
Abstract
An important strategy in the discovery of biological mechanisms involves the piecing together of experimental results from interventions. However, if mechanisms are investigated by means of ideal interventions, as defined by James Woodward and others, then the kind of information revealed is insufficient to discriminate between modular and non-modular causal contributions. Ideal interventions suffice for constructing webs of causal dependencies that can be used to make some predictions about experimental outcomes, but tell us little about how causally relevant factors are organized together and how they interact with each other in order to produce a phenomenon. I argue that lab research relies on more elaborated types of interventions targeting in a controlled fashion multiple variables at the same time in order to probe the temporal organization of causally-relevant factors along distinct causal pathways and to test for non-modular interaction effects, thus providing crucial spatial-temporal constraints guiding the formulation of more detailed mechanistic explanations.


Similar content being viewed by others
Notes
A mechanism is a set of “entities and activities organized such that they are productive of regular changes from start or set-up to finish or termination conditions” (Machamer et al. 2000, p. 3); “a structure performing a function in virtue of its component parts, component operations, and their organization [...] responsible for one or more phenomena” (Bechtel and Abrahamsen 2005, p. 423); “entities and activities organized in such a way that they are responsible for the phenomenon (McKay Illari and Williamson 2012, p. 124); “a complex system which produces that behavior by the interaction of a number of parts according to direct causal laws” (Glennan 1996, p. S344) or by interactions that “can be characterized by direct, invariant, change relating generalization” (Glennan 2002, p. 52).
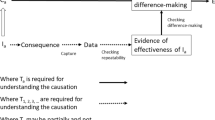
For the purposes of this paper, I rely on the observation that interventions play an indispensable role in the elucidation of mechanisms, as illustrated in the scientific practice of molecular biology and related fields. I leave aside the more speculative question of figuring out whether mechanistic productivity best describes the ultimate nature of causation while interventions play a strictly epistemic role in the discovery and confirmation of mechanistic explanations, or whether mechanisms presuppose a more primitive notion of causation as difference-making; the issue is debated in (Bogen 2004; Glennan 2010, 2011; Hall 2004; Psillos 2004; Waskan 2011; Woodward 2011).
Machamer, Darden and Craver (2000, p. 18) define a mechanism sketch as “an abstraction for which bottom out entities and activities cannot (yet) be supplied or which contains gaps in its stages. The productive continuity from one stage to the next has missing pieces, black boxes, which we do not yet know how to fill in. A sketch thus serves to indicate what further work needs to be done [...].”
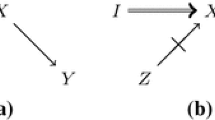
That is, without directly changing Y, or any other variable along the causal pathway from X to Y, or by simultaneously intervening on convergent causal pathways leading to Y. A fourth condition dictates that the intervention must be capable of overriding uncontrolled influences (act as a ‘switch’), such that its effect on the output conditions can be detected (Craver 2007, pp. 96–97; Woodward 2003, pp. 94–99).
This partial description will suffice to illustrate a philosophical point. The mechanism is in fact more complex, and monitors both the presence of lactose and glucose, triggering the expression of the lac operon only when bacteria live in an environment rich in lactose but poor in glucose; a more complete description can be found in (Griffiths et al. 2007, p. 307).
Many authors have challenged the view that mechanisms operate in a strictly regular manner and regularly succeed in producing the phenomena for which they are responsible (Andersen 2012; Bogen 2005; DesAutels 2011; Glennan 2010). Nevertheless, in scientific practice, an interventionist approach requires reproducible events: “if phenomena are infrequent to the point that they amount to irreproducible observations and experimental results, they are indistinguishable from the background noise of accidental happenings, thus making it impossible to distinguish phenomena generated by irregular mechanisms from chance correlations, as well as to interpret the results of experimental interventions required to demonstrate the causal contribution of mechanisms to the phenomena for which they are allegedly responsible” (Baetu 2013, p. 254). On this account, reproducibility amounts to the ability to produce a phenomenon with a consistent rate of success per number of experimental trials, such that, by conducting an adequate number of trials, one can determine whether an intervention on a given variable has or doesn’t have an effect on the phenomenon. The requirement for regularity can be reduced to a minimum as the number of trials increases, but cannot be completely eliminated.
