
Abstract
Despite the large and growing number of Miocene fossil catarrhine taxa, suitable common ancestors of great apes and humans have yet to be agreed upon. Considering a) the conservative and primitive nature of the hominoid molar cusp pattern, and b) the variability of secondary dental features, it is difficult to discern whether a hominoid dentition is primitive, secondarily simplified to the primitive condition or too far derived to be ancestral to any of the living forms. Nonetheless, the inability to recognize a common ancestor is primarly due to the absence of a model of hominoid differentiation that provides a basis for its recognition. Vertical climbing as the limiting component of cautious climbing, explains all of the locomotor anatomy shared by living hominoids. Comparison of the shared derived characters of hominoids to those of forms which have converged on hominoidsi.e colobines, atelines, lorisines, paleopropithecines and sloths suggest that early hominoids were probably folivores. In arboreal forms there is a strong link between a large body size, folivory and cautious climbing. Comparison of craniodental characters of committed folivores to committed frugivores from among each of the compared groups with the exception of lorisines, indicates that many of the distinguishing craniodental characters of humans and great apes are adaptations to folivory. Many of these characters, however, are also present in Jolly's seed eating complex. As such folivory may be the heritage factor which Jolly hypothesized to account for differential reduction of canines in fossilTheropithecus and hominids.
Similar content being viewed by others


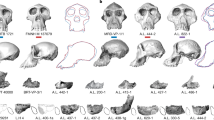
References
Aiello A., 1978.Sloth hair unanswered questions. In (G.G. Montgomery ed.).The ecology of arboreal folivores. pp. 213–218. Washington DC: Smitsonian Institution Press.
Ankle F., 1966.Morphologie von Wiberlsaule und Brustkorb. Biblio. Primatol. no. 4, Basel: Karger
Andrews P.J. and Tekkaya, 1980.A revision of the turkish Miocene Hominoid Sivapithecus metai. Palaeontology 23:85–95.
Ashton E.H. and Oxnard C.E., 1964.Functional adaptations in the primate shoulder girdle. Proc. Zool. Soc. London. 142:49–66.
Baldwin L.H. and Teleki G., 1976.Patterns of gibbon behavior on Hall's Island Bermuda. In (D.M. Rumbaugh ed.). Gibbon and Siamang vol 4 pp. 21–105 Basel: S. Karger.
Bauchop T., 1978.Digestion of leaves in vertebrate arboreal folivores. In (G.G. Montgomery ed.). The ecology of arboreal folivores, pp. 193–204. Washington DC: Smithsonian Institution Press.
Bauchop T. and Martucci R.W., 1968.Ruminant-like digestion of the langur monkey. Science 161:698–700.
Beebe W., 1927.The three toed sloth Bradypus cuculliger cuculliger Zoologica vol. 7 #1
Bishop N.H., 1979.Himalayan langurs: Temperate colobines. J. Hum. Evol. 8:251–281.
Bleisch W., Cheng A., Ren X., and Xie J., 1993.Preliminary results from a field study of wild Guizhou snub-nosed monkeys, Rhinopithecus brelichi. Folia primatol. 60: 72–82.
Britton S.W., 1941.Form and function in the sloth. Quart. Rev. Biol. 16:13–34, 190–207
Cant J.H., 1986.Locomotor and feeding postures of spider and howling monkeys: Field study and evolutionary interpretation. Folia Primatol. 49:90–105.
Carpenter C.R. and Durham M.N., 1969.A preliminary description of suspensory behavior in nonhuman primates. Proc. 2nd Int. Congr. Primatol. Atlanta, vol. 2 pp. 147–154. Basel: Karger.
Cartmill M., 1974.Pads and claws in arboreal primate locomotion. In (F.A. Jenkins Jr. ed.). Primate Locomotion, pp. 45–83. New York: Academic Press.
Cartmill M., 1979.The volar skin of primates: Its frictional characteristics and their functional significance. Amer. J. Phys. Anthrop. 50: 497–510.
Cartmill M. and Milton K., 1977.The lorisiform wrist joint and the evolution of brachiating adaptions in the Hominoidae. Amer. J. Phys. Anthrop. 47: 249–272.
Chalmers Mitchel P., 1905.On the intestinal tract of mammals Trans. Zool. Soc. London 17:437–536.
Charles-Dominique P., 1974.Ecology and feeding behavior of five sympatric species of lorisids in Gabon. In (R.D. Martin, G.A. Doyle, and A.C. Walker ed.) Prosimian Biology, pp. 131–150. Pittsburgh: University of Pittsburgh press.
Charles-Dominique P., 1977.Ecology and Behavior of Nocturnal Primates, New York: Columbia University press.
Chapman C., 1987.Flexibility in the diets of three species of Costa Rican primates. Folia Primatol. 40:90–105.
Chivers D.J., 1974.The Siamang in Malaysia. A Field Study of a Primate in a Tropical Rain Forest. Contr. Primatol. vol. 4 Basel: Karger.
Chivers D.J., 1977.The lesser apes. In (H.S. Rainier and G.H. Bourne, eds.) Primate Conservation, pp. 539–598.
Clutton-Brock T.H. and Harvey P.H., 1977.Primate ecology and social organization. Journal of Zoology (London). 183:1–39.
Conroy G.C. and Fleagle J.G., 1972.Locomotor behaviors in living and fossil pongids. Nature 237: 103–104.
DeBonis L. and Melentis J., 1980.Nouvelles remarques sur l'anatomie d'un primate hominoide du Miocene Ouranopithecus macedoniensis Implications sur la Phylogenie des hominoides. Cr. Acad. Sci. Paris, 290:755–758.
Defler T.R., 1987.Ranging and the use of space in a group of woolly monkeys (Lagothrix lagothricha) in the NW Amazon of Columbia. Int. J. Primatol. 8:420.
Deniker J., 1885.Recherches anatomiques et embryologiques sur les singes anthropoides: Foetus de Gorille et de gibbon. Arch Zool. Exp. Gen. ser. 2, 3. Suppl 1–265
Deniker J. and Boluart H. 1886.Les sacs laryngiens des singes anthropoides. Journal de l'Anatomie et de la Physiologie. Paris 22:51–62.
Dixson A.F., 1982.The Natural History of the Gorilla. New York: Columbia Univ. press.
Doran D.M., 1989.Chimpanzee and pygmy chimpanzee positional behavior: The influence of body size, morphology and ontogeny on locomotion and posture. Ph.D. Thesis, Stony Brook: SUNY.
