
Summary
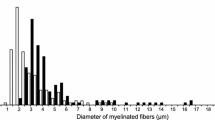
A significant reduction in the myelinated nerve fiber population was observed during quantitative electron-microscopic examination of peripheral nerves in chronic alloxan diabetic rats. Dystrophic axonal abnormalities and regenerating fibers were more numerous in diabetics than age-matched controls. Schwann cells showed reactive changes including prominent pi granules of Reich and intracytoplasmic filament accumulation. Enumeration of these alterations, however, revealed no singificant difference from controls. Endoneurial macrophages in diabetic rats were also filled with lamellar intracytoplasmic inclusions characteristic of a chronic neuropathy. Quantitation of pathologic lesions in teased nerve fibers confirmed the preponderance of axonal over demyelinative disease and showed demyelination to be segmental.
Microangiopathy was noted throughout the vasa nervorum of diabetic rats, and quantitative electron microscopy showed endothelial proliferation with doubling of the number of endothelial cells and proportional capillary mural thickening. Swollen, reactive endothelial cells appeared to effece the vascular lumen and may impair capillary perfusion. These microcirculatory changes, in the presence of biochemical and rheologic disturbances may contribute to tissue hypoxia and underly the loss of axons in experimental diabetic neuropathy.
Similar content being viewed by others


References
Behse F, Buchthal F, Carlsen F (1977) Nerve biopsy and conduction studies in diabetic neuropathy. J Neurol Neurosurg Psychiatry 40:1072–1082
Bischoff A (1980) Morphology of diabetic neuropathy. Horm Metab Res [Suppl] 9:18–28
Buja LM, Ferrans VJ, Levitskey S (1972) Occurrence of intramitochondrial glycogen in canine myocardium after prolonged anoxic cardiac arrest. J Mol Cell Cardiol 4:237–254
Carson KA, Bossen EH, Hanker JS (1980) Peripheral neuropathy in mouse hereditary diabetes mellitus. II. Ultrastructural correlates of degenerative and regenerative changes. Neuropathol Appl Neurobiol 5:361–374
Clements RS (1979) Diabetic neuropathy. New concepts of its etiology. Diabetes 28:604–611
Clements RS (1982) Pathogenesis of diabetic neuropathy. NY State J Med 82:864–872
Ditzel J (1980) Affinity hypoxia as a pathogenic factor of microangiopathy with particular reference to diabetic retinopathy. J Acta Endocrinol [Suppl] 238:39–55
Gabbay KH, Merola LO, Field RA (1966) Sorbitol pathway: presence in nerve and cord with substrate accumulation in diabetes. Science 151:209–210
Griffin JW, Price DL (1981) Segmental demyelination in experimental IDPN and hexacarbon neuropathies. Evidence for an axonal influence. Lab Invest 45:130–141
Grover-Johnson N, Baumann FG (1982) Neuropathology in diabetes: general aspects with emphasis on the nervous system. NY State J Med 82:860–863
Hanker JS, Ambrose WW, Yates PE, Koch GG, Carson KA (1980) Peripheral neuropathy in mouse hereditary diabetes mellitus. J Acta Neuropathol (Berl) 51:145–155
Hirano A, Dembitzer HM (1976) Eosinophilic rod-like structure in myelinated fibers of hamster spinal roots. Neuropathol Appl 2:225–232
Jakobsen J (1978) Peripheral nerves in early diabetes. Expansion of the endoneurial space as a cause of increased water content. Diabetologia 14:113–119
Johnson PC, Brendel K, Meezan E (1981) Human diabetic perincurial cell basement membrane thickening. Lab Invest 44:265–270
Lundborg G (1975) Structure and function of the intraneural microvessels as related to trauma, edema formation and nerve function. J Bone Joint Surg [Am] 57:938–948
Lundborg G (1979) The intrinsic vascularisation of human peripheral nerves — Structural and functional aspects. J Hand Surg 4:34–41
Lundborg G, Myers RR, Powell HC (1983) Increased endoneurial fluid pressure in experimental entrapment neuropathy. J Neurol Neurosurg Psychiatry 46:1119–1124