Isopropyl \(\beta \)-d-1-thiogalactopyranoside (IPTG) is a non-hydrolyzable (and therefore non-metabolizable) lactose analog capable of binding the lacI repressor and cause it to detach from the operator side. In lactose-induced cells, lacZ/\(\beta \)-galactosidase expression is turned on, then, as lactose is degraded by \(\beta \)-galactosidase, expression is turned back off. In IPTG-induced cells, this negative feedback loop is absent. Note that mechanisms involving cyclical causal pathways, such as feedback loops, are investigated using the same multi-variable interventions; for an example, consult Baetu (2012).
Complementation refers to a situation where a combination of mutations yields a wild-type phenotype (Benzer 1955; Lewis 1951). Note that complementation posits an immediate threat to the modularity assumption. If the causal contribution of the mechanistic components affected by the mutations is modular, then modifying one component, or the second, or both should lead to the same loss in the ability of the mechanism to produce the wild-type phenotype. Yet this is not what happens. Each separate mutation results in a loss of the wild-type phenotype, while a double mutation results in a wild-type phenotype. One way to account for complementation effects is to hypothesize an interaction between the two components, such that if any single component is modified, it fails to interact with the other—for instance, because their geometrical shapes don’t fit anymore according to a key and lock model of molecular interaction; however, if the two are simultaneously modified, it can happen that a new geometrical fit is produced and they can once again interact.
Mutations in the gene encoding a repressor protein (lacI) reveal that this protein is trans-acting; that is, it can act on any copy of the target DNA site in the cell (Fig. 2c, row 6). In contrast, operator (o) mutations reveal that such a site is cis-acting; that is, it regulates the expression of an adjacent transcription unit on the same DNA molecule (row 7).
This hypothesis was subsequently supported by in vitro binding assays coupled with DNase treatment demonstrating an overlap between the DNA sequences covered by lacI and RNA polymerase (Majors 1975).
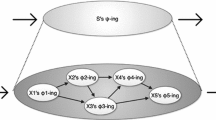
Levels are understood here as levels of composition (Wimsatt 1976), namely mechanisms and modules being composed of molecular component parts.
Alternative views include an ontic view, which treats mechanistic explanations as objective features of the world (Craver 2007; Salmon 1984), and a modified epistemic view according to which mathematical models provide a more rigorous understanding of some quantitative-dynamic details of phenomena by showing that they are consequences of rules and assumptions about the operation of mechanisms (Baetu 2015a, b; Bechtel 2012; Bechtel and Abrahamsen 2011; Braillard 2010; Brigandt 2013; Gross 2015).
References
Andersen, H. (2012). The case for regularity in mechanistic causal explanation. Synthese, 189(3), 415–432.
Baetu, T. M. (2012). Filling in the mechanistic details: Two-variable experiments as tests for constitutive relevance. European Journal for Philosophy of Science, 2(3), 337–353.
Baetu, T. M. (2013). Chance, experimental reproducibility, and mechanistic regularity. International Studies in History and Philosophy of Science, 27(3), 255–273.
Baetu, T. M. (2015a). From mechanisms to mathematical models and back to mechanisms: Quantitative mechanistic explanations. In P. A. Braillard & C. Malaterre (Eds.), Explanation in biology. An enquiry into the diversity of explanatory patterns in the life sciences (pp. 345–363). Dordrecht: Springer.
Baetu, T. M. (2015b). When is a mechanistic explanation satisfactory? Reductionism and antireductionism in the context of mechanistic explanations. In G. Sandu, I. Parvu, & I. Toader (Eds.), Romanian studies in the history and philosophy of science. Dordrecht: Springer.