Doran D.M., 1992.Comparison of instantaneous and locomotor bout sampling methods: A case study of adult male chimpanzee locomotor behavior and substrate use. Am. J. Phys. Anthrop. 89:85–99
Doyle G.A. and Bearder K.S., 1977. In (H.S. Rainier and G.H. Bourne, eds.)Primate Conservation, pp. 2–37. New York: Academic press
Dykyj D., 1980.Locomotion of the slow loris in a designed substrate context. Amer. J. Phys. Anthrop. 52:577–586.
Eaglen R.H., 1984.Incisor size and diet revisited: The view from a platyrrhine perspective. Am. J. Phys. Anthrop. 64:263–275
Eggeling von H., 1896.Zur Morphologie des Dammmuskulatur. Morph. Jahrb. Bd. 24:311–631.
Eggeling von H., 1920.In wie weit is der Wurm fortsatz am mensclichen blindarm ein rudimentares Gebilde. Anat. Anz. Bd. 53:401–428.
Eisenberg J.F., 1977.Comparative ecology and reproduction of New World monkeys. In (D.G. Kleiman, ed.) The Biology and Conservation of the Callithricidae, pp. 13–22. Washington D.C. Smithsonian Inst. Press.
Eisenberg J.F. and Maliniak, E. 1978.Reproduction by two-toed sloth Choloepus hoffmani in captivity. Amer. Assoc. of Mammalogist Abstr. Tech. Pap. 58th Annual meeting pp. 41–42.
Elftman H., 1932.Evolution of the pelvic floor of primates. Am. J. Anat. 51:307–346.
Elftman J.F. and Atkinson W.B., 1950.The abdominal viscera of the gorilla. In (W.K. Gregory ed.) Anatomy of the Gorilla, pp. 197–204. New York: Columbia University press.
Ellefson J.O., 1974.A natural history of white-handed gibbons in the Malayan penninsula. In (D.M. Rumbaugh ed.) Gibbon and Siamang, vol. 3, pp. 1–136. Basel: Karger.
Erikson G.E., 1963.Brachiation in New World monkeys and in anthropoid apes. Symp. Zool. Soc. London 10: 135–164.
Estrada A., 1984.Resource use by Howler monkeys in the rain forest of los Tuxtlas, Vera Cruz, Mexico. Int. J. Primatol. 5: 105–131.
Etter H.F., 1973.Terrestrial adaptations in the hands of the cercopithecinae. Folia primato. 20: 331–350.
Etter, H.F., 1974.Morphologisches und metrisches vergleichende Untersuchung am Handskelett rezenten Primaten. Gegenb. Morpholo. Jb., 120: 153–171.
Fick R., 1895.Vergleichend anatomische studien an einem erwachsenen orang-utang. Arch. Anat. Physiol., Anat Abth., 1–96.
Fleagle J.G., 1976a.Locomotion and posture of the Malayan siamang and implications for hominoid evolution. Folia Primatol. 26: 245–269.
Fleagle J.G., 1976b.Locomotion, posture and habitat utilization in two sympatric Malaysian leaf monkeys (Presbytis obscura and Presbytis melalophus). Yrb. Phys. Anthrop. 20:243–251.
Fleagle J.G., 1977.Locomotor behavior and muscular anatomy of sympatric Malaysian leaf monkeys. Amer. J. Phys. Anthrop. 46:297–307.
Fleagle J.G., Stern J.T., Jungers W.L., Susman R.I., Vangor A.K. and Wells J., 1981.Climbing a biomechanical link with brachiation and bipedalism. Symp. Zool. Soc. London 48: 359–373.
Fooden J., 1963.A revision of the woolly monkeys (genus Lagothrix). J. Mammal. 44:213–247
Fooden J., 1964.Stomach contents and gastrointestinal proportions in wild shot Guianan monkeys Amer. J. Phys. Anthrop. 22:227–232
Ford S.M., 1980.A systematic revision of the platyrrhinni based on features of the postcranium. Ph.D. Thesis, Univ. of Pittsburgh.
Ford S.M. and Davis L.C., 1992Systematics and body size: Implications for feeding adaptations in New World monkeys. Amer. J. Phys. Anthrop. 88(4): 415–468.
Forster A., 1917.Die muscle contrahentes und interosei manus in der saugetierreihe und beim menschen. Arch. Anat. Physiol. Anat. Abt. 101–378.
Forster A., 1918.Zur anatomie des primaren peritoneumfalten des Calcalapparates und der recessus ileo-calcalis. Arb. Anat. Inst. Weisbaden 56:1–170
Fossey D., 1974.Observations on the home range of one group of mountain gorillas (Gorilla gorilla beringei). Anim. Behav. 22:568–581.
Fossey D. and Harcourt A.H. 1977.Feeding ecology of free-ranging mountain gorillas (Gorilla gorilla beringei). In (T.H. Clutton-Brock ed.)Primate Ecology, pp. 415–499. London: Academic press.
Galidikas-Brindamour B.M.F., 1979.Orang-utan adaptation at Tanjung Puting Reserve: Mating and Ecology. In (D.R. Hamburg and E.R. McCown eds.).The Great Apes, pp. 194–233. Menlo Park: Benjamin Cummings.
Gaulin S.J.C., 1979.A Jarman/bell model of primate feeding niches. Human Ecol. 7:1–19.
Gaulin S.J.C. and Konner, M., 1977.On natural diet of primates, including humans. In (R.J. Wurtman and J.J. Wurtman eds.). Nutrition and the Brain, pp. 1–86. New York: Raven press.
Gaulin S.J.C. and Gaulin C.K., 1982.Behavioral ecology of Alouatta seniculus in Andean cloud forrest Int. J. Primat. 3:1–32.
Gautier-Hion A., 1978.Food niches and coexistence in sympatric primates in Gabon. In (D.J. Chivers and J. Herbert eds.). Recents Advances in Primatology vol. 1 Behavior, pp. 269–286. New York: Academic press.
Gautier Hion A. and Emmons, L.H. and Pobost, G., 1980.A comparison of the diet of 3 major groups of primary consumers of Gabon (Primates squirrels and ruminants) Oecologia (Berlin) 45:182–189
Gebo D.L., 1986.The anatomy of the prosimmian foot and its application to the primate fossil record. Ph.D. thesis. Duke University.
Gebo D.L., 1989.Postcranial adaptations and evolution in the lorisidae. Primates 30(3):347–367.
Gittleman J.L. and Thompson S.D., 1988.Energy allocation in mammalian reproduction. Am. Zool. 28: 863–868.
Gittins J.P. and Raemakers J.J., 1980.Siamang, lar and agile gibbons. In (D.J. Chivers ed.). Malayan Forrest Primates, pp. 63–105 New York: Plenum press.
Glander K.E., 1978.Howling monkey feeding behavior and plant secondary compounds: a study of strategies. In (G.G. Montgomery ed.) The ecology of arboreal folivores. pp. 561–574. Washington DC: Smithsonian Institution Press.