McMillan DE (1982) Further observations on serum visosity changes in diabetes mellitus. Metabolism 31:274–278
Miller JA, Pizzighella S, Gravallese E, Bunn HF (1980) Nonenzymatic glycosylation of erythrocyte membrane proteins. Relevance to diabetes. J Clin Invest 65:896–901
Myers RR, Powell HC, Shapiro HM, Costello ML, Lampert PW (1980) Changes in endoneurial fluid pressure permeability, and peripheral nerve ultrastructure in experimental lead neuropathy. Ann Neurol 8:392–401
Myers RR, Mizisin AP, Powell HC, Lampert PW (1982) Reduced nerve blood flow in hexachlorophene neuropathy. Relationship to elevated endoneurial fluid pressure. J Neuropathol Exp Neurol 41:391–399
Powell HC, Knox D, Lee S (1977) Alloxan diabetic neuropathy. Electron microscopic studies. Neurology (Minn) 27:60–66
Powell HC, Koch T, Garrett R, Lampert PW (1978) Schwann cell abnormalities in 2,5 hexanedione neuropathy. J Neurocytol 7:517–528
Powell HC, Ward HW, Garrett RS (1979) Glycogen accumulation in the nerves and kidneys of chronically diabetic rats. A quantitative electron-microscopic study. J Neuropathol Exp Neurol 38:114–127
Powell HC, Costello ML, Myers RR (1981) Galactose neuropathy. Permeability studies, mechanism of edema and mast cell abnormalities. Acta Neuropathol (Berl) 55:89–95
Powell HC, Ivor LP, Costello ML, Wolf PL (1982) Elevated hemoglobin A1 in streptozotocin diabetes and in rats with sucrose and galactose diets. Clin Biochem 15:133–137
Powell HC, Lampert PW (1983) Peripheral neuropathies. In: Rosenberg R, Schochet S (eds) Clinical neurosciences, chapt 9, sect III. Churchill Livingstone, Edinburgh Harlow New York, pp 325–362
Rosai J, Lascano EF (1970) Basophilic (mucoid) degeneration of the heart. Am J Pathol 61:99–115
Said G, Slama G, Selva J (1983) Progressive centripetal degeneration of axons in small fiber type diabetic polyneuropathy. A clinical and pathologic study. Brain 106:791–807
Sima AAF, Robertson DM (1979) Peripheral neuropathy in the diabetic mutant mouse. An ultrastructural study. Lab Invest 40:727–732
Sharma AK, Thomas PK, Gabriel G, Stolinski C, Dockery P, Hollins GW (1983) Peripheral nerve abnormalities in the diabetic mutant mouse. Diabetes 32:1152–1161
Spritz N, Singh H, Mariman B (1975) Decrease in myelin content of rabbit nerve with aging and diabetes. Diabetes 24:680–683
Stewart MA, Sherman WR, Anthony S (1966) Free sugars in alloxan diabetic nerve. Biochem Biophys Res Commun 22:488–491
Sugimura K, Dyck PJ (1980) Multifiber fiber loss in proximal sciatic nerve in symmetric distal diabetic neuropathy. J Neurol Sci 53:501–509
Thomas PK, Lascelles RG (1966) The pathology of diabetic neuropathy. Q J Med 35:489–509
Thomas PK, Elliason S (1984) Diabetic neuropathy. In: Dyck PJ, Thomas PK, Lambert EH (eds) Peripheral neuropathy, chap 76. Saunders, Philadelphia, PA, pp 1776–1810
Vital C, Brechenmacher C, Servise JM, Bellance R, Vital A, Dartigues JF, Boissieras P (1983), Ultrastructural study of peripheral nerve in arteritic diabetic patents. Acta Neuropathol (Berl) 61:225–231
Vracko R (1982) A comparison of the microvascular lesions in diabetes mellitus with those of normal aging. J Am Geriatr Soc 30:201–205
Yagihashi S, Matsunaga M (1979) Ultrastructural pathology of peripheral nerves in patients with diabetic neuropathy. Tohoku J Exp Med 129:357–366
Author information
Authors and Affiliations
Additional information
Supported in part by NS-14162 and NS-09053 from the National Institute for Neurological and Communicative Disorders and Stroke and the Veterans Administration Research Service
Rights and permissions
About this article
Cite this article
Powell, H.C., Myers, R.R. Axonopathy and microangiopathy in chronic alloxan diabetes. Acta Neuropathol 65, 128–137 (1984). https://doi.org/10.1007/BF00690466
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1007/BF00690466