Bechtel, W. (2012). Understanding endogenously active mechanisms: A scientific and philosophical challenge. European Journal for Philosophy of Science, 2, 233–248.
Bechtel, W. (2006). Discovering cell mechanisms: The creation of modern cell biology. Cambridge: Cambridge University Press.
Bechtel, W. (2008). Mental mechanisms: Philosophical perspectives on cognitive neuroscience. New York: Routledge.
Bechtel, W., & Abrahamsen, A. (2005). Explanation: A mechanist alternative. Studies in History and Philosophy of Biological and Biomedical Sciences, 36, 421–441.
Bechtel, W., & Abrahamsen, A. (2011). Complex biological mechanisms: Cyclic, oscillatory, and autonomous. In C. A. Hooker (Ed.), Philosophy of complex systems (pp. 257–285). New York: Elsevier.
Bechtel, W., & Richardson, R. (2010). Discovering complexity: Decomposition and localization as strategies in scientific research. Cambridge, MA: MIT Press.
Benzer, S. (1955). Fine structure of a genetic region in bacteriophage. Proceedings of the National Academy of Science, 41, 344–354.
Bogen, J. (2004). Analyzing causality: The opposite of counterfactual is factual. International Studies in the Philosophy of Science, 18, 3–26.
Bogen, J. (2005). Regularities and causality; generalizations and causal explanations. Studies in History and Philosophy of Biological and Biomedical Sciences, 36, 397–420.
Braillard, P.-A. (2010). Systems biology and the mechanistic framework. History and Philosophy of Life Sciences, 32, 43–62.
Brigandt, I. (2013). Systems biology and the integration of mechanistic explanation and mathematical explanation. Studies in History and Philosophy of Biological and Biomedical Sciences, 44(4), 477–492.
Callebaut, W., & Rasskin-Gutman, D. (2005). Modularity: Understanding the development and evolution of natural complex systems. Cambridge, MA: MIT Press.
Cartwright, N. (2002). Against modularity, the causal Markov condition and any link between the two: Comments on Hausman and Woodward. British Journal for the Philosophy of Science, 53(3), 411–453.
Casini, L. P., McKay Illary, P., Russo, F., & Williamson, J. (2011). Models for prediction, explanation and control: Recursive Bayesian Networks. Theoria. An International Journal for Theory, History and Foundations of Science, 70(1), 5–33.
Craver, C. (2007). Explaining the brain: Mechanisms and the Mosaic unity of neuroscience. Oxford: Clarendon Press.
Craver, C., & Darden, L. (2013). In search of biological mechanisms: Discoveries across the life sciences. Chicago, IL: University of Chicago Press.
Darden, L. (2006). Reasoning in biological discoveries: Essays on mechanisms, interfield relations, and anomaly resolution. Cambridge: Cambridge University Press.
DesAutels, L. (2011). Against regular and irregular characterizations of mechanisms. Philosophy of Science, 78(5), 914–925.
Gilbert, J., & Müller-Hill, B. (1967). The lac operator is DNA. Proceedings of the Aristotelian Society, 58(6), 2415–2421.
Gilbert, W., & Müller-Hill, B. (1966). Isolation of the lac represor. Proceedings of the Aristotelian Society, 56(6), 1891–1898.
Glennan, S. (1996). Mechanisms and the nature of causation. Erkenntnis, 44, 49–71.
Glennan, S. (2002). Rethinking mechanistic explanation. Philosophy of Science, 69, S342–S353.
Glennan, S. (2010). Ephemeral mechanisms and historical explanation. Erkenntnis, 72, 251–266.
Glennan, S. (2011). Singular and general causal relations: A mechanist perspective. In P. McKay, J. Williamson, & F. Russo (Eds.), Causality in the sciences (pp. 789–817). Oxford: Oxford University Press.
Griffiths, A., Wessler, S., Lewontin, R., & Carroll, S. (2007). Introduction to genetic analysis (9th ed.). New York: W. H. Freeman.