Glander K.E., 1982.The impact of plant secondary compounds on primate feeding behavior. Yrbk. of Phys. Anthrop. 25:1–18.
Goodall J., 1986.The chimpanzees of Gombe Patterns of Behavior. Cambridge: Harvard Univ. Bellknap press.
Godfrey L.R., 1991.How big were the giant lemurs of Madagascar? Am. J. Phys. Anthrop. Suppl. (Abstract) 12:80.
Godfrey L.R. Simons, E.L., Chatrath, P.S., and Rakotosamimanana B., 1990.A new fossil lemur (Babakoia, Primates) from Northern Madagascar. C.R.S. Acad. Sci. Paris 310:81–87.
Gordon K.D., 1980.Dental attrition in the chimpanzee (Pan troglodytes verus): a scanning electron microscope study. Ph.D. Thesis, Yale.
Greenfield L.O. and Washburn, A. 1991.Polymorphic aspects of male anthropoid canines. Amer. J. Phys. Anthrop. 84(1):17–34.
Greenfield L.O., 1992.A non-adaptive dental trait. Int. J. Primatol. 13:631–657.
Gregory W.K., 1922.Origin and Evolution of the Human Dentition. Baltimore: Williams and Wilkins.
Gregory W.K., 1934.Man's place among the anthropoids. Oxford: Clarendon Press.
Gregory, W.K. and Raven, H.C., 1937.In Quest of Gorillas. New Bedford, Mass: Darwin Press,
Harcourt A.H., Fossey D., Stewart K.T., and Watts D.D., 1980Reproduction in wild gorillas and some comparisons with chimpanzees. J. Reprod. Fert., Suppl. 28: 59–70.
Hasewaga M., Kishino H. and Yano T., 1984.Estimation of branching dates among primates by molecular clocks of nuclear DNA which slow down in Hominoidea. J Hum. Evol. 18:461–476.
Hasewaga M., Kishino H. and Yano T. 1985.Dating of the human ape split by a molecular clock of mitochondrial DNA. J. Hum. Evol. 22:16–174.
Hildwein G. and Goffart M., 1975.Standard metabolism and thermoregulation in a prosimian Perodicticus potto. Comp. Biochem. Physio. A: Comp. Physio. 50:201–213.
Hill C.A., 1967.A note on the gestation period of the siamang Hylobates syndactylus.Int. Zoo Yrbk. 7:93–94.
Hill W.C.O., 1953.Primates: Comparative Anatomy and Taxonomy. I. Strepsirrhini. Edingburgh: Edingburgh Univ. Press.
Hill W.C.O., 1962.Primates: Comparative Anatomy and Taxonomy. V. Cebidae Part B. Edingburgh: Edingburgh Univ. Press.
Hill W.C.O., 1970.Primates: Comparative Anatomy and Taxonomy, VII. Cynopithecidae, New York: Wiley.
Hladik C.M., 1977.Chimpanzees of Gabon and chimpanzees of Gombe: some comparative data on the diet In (T.H. Clutton-Brock ed.). Primate Ecology, pp. 481–501. New York: Academic press.
Hladik C.M., 1978.Adaptive strategies in primates in relation to leaf eating. In (G.G. Montgomery ed.). The ecology of arboreal folivores, pp. 215–254. Washington DC: Smitsonian Instituition Press.
Hladik C.M. and Hladik A., 1972.Disponibilities alimentaires et domaines vitaux des primates a Ceylon. Terre Vie. 26:149–215
Holme E.C., Pesole G., and Saccone C., 1989.Stochastic models of molecular evolution and estimated phylogeny of nucleotide substitutions in the hominoid primates. J. Hum. Evol. 18:775–799.
Hooton E.A., 1946.Up From The Ape. New York: MacMillan.
Hunt K.D., 1989.Positional behavior in Pan troglodytesat the Mahale mountains and the Gombe stream national parks, Tanzania. Ph.D. Thesis. Univ. of Michigan.
Huntington G.S., 1903.The anatomy of the Human Peritoneum and Abdominal Cavity Considered from the Standpoint of Development and Comparative Anatomy. New York: Philadelphia press.
Huxley T.H., 1964.The structure and classification of the Mammalia. The Medical Times and Gazette. Vol. 1 and Vol. 2
Izard M.K., 1987.Lactation length and three species of galago. Am. J. Phys. Anthrop.
Izard M.K. and Rasmussen T.D., 1985.Reproduction in the slender loris (Loris tardigradus malabaricus) Amer. J. Primat. 8:153–165.
Izawa K., 1987.Group sizes and composition of primates in the upper Amazon basin. Primates 17:367–399.
Jablonski N.G., 1992.Upper limb morphology and locomotion in the Yunnan snub-nosed langur. Rhinopithecus bieti. Am. J. Phys. Anthrop. Suppl. (Abstract) 14:94.
Jacobs-Lagen E., 1923.Das Schgleimhaut relief des Prosimien-Rumpfdarma mit beitrager zur kenntnis der Kerkringschen Faltensysteme der Anthropoiden und des Menschen. Jan. Zschr. Naturwies. 64:1–90.
Jansen D.H., 1978.Complications in interpreting the chemical defenses of trees against tropical arboreal planteating vertebrates. (In G.G. Montgomery ed.). The ecology of arboreal folivores, pp. 73–84. Washington DC: Smitsonian Institution Press.
Jenkins F.A. Jr., 1970.Anatomy and function of expanded ribs in certain edentates and primates. J. Mammal. 51:288–301.
Jenkins F.A. Jr., 1981.Wrist rotation in primates: a critical adaptation for brachiators. Symp. Zoo. Soc. London vol. 48, pp. 429–451.
Jolly C.J., 1970.The seed-eaters: A new model of hominid differentiation based on a baboon analogy. Man 5:5–26.
Jolly C.J., 1972.The classification and natural history of Theropithecus (Simopithecus)(Andrews 1916) baboons of the African Plio-pleistocene. Bull. Brit. (Nat. Hist.) Mus. Geol. 22:1–122.
Jolly C.J. and Gorton, J.T., 1974.Proportions of the extrinsic foot muscles in some lorisoid prosimians. In (R.D. Martin, G.A. Doyle, and A.C. Walker, eds.). Prosimian Biology, pp. 801–811 Pittsburgh: University of Pittsburgh press.
Jones C. and Sabater Pi J., 1971.Comparative ecology of Gorilla gorilla (Savage and Wyman) and Pan troglodytes (Blumenbach) in Rio Muni, West Africa. Biblio. Primat. No. 13 Karger, Basel
Jouffroy F.K. and Lessertisseur J., 1959.Reflexions sur les muscles contracteurs des doigts et des ortiels (Contrahentes digitorum) chez les primates. Ann. des Sc. Nat. Zool., Ser. 12, 1:211–235.
