
Abstract
In this work, we have introduced an ecological model of a prey–predator system. It is assumed that the prey species grows logistically, but the total number of predator is constant in the time interval. Positivity and boundedness of the solution ensure that the proposed model is well-posed. Local stability conditions of the equilibrium points have been analysed by the Routh–Hurwitz criterion. The persistence of the system has also been shown under a parametric restriction. Numerical analysis has indicated that both axial and interior steady states can exist only for moderate consumption rate (searching efficiency). But if this rate becomes high (or low), then only the prey-free equilibrium (or one of the interior equilibriums) exists as a steady state. Further, the equilibrium points can change their stability through transcritical and saddle-node bifurcations by varying the consumption rate of the predator. Analytical results provide an interesting phenomenon about this model: the system can never show any oscillating behaviour for any parametric values, i.e. no limit cycle can occur through Hopf bifurcation around an equilibrium point. The axial equilibrium becomes stable from an unstable situation when the consumption rate becomes high and the interior state which is stable remains stable as time goes by.








Similar content being viewed by others
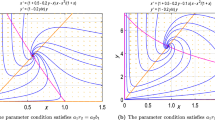
References
Freedman HI, Ruan S (1995) Uniform persistence in functional differential equations. J Differ Equ 115:173–192
Gilpin ME (1972) Enriched predator-prey systems: theoretical stability. Science 177:902–904
Hale JK (1977) Theory of functional differential equations. Springer, Heidelberg
Hethcote H, Wang W, Han L, Ma Z (2004) A predator–prey model with infected prey. Theor Popul Biol 66:259–268
Holling CS (1959) The components of predation as revealed by a study of small mammal predation of the European pine sawfly. Canad Entomol 91:293–320
Holt RD (1977) Predation, apparent competition, and the structure of prey communities. Theor Popul Biol 12:197–229
Holt RD, Lawton JH (1994) The ecological consequences of shared natural enemies. Ann Rev Ecol Syst 25:495–520
Jha PK, Ghorai S (2017) Stability of prey–predator model with holling type response function and selective harvesting. J Appl Comput Math 6:358. https://doi.org/10.4172/2168-9679.1000358
Kot M (2001) Elements of mathematical biology. Cambridge University Press, Cambridge
Kuznetsov Y, Rinaldi S (1996) Remarks on food chain dynamics. Math Biosci 134:1–33
Lotka A (1925) Elements of physical biology. Williams and Wilkins, Baltimore
Ma Z, Wang S, Wang T, Tang H (2017) Stability analysis of prey–predator system with Holling type functional response and prey refuge. Adv Differ Equ 2017:243. https://doi.org/10.1186/s13662-017-1301-4
Mondal S, Samanta GP (2019) Dynamical behaviour of a two-prey and one-predator system with help and time delay. Energ Ecol Environ. https://doi.org/10.1007/s40974-019-00141-6
Murray JD (1993) Mathematical biology. Springer, New York
Perko L (2001) Differential equations and dynamical systems. Springer, New York
Pielou EC (1977) Mathematical ecology. Wiley, New York
Saha S, Samanta GP (2019) Analysis of a predator–prey model with herd behaviour and disease in prey incorporating prey refuge. Int J Biomath 12(1):1950007
Shi C, Chen X, Wang Y (2017) Feedback control effect on the Lotka-Volterra prey-predator system with discrete delays. Adv Differ Equ. https://doi.org/10.1186/s13662-017-1410-0
Sugie J, Saito Y (2012) Uniqueness of limit cycles in a Rosenzweig–Macarthur model with prey immigration. Siam J Appl Math 72(1):299–316
