species predator-prey model with long-term transients. J. Math. Biol. 85, 38 (2022). https://doi.org/10.1007/s00285-022-01805-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00285-022-01805-4
Keywords
- Long-term transients
- Early warning signs
- Regime shift
- Mixed-mode oscillations
- Singular Hopf bifurcation
- Canard explosion
- Separatrix crossing