Jouffroy F.K. and Lessertisseur J., 1960.Les specialisations anatomiques de la main chez les singes a progression suspendue. Mammalia 24:93–151.
Jungers W.L., 1977.Hindlimb and pelvic adaptations to vertical climbing and clinging in Megaladapis, a giant sub-fossil prosimian from Madagascar. Yrbk. Phys. Antrhop. 53:471–478.
Jungers W.L., 1978.The functional significance of skeletal allometry in Megaladapis in comparison to living prosimians. Amer. J. Phys Anthrop. 49:303–314.
Jungers W.L., Godfrey L.R., Simons E.L., Chatrath P.S. and Rakotosamimanana B., 1991.Phylogenetic and functional affinities of Babakotia (Primates) a fossil lemur from northern Madagascar. Proc. Nat. Acad. Sci. USA 88:9082–9086.
Juraniec J. and Szostakiewicz-Sawicka H., 1968.The central tendon of the diaphragm in primates. Folia Morphol. 27:183–194.
Kano T., 1983.An ecological study of the pygmy chimpanzees (Pan paniscus) of Yalosidi, Republic of Zaire. Int. J. Primatol. 4(1):1–31.
Kavanagh M. and Dresdale L., 1975.Observations on the wooly monkey (Lagothrix lagothricha) in Northern Colombia Primates 16:285–294.
Kay R.F., 1978.Molar structure and diet in extant Cercopithecidae. In (P.M. Butler and K.A. Joysey eds.). Development, Function and Evolution of Teeth, pp. 309–339. London: Academic Press.
Kay R.F., Fleagle J.G. and Simons E.L., 1981.A revision of the Oligocene apes of the Fayum Province Egypt. Am. J. Phys. Anthrop. 55:293–322
Kay R.F. and Hylander W.L., 1978.The dental structure of mammalian follivores with special reference to primates and phalangeroids (Marsupalia). In (G.G. Montgomery ed.). The ecology of arboreal folivores. pp. 173–192. Washington DC: Smithsonian Institution Press.
Kay R.F., Plavacan J.M., Glander K.E. and Wright P.C., 1988.Sexual selection and canine dimorphism in New World primates. Amer. J. Phys Anthrop. 77:385–397.
Keith A., 1903.The extent to which the posterior segments of the body have been transmuted and suppressed in the evolution of man and allied primates. J. Anat. Physiol. London. 37:18–40.
Keith A., 1912.Certain phases in the evolution of man. Brit. Med. J. 1:734–737, 788–790.
Keith A., 1923.Man's posture: Its evolution and disorders. Brit. Med. J. 1:451–54, 499–502, 545–48, 587–90, 624–26, 669–72.
Keith A., 1934.The Construction of Man's Family Tree. Watts, London.
Klatsch H., 1913.Die erwerbung der auftrehen haltung un ihre folgen. Anat. Anz. suppl. 44:161–186.
Klein L.L. and Klein D.J., 1977.Feeding Behavior of the Columbian spider monkey. In (T.H. Clutton-Brock ed.) Primate Ecology, pp. 154–181. London: Academic press.
Kortlandt A., 1968.Handgebrauch bei Freilebenden Shimpansen. In (B. Rensch ed.). Handgebrauch und Verstandigung bei Affen und Fruhmenschen, pp. 59–102. Bern: Hans Huber.
Kostanecki K., 1926.Le caecum des vertebres. Bull. Int. Acad. Cracovie Cl. Sci. Nat. Suppl.
Lamarck J.B., 1809.Philosophie Zoologique ou Exposition des considerations relative al'Histoire Naturelle des Animaux etc. Paris: J.B. Ballier.
Lamberton C.L., 1947.Contribution a la connaissance de la faune subfossile de Madagascar. Note XVI: Bradytherium ou Paleopropithique? Bull. Acad. Malagache (Nuov. Ser.) 26:89–140.
Langer P., 1988.The Mammalian Herbivore Stomach. Comparative anatomy, Function and Evolution. Stuttgart: Gustav Fischer.
Leakey R.E.F. and Walker A., 1985.New higher primates from the early Miocene Buluk, Kenya Nature 318: 173–175.
Leakgul B. and McNeely J.A. 1977.Mammals of Thailand. Bangkok li: Sahakarnbhat.
Leidel G., 1940.Zur Anatomie des Anthropmorphes herzen. Morph. Jb 84:457–490.
Lemos de Sa R.M. and Glander K.E., 1993.Capture techniques and morphometrics for the wooly spider monkey or muriqui (Brachyteles arachnoides). Am. J. Primatol. 29:145–153.
Leo Luna M., 1981.First field study of the yellow tailed wooly monkey Oryx 15:386–389
Lewis O.J., 1969.The hominoid wrist joint. Amer. J. Phys. Anthrop. 30:251–267.
Lessertisseur J., 1958.Doit-on distinguer deux plans de muscles interosseux a la main et au pied des primates? Ann. Sci. Nat. Zool. Biol. Anim. Ser. 11, 20:77–103.
Lineback P., 1933.The respiratory digestive and urinary system In (C. Hartman and W.L. Straus eds.) Anatomy of the Rhesus monkey pp 210–230. Baltimore: Williams and Wilkins.
Loghem van J.J., 1904.Das Colon und Mesocolon der Primaten. Petrus Camper Nederlandische Bijdragen tot de Anatomie 2:350–347.
Lorin-Epstein M.J., 1932.Evolution und Bedeutung des wurmfortsatzes und der valvula ileocecalis im Zusammanhang mit der Aufrichtung des Rumpfes. Zschr. Anat. und Entwick. 97:68–144.
Loth E., 1908.Die aponeurosis plantaris in der Primatereihe. Mit spezieller Berucksichtingung des Menschen. Morph. Jahrb. 38:194–322.
Lovejoy C.O., 1981.The origin of man. Science 211:341–351
MacArdle, J.E., 1981.Functional Morphology of the hip and thigh of the Lorisiformes. Contr. Primatol. vol. 17 Basel: Karger.
MacKenzie J.M.D., 1929.Food of the slow loris Nycticebus coucang. J. Bombay nat. Hist. Soc. (1929):971–1000
MacKinnon, J.R., 1974.The behavior and ecology of wild orang-utans (Pongo pygmaeus). Anim. Behav. 22:3–74.
MacKinnon J.R. and MacKinnon K.S., 1978.Comparative feeding ecology of 6 sympatric primates in West Malaysia. In (D.J. Chiver and J. Herbert eds). Recent Advances in Primatology, pp. 305–321. New York: Academic press.