van Baalen M, Krivan V, van Rijn PCJ, Sabelis MW (2001) Alternative food, switching predators, and the persistence of predator–prey systems. Am Nat 157(5):512–524
van Rijn PCJ, van Houten YM, Sabelis MW (2002) How plants benefit from providing food to predators even when it is also edible to herbivores. Ecology 83:2664–2679
Volterra V (1926) Variazioni e fluttuazioni del numero di individui in specie animali conviventi. Mem Accl Lincei 2:31–113
Xiao D, Ruan S (2001) Global analysis in a predator–prey system with nonmonotonic functional response. SIAM J Appl Math 61(4):1445–1472
Yodzis P (1998) Local trophodynamics and the interaction of marine mammals and fisheries in the Benguela ecosystem. J Anim Ecol 67:635–658
Acknowledgements
The authors are grateful to the anonymous referees, Prof. Santanu Ray, Associate Editor, and Prof. Bin Chen, Editor-in-Chief, for their careful reading, valuable comments and helpful suggestions, which have helped them to improve the presentation of this work significantly. The first author (Sangeeta Saha) is thankful to the University Grants Commission, India, for providing SRF.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Appendix
Appendix
Proof of Theorem 1
Proof
As the right-hand side of system (2) is continuous and locally Lipschitzian on space of continuous functions C, the solution (x(t), y(t)) of (2) with positive initial conditions \((x_{0},y_{0})\) exists and is unique on \([0,\tau ),\) where \(0<\tau \le +\infty \) (Hale 1977). From the first equation of (2), we have
Next we show that \(y(t)>0, \ \forall \, t\in [0,\tau )\). If it does not hold, then \(\exists t_{1} \in [0,\tau )\) such that \(y(t_{1})=0,\ {\dot{y}}(t_{1})\le 0\) and \(y(t)>0, \ \forall \, t\in [0,t_{1})\). From the \(2^{nd}\) equation of (2), we have
which is a contradiction to \({\dot{y}}(t_{1})\le 0\). So, \(y(t)> 0, \ \forall \, t\in [0,\tau )\). Hence, the theorem is proved. \(\square \)
Proof of Theorem 2
Proof
From the first equation of (2):
Let \(W= x(t)+ y(t)\).
So, all solutions of system (2) will enter into the region:
\(\square \)
Proof of Theorem 3
Proof
Choose \(\epsilon >0\), then \(\exists \ T_{1}>0,\) such that \(x(t)< {\overline{x}}+ \epsilon ,\ \forall \ t>T_{1}\).
Also, for \(0<\epsilon _{1}<{\underline{y}}-\frac{(1+2a)}{P\xi }, \ \exists \, \, T_{2}>0,\) such that \(y(t)> {\underline{y}}- \epsilon _{1}\), \(\forall \, t>T_{2}\).
Therefore, for all \(t> \max \{T_{1},T_{2}\}\), we have
Hence, \( \lim _{t\rightarrow \infty }x=0\). \(\square \)
Proof of Theorem 4
Proof
Let us consider the average Lyapunov function is \(V(x, y)=x^{\theta _{1}}y^{\theta _{2}}\) where each \(\theta _{i}\) for \(i=1,2\) is assumed to be positive. In the interior of \({\mathbb {R}}_{+}^{2},\) we have
To prove the permanence, we need to show \(\phi (x, y)>0\) at \(E_{1}\left( 0,\frac{\sigma }{\sigma +\zeta }\right) \). The value of \(\phi (x, y)\) at the boundary equilibria \(E_{1}\) is as follows:
\(E_{1}:\phi \left( 0,\frac{\sigma }{\sigma +\zeta }\right) = \frac{\theta _{1}}{\sigma +\zeta }[\sigma +\zeta -P\xi \sigma ]\).
Now, \(\phi \left( 0,\frac{\sigma }{\sigma +\zeta }\right) \) is positive only when \(\sigma +\zeta -P\xi \sigma >0\) holds. So, system (2) is permanent (Freedman and Ruan 1995; Mondal and Samanta 2019) if the stated condition is fulfilled. \(\square \)
Proof of Theorem 5
Proof
For \(E_{1}=\left( 0,\frac{\sigma }{\sigma +\zeta }\right) \):
The eigenvalues are the roots of the equation: \(\lambda ^{2}+P_{1}\lambda +P_{2}=0,\) where \(P_{1}=\sigma +\zeta +\frac{P\xi \sigma }{\sigma +\zeta }-1\) and \(P_{2}=(\sigma +\zeta )\left( \frac{P\xi \sigma }{\sigma +\zeta }-1\right) \).