Gross, F. (2015). The relevance of irrelevance: Explanation in systems biology. In P.-A. Braillard & C. Malaterre (Eds.), Explanation in biology. An enquiry into the diversity of explanatory patterns in the life sciences. Dordrecht: Springer.
Hall, N. (2004). Two concepts of causation. In J. Collins, N. Hall, & L. Paul (Eds.), Causation and counterfactuals (pp. 225–276). Cambridge, MA: MIT Press.
Jacob, F., & Monod, J. (1961). Genetic regulatory mechanisms in the synthesis of proteins. Journal of Molecular Biology, 3, 318–356.
Lewis, E. B. (1951). Pseudoallelism and gene evolution. Cold Spring Harbor Symposia on Quantitative Biology, 16, 159–174.
Machamer, P. (2004). Activities and causation: The metaphysics and epistemology of mechanisms. International Studies in the Philosophy of Science, 18(1), 27–39.
Machamer, P., Darden, L., & Craver, C. (2000). Thinking about mechanisms. Philosophy of Science, 67, 1–25.
Majors, J. (1975). Initiation of in vitro mRNA synthesis from the wild-type lac promoter. Proceedings of the Aristotelian Society, 72(11), 4394–4398.
McKay Illari, P., & Williamson, J. (2012). What is a mechanism? Thinking about mechanisms across the sciences. European Journal for Philosophy of Science, 2(1), 119–135.
McKay, P., & Williamson, J. (2011). What is a mechanism? Thinking about mechanisms across the sciences. Journal for Philosophy of Science, 2(1), 119–135.
Mitchell, S. (2008). Exporting causal knowledge in evolutionary and developmental biology. Philosophy of Science, 75, 697–706.
Mitchell, S. (2009). Unsimple truths: Science, complexity, and policy. Chicago: University of Chicago Press.
Morange, M. (1998). A history of molecular biology. Cambridge, MA: Harvard University Press.
Pearl, J. (2000). Causality: Models, reasoning, and inference. Cambridge: Cambridge University Press.
Psillos, S. (2004). A glimpse of the secret connexion: Harmonizing mechanisms with counterfactuals. Perspectives on Science, 12, 288–391.
Salmon, W. (1984). Scientific explanation and the causal structure of the world. Princeton: Princeton University Press.
Simon, H. (1962). Architecture of complexity. Proceedings of the American Philosophical Society, 106(6), 467–482.
Wagner, G., & Altenberg, L. (1996). Perspective: Complex adaptations and the evolution of evolvability. Evolution, 50, 967–976.
Waskan, J. (2011). Mechanistic explanation at the limit. Synthese, 183(3), 389–408.
Weber, M. (forthcoming). On the incompatibility of dynamical biological mechanisms and causal graphs. Philosophy of Science.
Wimsatt, W. C. (1972). Complexity and organization. In K. F. Schaffner & R. S. Cohen (Eds.), PSA 1972, proceedings of the philosophy of science association. Dordrecht: Reidel.
Wimsatt, W. C. (1976). Reductive eplanation: A functional account. In A. C. Michalos (Ed.), Boston studies in the philosophy of science (pp. 671–710). Dordrecht: Reidel.
Woodward, J. (2002). What is a mechanism? A counterfactual account. Philosophy of Science, 69, S366–S377.
Woodward, J. (2003). Making things happen: A theory of causal explanation. Oxford: Oxford University Press.
Woodward, J. (2010). Causation in biology: stability, specificity, and the choice of levels of explanation. Biology and Philosophy, 25, 287–318.
Woodward, J. (2011). Mechanisms revisited. Synthese, 183(3), 409–427.
Woodward, J. (2013). Mechanistic explanation: Its scope and limits. Proceedings of the Aristotelian Society, 87(1), 39–65.
Woodward, J., & Hitchcock, C. (2003). Explanatory generalizations, part I: A counterfactual account. Nous, 37, 1–24.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Baetu, T.M. From interventions to mechanistic explanations. Synthese 193, 3311–3327 (2016). https://doi.org/10.1007/s11229-015-0930-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11229-015-0930-y