Mac Phee R.D. and Vuillaume-Randriamanantana M., 1984.Hand and foot of Paleopropithecus, anatomy and function. Amer. J. Phys. Anthrop. Abstract 63:189.
Makintosh H.W., 1875.On the muscular anatomy of Choloepus didactylus. Proc. Roy. Irish Acad. 2nd ser. 2:66–78.
Martin R.D., 1990.Primate origins and evolution. New Brunswick: Princeton University Press.
Marzke M.W., 1971.The origin of the human hand. Amer. J. Phys. Anthrop. 34:61–84.
McHenry H.M. and Temerin L.A., 1979.The evolution of human bipedalism: Evidence from the fossil record. Yrbk. Phys. Anthrop. 22:105–131.
McKey D.B., 1978.Soils vegetation and seed eating by black colobus monkeys. In (G.G. Montgomery ed.). The ecology of Arboreal Folivores, pp. 423–427. Washington DC: Smitsonian Institution Press.
McKey D.B., Gartlan J.S., Watterman P.G. and Choo G.M., 1981.Food selection by black colobus monkey (Colobus satanas) in relation to plant chemistry. Biol. J. Linn. Soc. 16:115–146.
McKnab B.K., 1978.Energetics of arboreal folivores In (G.G. Montgomery ed.) The ecology of arboreal folivores, pp. 153–162. Washington DC: Smithsonian Institution Press.
McKnab B.K., 1980.Food habits and energetics and the population biology of mammals Am. Nat. 116:106–124.
McKnab B.K., 1986.The influence of food habits on the energetics of eutherian mammals. Ecol. Mono. 56(1): 1–19.
Meinke F., 1911.Morphologisches untersuchungen uber die myologie an den extremitaten bei Bradypus trydactylus. Morph. Jb. 42:309–360.
Mendel F.C., 1979.The wrist joint of two-toed sloths and its resemblance to brachiating adaptations of the Hominoidea. J. Morph. 162:413–424.
Mendel F.C., 1981a.Use of Hands and feet in two toed Sloths, Choloepus hoffmani during climbing and terrestrial locomotion. J. Mammalogy 62:413–421
Mendel F.C., 1981b.Hand of two-toed sloth: Its anatomy and potential use relative to the size of the support. J. Morphology 169:1–19.
Mendel F.C., 1985.Use of hands and feet in three toed Sloths, Bradypus tridactylus during climbing and terrestrial locomotion. J. Mammalogy 66:359–366
Midlo C., 1934.Form of hand and foot in primates. Am. J. Phys Anthrop. (o.s.) 19:337–389.
Miller R.A., 1939.Functional adaptation in the forelimb of the sloth J. Mammal. 16:38–51
Milton K., 1980.Foraging strategy of howler monkeys. New York: Columbia University press.
Milton K., 1984.Habitat, diet and activity patterns of free-ranging wooly spider monkeys (Brachyteles arachnoides E. Geoffroy 1906) Int. J. Primatol. 5:491–514.
Milton K., 1985.Multimale mating and absence of canine tooth dimorphism in wooly spider monkeys (Brachyteles arachnoides) Amer. J. Phys. Anthrop. 68:519–523.
Mittermeier R.A., 1978.Locomotor posture in Ateles geoffroy and Ateles paniscus. Folia primatol. 30:161–193.
Mittermeier R.A. and Coimbra-Filho A.F., 1977.Primate conservation in Brazilian Amazonia. In (H.S. Rainier and G.H. Bourne eds.). Primate Conservation, pp. 117–166. New York: Academic Press.
Mivart St. G., 1866.Contributions towards a more complete knowledge of the skeleton of primates. Part I The appendicular Skeleton of Simia. Trans. Zool. Soc. London (1886):175–225.
Mivart St. G., 1867.On the appendicular skeleton of primates. Phil. Trans. Roy. Soc. London. 157:299–429.
Mivart St. G. and Murie J. 1865.Observation on the anatomy of Nycticebus tardigradus. Proc. Zool. Soc. London. (1865):240–256
Montgomery G.G., and Sunquist M.E., 1978.Habitat selection and use by two-toed and 3 toed sloths. In (G.G. Montgomery ed.). The ecology of arboreal folivores, pp. 329–359. Washington DC: Smithsonian Instituition Press.
Morrison P.R., and Sinois J. Jr., 1962.Body temperature in two Brazilian primates. Boletin de Faculdad de Filosofia, Ciencias y Letras Universidad de Sao Paulo 24:167–178
Morgan E., 1972.The Descent of Woman. New York: Stein and Day
Morton D.J., 1924,Evolution of the human foot.II. Amer. J. Phys. Anthrop. 7:1–52.
Morton D.J. and Fuller D.D., 1952.Human Locomotion and Body Form. A study of Gravity and Man. Baltimore: Williams and Wilkins Co.
Muller E.F., 1979.Energy metabolism thermoregulation and water budget in the slow loris Nycticebus coucang Comp. Biochem physio. A: Comp. Physio. 64:109–119.
Muller E.F., Kazan J.M.Z. and Maloiy G.M.O., 1983.Comparative study of the basal metabolism and thermoregulation of a folivorous (Colobus guereza) and an omnivorous (Cercopithecus mitis) primate species. Comp. Biochem Physio. a: Comp. Physio 74:319–322.
Nadler R.D., Graham C.E. and Kling O.R., 1981.Postpartum amenorrhea and behavior of apes. In (C.E. Graham ed.). Reproductive Biology of the Great Apes, pp 69–81 New York: Academic press.
Nagy K.A. and Montgomery G.G., 1980.Field metabolic rate water flux and food consumption in three toed sloth B. variegatus. J. Mammal. 61:465–472.
Napier J.R., 1963.Brachiation and brachiators Symp. Zool. Soc. London 10:183–194
Napier J.R. and Napier P.H., 1967.A Handbook of Living Primates. New York: Academic press.
Narath A., 1901.Der bronchialbaum der Saugetheire in des Menschen. Bibl. Med. Stuttgart Abt A. Heft 3:1–380
Nash and Harcourt A.H., 1986.Social organization of galagos in Kenyan Wasta forrest. Amer. J. Primatol. 10: 357–69
Nicoll M.E. and Thompson S.D., 1987.Basal metabolic rates energetics and reproduction in Therian mammals marsupials and placentals compared. In (A.S. Loudon and P.A. Racey eds.) Reproduction Energetics in Mammals, pp. 7–24. Oxford: Clarendon press.
Nishimura A., 1990.A sociological and behavioral study of wooly monkeys Lagothrix lagothricha in the Upper Amazon. Sci. Eng. Rev. Doshisha University 31:87–121.