By Routh–Hurwitz criterion, the roots have negative real parts if \(P_{1},P_{2}>0\). Now, \(P_{2}>0\) implies \(P\xi \sigma >\sigma +\zeta \) and this condition gives \(P_{1}>0\) also. \(\square \)
Proof of Theorem 6
Proof
For \(E^{*}_{L}=(x^{*}_{L},y^{*}_{L})\):
where \(b_{11}=-x^{*}_{L}+\frac{aP\xi x^{*}_{L}y^{*}_{L}}{(1+ax^{*}_{L})^{2}}; \ b_{12}=-\frac{P\xi x^{*}_{L}}{1+ax^{*}_{L}}; \ b_{21}=-\frac{K\xi y^{*}_{L}}{(1+ax^{*}_{L})^{2}}; \ b_{22}=-\frac{\sigma }{y^{*}_{L}}.\)
The eigenvalues are the roots of the equation: \(\lambda ^{2}+Q_{1}\lambda +Q_{2}=0,\) where \(Q_{1}\equiv Q_{1}(x^{*}_{L}, y^{*}_{L})=x^{*}_{L}+\frac{\sigma }{y^{*}_{L}}-\frac{aP\xi x^{*}_{L}y^{*}_{L}}{(1+ax^{*}_{L})^{2}}=\frac{\sigma }{y^{*}_{L}}+\frac{(1-a)x^{*}_{L}+2ax^{*2}_{L}}{1+ax^{*}_{L}}\)
and \(Q_{2}\equiv Q_{2}(x^{*}_{L}, y^{*}_{L})=\frac{\sigma }{y^{*}_{L}}\left\{ x^{*}_{L}-\frac{aP\xi x^{*}_{L}y^{*}_{L}}{(1+ax^{*}_{L})^{2}}\right\} -\frac{PK\xi ^{2}x^{*}_{L}y^{*}_{L}}{(1+ax^{*}_{L})^{3}}=\frac{\sigma x^{*}_{L}(1-a+2ax^{*}_{L})}{y^{*}_{L}(1+ax^{*}_{L})}-\frac{K\xi x^{*}_{L}(1-x^{*}_{L})}{(1+ax^{*}_{L})^{2}}\).
By Routh–Hurwitz criterion, the roots have negative real parts if \(Q_{1},Q_{2}>0\).
Now \(x^{*}_{L}<1\). So, \(Q_{2}>0\) implies \(1-a+2ax^{*}_{L}>0\) and this implies \(Q_{1}> 0\) too. \(\square \)
Proof of Theorem 8
Proof
For \(E^{*}_{L}(x^{*}_{L},y^{*}_{L}):\) let \(H(x,y)=\frac{1}{xy};\ h_{1}(x,y)=x(1-x)-\frac{P\xi xy}{1+ax}\ {{\text {and}}}\ h_{2}(x,y)=\sigma (1-y)-\frac{K\xi xy}{1+ax}-\zeta y\).
Now, \(H(x,y)>0\) in the interior of positive quadrant of x–y plane. At \(E^{*}_{L}\):
Hence, \(\Delta (x,y)\) does not change sign and is not identically zero in \(\Omega \). So, by Bendixson–Dulac criterion, there exists no limit cycle in \(\Omega \). Therefore, if \(E^{*}_{L}(x^{*}_{L},y^{*}_{L})\) is locally asymptotically stable, then it will be globally asymptotically stable in \(\Omega \) in the positive quadrant of x–y plane. \(\square \)
Proof of Theorem 10
Proof
Let \(\xi _{[TC]}\) be the value of \(\xi \) such that \(J|_{E_{1}}\) has a simple zero eigenvalue at \(\xi =\xi _{[TC]}\).
So, at \(\xi =\xi _{[TC]}:\)
Here, \(\lambda _{1}=-(\sigma +\zeta )<0\).