Nowack R.M., 1991.Walker's Mammals of the World. 5th ed. Baltimore: John Hopkins Univ. press.
Oates J.F., 1977a.The guereza and its food. In (T.H. Clutton-Brock ed.) Primate Ecology. pp. 276–321. London: Academic press.
Oates J.F., 1977b.The guereza and man. In (H.S. Rainier and G.H. Bourne, eds.), Primate Conservation, pp. 419–467. New York: Academic press.
Oates J.F., 1978.Water-plant and soil consumption by guereza monkeys (Colobus guereza): a relationship with mineral and toxins in the diet? Biotropica 10:241–253.
Owen R., 1835a.On the osteology of the chimpanzee and the orang-utan. Trans. Zool. Soc. London I:343–380.
Owen R., 1835b.On the saculate form of the stomach as it exits in the genus Semnopithecus. Trans. Zool. Soc. London I:65–70.
Parra R., 1978.Comparison of hindgut and foregut fermentation in herbivores. In (G.G. Montgomery ed.). The ecology of arboreal folivores, pp. 205–230. Washington DC: Smithsonian Instituition Press.
Peng Y., Ye Z., Zhang Y., Liu R., 1985.Observations on the position of the genus Rhinopithecus in phylogeny. Acta Theriol. Sinica 5:173–181.
Petter J.J. and Hladik C.M., 1970.Observation sur le domaine vital et la densite de population de Loris tardigradus dans le forest de Ceylon. Mammalia 34:394–309.
Polak C., 1908.Die anatomie des genus Colobus. Verh. Konniklijke Akad. Wetenshappen te Amsterdam II (14) 2:139–154.
Primrose A., 1899.The anatomy of the orang-utang (Simia satyrus). Trans. Roy. Soc. Canad. Inst. 6:507–598.
Prost J.H. 1965.A definitional system for the classification of primate locomotion. Amer. Anthrop 67:1198–1214.
Prost J.H., 1980.Origin of bipedalism. Amer. J. Phys Anthrop. 52:175–190.
Raven H.C., 1931.The gorilla the greatest of all apes. Nat. Hist. 31(3):231–242.
Remane A., 1960.Zahne und Gebiss. Primatologia III(2):637–846 Basel: Karger.
Richard A.F. and Nicoll M.E., 1987.Female social dominance and basal metabolism in Malagasy primates Propithecus verrauxi. Amer. J. Primat. 12:309–314.
Rijksen H.D., 1978.A Field Study on Sumatran Orangutans, Ecology Behavior and Conservation, Wagenigen.
Roberts D., 1974.Structure and function of the primate scapula. In (F.A. Jenkins ed.) Primate Locomotion, pp. 171–200. New York: Academic Press.
Robinson J.T., 1956.The dentition of the Australopithecinea, Transvaal Mus. Mem. 9.
Robinson J.T., 1972.Early Hominid Posture and Locomotion. Chicago: Univ. of Chicago.
Rodman P.S., 1973.Population composition and adaptive organization among orangutans of the Kutai Reserve. In (R.P. Michael and J.H. Crook eds.). Comparative Behavior and Ecology of Primates, pp. 171–209. New York: Academic Press.
Rodman P.S., 1979.Feeding behavior of orang-utans of the Kutai Nature Reserve East Kalima. In (D. Hamburg ed.) Perspectives of Human Evolution. Vol 4, pp 383–413 New York: Addison Wesley.
Rodman P.S., 1984.Foraging and social systems of orang-utans and chimpanzees. In (P.S. Rodman and J.H. Cant, eds.). Adaptations for Foraging in Non-Human Primates, pp. 134–160. New York: Columbia Univ. press.
Rodman P.S. and McHenry H.M., 1980.Bioenergetics and the origin of human bipedalism. Amer. J. Phys. Anthrop. 52:103–106.
Roonwal M.L. and Mohnot S.M., 1977.Primates of South Asia: Ecology, Sociobiology and Behavior. Cambridge: Harvard Univ. press.
Rose M.D., 1979.Positional behavior of natural populations: some quantitative results of a field study of Colobus guereza and Cercopithecus aethiops In (M.E. Morbeck, H. Preuschoft, N. Gomberg, eds.) Environment Behavior and Morphology. Dynamic Interactions in Primates, pp 75–94. New York: Fischer.
Rosenberger A.L., 1979.Phylogeny, evolution and classification of New World monkeys. Ph.D. Dissertation City Univ. New York.
Rosenberger A.L., 1992.Evolution of feeding niches in New World monkeys. Am. J. of Phys. Anthrop. 88(4): 525–562.
Rosenberger A.L. and Strier K.B., 1989.Adaptive radiations of the Ateline primates. J. Hum. Evol. 18:717–750.
Rudran R. 1973.The reproductive cycle of two species of langur with relation to environmental factors. Folia Primatol. 19:41–60.
Ruge G., 1892.Die Grenzlinien der Pleura-sacke un der Lagerung des Herzens bei Primaten inbesondere bei den Anthropoiden: Zuegnisse fur die metamere Verkurzung des Rumpfes. Morph. Jb. 19:149–249.
Ruge G., 1910.Grenzen der Pleurasacke der Affen und des Menschen. Morph. Jb. 41:1–109
Sarich V.M. and Wilson A.C., 1967Immunological time scale for hominoid evolution. Science 158:1200–1204.
Sarmiento E.E., 1983.The significance of the heel process in anthropoids. Int. J. Primatol. 4(2):127–152.
Sarmiento E.E., 1985.Functional differences in the skeleton of wild and captive orang-utans and their adaptive significance. Ph.D. Thesis New York University.
Sarmiento E.E., 1986.The evidence for terrestrial quadrupedalism in the Hominid lineage. Am. J. Phys. Anthrop. (Abstract) 66(1):226.
Sarmiento E.E., 1987.The phylogenetic position of Oreopithecus and its significance in the origin of the Hominoidea. Amer. Mus. Novitates #2881
Sarmiento E.E., 1988.Anatomy of the hominoid wrist joints, its evolutionary and functional implications. Int. J. Primatol. 9:281–345
Sarmiento E.E., 1991.Functional and phylogenetic implications of the differences in the pedal skeleton of australopithecines. Amer. J. Phys. Anthrop. (Abstract) 12:157–158.
Sarmiento E.E., 1994.Terrestrial traits in the hands and feet of gorillas. Amer. Mus. Nat. Hist. Novitates #3091.
Sawalaschin M., 1911.Der musculus flexor communis brevis digitorum pedis in der primatenreihe mit spezielier Berucksichtigung des Menschlichen varietaten. Morphol. Jahrb. 42:557–663.