After some calculations: \(V=\left( -(\sigma +\zeta ), \frac{K\xi \sigma }{\sigma +\zeta }\right) ^{{{\mathrm{T}}}}=(v_{1},v_{2})\ {{\text {and}}} \ W=(1,0)^{{{\mathrm{T}}}}\).
Therefore,
By Sotomayor’s theorem, system (2) undergoes a transcritical bifurcation around \(E_{1}\) at \(\xi =\xi _{[TC]}\). \(\square \)
Proof of Theorem 11
Proof
where \(b_{11}=-{\overline{x}}^{*}+\frac{aP\xi {\overline{x}}^{*}{\overline{y}}^{*}}{(1+a{\overline{x}}^{*})^{2}}; \ b_{12}=-\frac{P\xi {\overline{x}}^{*}}{1+a{\overline{x}}^{*}}; \ b_{21}=-\frac{K\xi {\overline{y}}^{*}}{(1+a{\overline{x}}^{*})^{2}}; \ b_{22}=-\frac{\sigma }{{\overline{y}}^{*}}\).
The eigenvalues are the roots of the equation: \(\lambda ^{2}+Q_{1}\lambda +Q_{2}=0,\) where \(Q_{1}=-(b_{11}+b_{22})={\overline{x}}^{*}+\frac{\sigma }{{\overline{y}}^{*}}-\frac{aP\xi {\overline{x}}^{*}{\overline{y}}^{*}}{(1+a{\overline{x}}^{*})^{2}}=\frac{\sigma }{{\overline{y}}^{*}}+\frac{(1-a){\overline{x}}^{*}+2a{\overline{x}}^{*2}}{1+a{\overline{x}}^{*}}\)
and \(Q_{2}=(b_{11}b_{22}-b_{12}b_{21})=\frac{\sigma }{{\overline{y}}^{*}}\left\{ {\overline{x}}^{*}-\frac{aP\xi {\overline{x}}^{*}{\overline{y}}^{*}}{(1+a{\overline{x}}^{*})^{2}}\right\} -\frac{PK\xi ^{2}{\overline{x}}^{*}{\overline{y}}^{*}}{(1+a{\overline{x}}^{*})^{3}}=\frac{\sigma {\overline{x}}^{*}(1-a+2a{\overline{x}}^{*})}{{\overline{y}}^{*}(1+a{\overline{x}}^{*})}-\frac{K\xi {\overline{x}}^{*}(1-{\overline{x}}^{*})}{(1+a{\overline{x}}^{*})^{2}}\).
At \(\zeta =\zeta _{\left[ {{\mathrm{Sn}}}\right] },\ Q_{2}=Det\left( J|_{{\overline{E}}^{*}}\right) =0\). Then the characteristic equation for \({\overline{E}}^{*}\) reduces to \(\lambda ^{2}-Tr\left( J|_{{\overline{E}}^{*}}\right) \lambda = 0\). So, \(J|_{E^{*}}\) has a simple zero eigenvalue and a eigenvalue with negative real part at \(\zeta =\zeta _{\left[ {{\mathrm{Sn}}}\right] }\).
At \(\zeta =\zeta _{\left[ {{\mathrm{Sn}}}\right] }\):
where \(A=K\xi {\overline{x}}^{*}\frac{(1-{\overline{x}}^{*})}{(1+a{\overline{x}}^{*})^{2}}\). Here, \(V=(1,0)^{{{\mathrm{T}}}}\ {{\text {and}}}\ W=(1,-1)^{{{\mathrm{T}}}}\).
Thus, using Sotomayor’s theorem, system (2) undergoes a saddle-node bifurcation around \({\overline{E}}^{*}\) at \(\zeta =\zeta _{\left[ {{\mathrm{Sn}}}\right] }\). \(\square \)
Rights and permissions
About this article

Cite this article
Saha, S., Samanta, G.P. Local dynamics of a predator–prey community in a moderate period of time. Energ. Ecol. Environ. 5, 47–60 (2020). https://doi.org/10.1007/s40974-019-00146-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40974-019-00146-1