Schaller G.B., 1963.The Mountain Gorilla: Ecology and Behavior Chicago: Univer. Chicago press.
Schon Ybarra M.A., 1968.The muscular system of the red howling monkey U.S. Nat. Mus. Bull. 273:1–185.
Schon Ybarra M.A., 1984.Locomotor postures of red howlers in deciduous forrest savanna. Amer. J. Phys. Anthrop. 63:65–76.
Schon-Ybarra M.A., 1986.Loud calls of adult male red howler monkeys (Alouatta seniculus). Folia primatol. 48:284–316.
Schultz A.H., 1930.The skeleton of the trunks and limbs of higher primates. Hum. Biol., 2:303–438.
Schultz A.H., 1936. Characters common to higher primates and characters specific to man.Quart. Rev. Biol. 11:259–283; 425–455.
Schultz A.H., 1937.Proportions, variability and assymetries of the long bones of the limbs and clavicles of man and apes. Hum. Biol., 9:281–328.
Schultz A.H., 1942.Growth and development of the proboscis monkey. Bull. Mus. Comp. Zool. 89:277–314.
Schultz A.H., 1953.The relative thickness of the long bones and the vertebrae in primates. Amer. J. Phys. Anthrop., 11:277–311.
Schultz A.H., 1956.Postembryonic age changes. Primatologia: 1:887–964.
Schultz A.H., 1960.Einege Beobachtungen und masse am skelet von Oreopithecus. Z. Morph. Anthropol. 50:136–149.
Schultz A.H., 1961.Vertebral column and thorax. Primatologia 4(Lf.5):1–66.
Schultz A.H., 1963.Relations between the lengths of the main part of the foot skeleton in primates Folia.Primat. 1:150–171.
Schultz A.H., 1968.The recent hominoid primates In (S.L. Washburn and P.C. Jay, eds.). Perspectives on Human Evolution 1, pp. 122–195. New York: Holt Rinehart and Winston, Inc.
Schwartz J.H. 1984.The evolutionary relationships of man and orang-utans. Nature 308:501–505.
Simons E.L., 1972.Primate Evolution. New York: MacMillan
Simons E.L., Godfrey L.R., Jungers W.L., Chatrath P.S., Rakotosamimanana B., 1992.A new subfossil lemur, Babakotia, and the evolution of the sloth lemurs. Folia Primatol. 58:197–203
Soini P., 1986.A synecological study of a primate community in the Pacaya-Saimiria National reserve Peru Primate Conserv. 7:63–71.
Sonntag C.F., 1921.A contribution to the anatomy of the three-toed sloth (Bradypus tridactylus). Proc. Zool. Soc. London. 21: 157–177.
Sonntag C.F., 1923.On the anatomy, physiology and pathology of the chimpanzee. Proc. Zool. Soc. London. 23: 323–429.
Sonntag C.F., 1924On the anatomy, physiology and pathology of the orang-outan. Proc. Zool. Soc. London. 24: 349–449.
Stern J.T., 1971.Functional myology of the hip and thigh of cebid monkeys and its implications for the evolution of erect posture. Biblioth. Primatol., 14: 1–318.
Stern J.T. and Susman R.L., 1981.Electromyography of the gluteal muscles in Hylobates, Pongo and Pan, implications for the evolution of hominoid bipedality. Amer. J. Phys. Anthro. 55: 153–166.
Strasser E., 1989.Form, function and allometry of the cercopithecoid foot. Ph.D. Thesis. New York: CUNY.
Strasser E., 1992.Hindlimb proportions, allometry and biomechanics in Old World monkeys. Amer. J. Phys. Anthrop. 87:187–213.
Straus W.L. Jr., 1930.The foot musculature of the highland gorilla. Quart. Rev. Biol. 5:261–317.
Straus W.L., 1934.The thoracic and abdominal viscera of primates with special reference to the orang-utan Proc. Am. Philo. Soc. 76:1–85.
Straus W.L., 1937.Cervical ribs in the Wooly monkey J. Mammal. 18:241–242.
Straus W.L., 1942.Rudimentary digits in primates. Quart. Rev. Biol. 17:228–243.
Straus W.L. and Wislocki G.B., 1932.On certain similarities between sloths and slow lemurs. Bull. Mus. Comp. Zool. Harvard. 74:45–56.
Strier K.B., 1991.Diet in one group of spider monkey or Muriquis (Brachyteles arachnoides) Amer. J. Primatol. 21:113–126.
Struhsaker T.T., 1975.The red Colobus monkey. Chicago: University of Chicago press
Struhsaker T.T., 1978.Food habits of five monkey species in the Kibale forrests Uganda. In (D.J. Chivers and J. Herbert eds.). Recents Advances in Primatology vol. 1 Behavior, pp. 269–286. New York: Academic press.
Struhsaker T.T., 1980.Comparison of the behavior and ecology of the red colobus and redtail monkey in the Kibale forrest Uganda. Afr. J. Ecol. 18:35–51.
Subramoniam S., 1957.Some observations on the habits of the slender loris Loris tardigradus (Linneaus). J. Bombay Nat. Hist. Soc. 54:387–398
Susman R.L., 1984.The locomotor behavior of Pan paniscus in the Lomako forrest. In (R.L. Sussman (ed.) The pygmy Chimpanzee: Evolutionary Biology and Behavior, pp. 369–393. New York: Plenum press.
Susman R.L., Stern J.T. Jr. and Jungers W.L., 1984.Arboreality and Bipedality in the Hadar Hominids. Folia primatol. 43:113–156.
Symmington M.M., 1987.Ecological and social correlates of party size in the black spider monkey Ateles paniscus chamek. Ph.D. Thesis Princeton University.
Symmington M.M., 1988a.Food composition and foraging party size in the black spider monkey Ateles paniscus chamek Behavior. 105:117–134.
Symmington M.M., 1988b.Demography ranging patterns and activity budgets of black spider monkeys Ateles paniscus chamek in the Manu National Park Peru. Am. J. Primatol. 15:45–67.
Szalay F.S. and Delson, E., 1979.Evolutionary History of the Primates. New York: Academic press.
Terborgh J., 1983.Five New World Primates. A study in Comparative Ecology. Princeton: Princeton University Press.
Thompson S.D. and Nicoll M.E., 1986.Basal metabolic weight and the energy of reproduction in therian mammals. Nature 321:690–696.
Treves F., 1885.Lectures on the anatomy of the intestinal canal and peritoneum in man. Brit. Med. J. I
Tutin C.E.G., 1980.Reproductive behavior of wild chimpanzees in the Gombe National park, Tanzania. J. Repro. Biol. 28:23–59.
Tutin C.E.G. and Fernandez M., 1985.Foods consumed by sympatric populations of Gorilla gorilla gorilla and Pan troglodytes troglodytes in Gabon, some preliminary data. Int. J. Primatol. 6:27–43.
Tuttle R.H., 1969a.Knuckle-walking and the problem of human origins. Science 166:953–961.
Tuttle R.H. 1969b.Quantitative and functional studies on the hands of the Anthropoidea. I. The Hominoidea. J. Morph. 128:309–363.
Tuttle R.H., 1969c.Terrestrial trends in the hands of the anthropoidea: A preliminary report. Proc. of the 2nd Int. Congr. Primatol. vol 2. pp. 192–200. Basel: Karger.
Tuttle R.H., 1970.Postural, propulsive and prehensile capabilities in the cheiridia of chimpanzees and other great apes. In (G.H. Bourne, ed.). The Chimpanzee Vol. II., pp. 167–253. Baltimore: Univer. Park Press.
Tuttle R.H. 1974.Darwin's Apes dental apes and the descent of man: normal science in evolutionary anthropology. Curr. Anthrop. 15:389–426.
Tuttle R.H. 1975a.Knuckle-walking and knuckle-walkers: a commentary on some recent perspectives of hominoid evolution. In (R.H. Tuttle ed.). Primate Functional Morphology and Evolution. pp. 203–212. The Hague: Mouton.
Tuttle R.H. 1975b.Parallelism, Brachiation and Hominoid Phylogeny. In (W.P. Luckett and F.S. Szalay eds.). Phylogeny of the Primates. pp. 447–480. New York. Plenum Press.
Tuttle R.H. 1981.Evolution of hominid bipedalism and prehensile capabilities. Phil. Trans. Roy. Soc. 292:89–94.
Tuttle R.H. and Beck B.B., 1972.Knucklewalking hand postures in an orang-utan Pongo pygmaeus. Nature 236:33–44.
Uhlman K., 1968.Huft und Oberschenkel-muskulatur systematische und vergleichenden Anatomie. Primatologia 4(10):1–442
Van Horn R.N. and Eaton G.C., 1979.Reproductive physiology and behavior in Prosimians. In (G.A. Doyle and R.D. Martin eds.) The Study of Prosimian Behavior pp. 79–122. New York: Academic press.
Van Roosmalen M.G.M., 1980.Habitat preferences diet feeding behavior and social organization of the black spider monkey Ateles paniscus chamek in Surinam. Ph.D. Dissertation, University of Wageningen, The Netherlands.
Vogel C., 1962.Morphologische studien am gesichtsschadel Catarrhiner Primaten. Biblo. Primatol. 4:1–226.
von Koenigswald G.H.R., 1952.Gigantopithecus blacki a giant fossil hominoid from the pleistocene of south China. Anthrop. Pap. Amer. Mus. Nat. Hist. 43:291–326.
Walker A.C., 1967.Locomotor Adaptations in Recent and Fossil Madagascan Lemurs. Ph.D. Thesis University of London.
Walker A. C., 1974.Locomotorn adaptations in past and present prosimian primates. In (F.A. Jenkins ed.). Primate Locomotion, pp. 349–381. New York: Academic press.
Walker P. and Murray, P., 1975.An assessment of masticatory efficiency in a series of anthropoid primates with special reference to the colobinae and the cercopithecinae. In (R. Tuttle ed.). Primate Functional Morphology and Evolution, pp 135–150. The Hague: Mouton.
Washburn S.L., 1950a.Thoracic viscera of the gorilla. In (W.K. Gregory ed.) Anatomy of the Gorilla, pp. 189–195. New York: Columbia University press.
Washburn S. L., 1950b.The specializations of man and his place among the catarrhine primates, Cold Harbor Symp. Quant. Biol. Vol. XV: Origin and Evolution of Man, pp. 37–52. Cold Spring Harbor, Long Island
Washburn S. L., 1963.Behavior and Human Evolution In (S.L. Washburn ed.). Classification and Human Evolution. pp. 190–201. Chicago: Aldine.
Washburn S.L., 1967.Behavior and the orgin of man: The Huxley memorial lecture. Proc. of the Roy. Anthrop. Inst. of Gr. Brtn and Irlnd. pp. 21–27.
Washburn S.L., 1968.Speculations on the problem of man's coming to the ground. In (B. Rothblatt ed.) Changing Perspectives on Human Evolution, pp. 193–206. Chicago: University of Chicago Press.
Washburn S.L., 1983.Audiotape recording of the concluding speech to the symposium on Hadar hominids, held April 1983 in Berkeley CA.Institute of Human Origins.
Wasser P.M., 1977.Feeding, ranging and group size in the Mangabey Cercocebus albigena. In (T.H. Cluttonbrock ed.). Primate Ecology. pp. 183–222. London: Academic press.
Wilson D.R., 1972.Tail reduction in Macaca. In (R.H. Tuttle ed.). Functional and Evolutionary biology of Primates, pp. 241–261 Chicago: Aldine-Atherton.
Wislocki G.B., 1928.Observations on the Gross and microscopic anatomy of the sloth (Bradypus grisseus grisseus andCholoepus hoffmani). J. Morphophysio. 46:317–377.
Wislocki G.B. and Straus W.L., 1932.On the blood-vascular bundles in the limbs of certain edentates and lemurs. Bull. Mus. Comp. Zool., 74:1–16.
Wood Jones F., 1929.Man's Place Among the Mammals. New York: Longmans, Green and Co.
Wrangham R.W., 1977.Feeding behavior of chimpanzees in Gombe National park Tanzania. In (T.H. Cluttonbrock ed.). Primate Ecology. pp. 504–538. London: Academic press.
Wu R., 1984.A revision of the classification of the Lufeng great apes. Acta Anthrop. Sinic. 6:265–271
Yalden D.W., 1972.The form and function of the carpal bones in some arboreally adapted primates. Acta Anat. 82:383–406.
Young A.L., Richard A.F., and Aiello L.C., 1990.Female dominance and maternal investment in strpsirrhin primates. Am. Nat. 135:473–488.
Ziemer L.K., 1978.Functional morphology of the forelimb joints in the woolly monkey Lagothrix lagotricha. Contr. Primatol. 14:1–130.
Zihlman A.L., 1969.Human locomotion a reappraisal of the functional and anatomical evidence. Ph.D. Thesis, University of California at Berkeley
Zihlman A.L. and Bunker L., 1979.Hominid bipedalism: Then and now. Yrb. Phys. Antrhop. 22:132–162.
Zingeser M.R., 1973.Dentition of Brachyteles arachnoides with reference to alouattine and Atelinine affinites. Folia primatol. 20:351–390.