
Abstract
Abstract relational concepts depend upon relationships between stimuli (e.g., same vs. different) and transcend features of the training stimuli. Recent evidence shows that learning abstract concepts is shared across a variety species including birds. Our recent work with a highly-skilled food-storing bird, Clark’s nutcracker, revealed superior same/different abstract-concept learning compared to rhesus monkeys, capuchin monkeys, and pigeons. Here we test a more social, but less reliant on food-storing, corvid species, the Black-billed magpie (Pica hudsonia). We used the same procedures and training exemplars (eight pairs of the same rule, and 56 pairs of the different rule) as were used to test the other species. Magpies (n = 10) showed a level of abstract-concept learning that was equivalent to nutcrackers and greater than the primates and pigeons tested with these same exemplars. These findings suggest that superior initial abstract-concept learning abilities may be shared across corvids generally, rather than confined to those strongly reliant on spatial memory.
Similar content being viewed by others


Introduction
The ability to learn abstract relational concepts provides a fundamental building block for many cognitive functions. In contrast to item-specific, stimulus-bound associative learning, abstract concepts transcend the specific identity of the objects involved, allowing relations among objects to be extrapolated to larger domains of novel objects (Katz, Wright, & Bodily, 2007). Research investigating nonhuman abstract-concept learning has often focused on the same/different task, a task that requires comparing the relationships between two items, rather than responding to the identity of the items. If the abstract concept of sameness is so fundamental to higher cognition, then it might be expected to be a property of cognitive systems generally, possibly with some aspects varying across species in accordance with ecological demands and brain development. We previously showed that Clark’s nutcrackers can show abstract-concept learning with a very small training set size of eight picture stimuli (Magnotti, Katz, Wright, & Kelly, 2015). In the current study, we assessed same/different abstract-concept learning in Black-billed magpies (Pica hudsonia), another corvid species. Magpies provide an important comparison to nutcrackers because magpies have a more complex social structure requiring relational processing (Dhindsa & Boag, 1992; Komers, 1989; Trost & Webb, 1997) and have a more generalized diet (Trost, 1999), relying less on cache recovery than nutcrackers (Bond, Wei, & Kamil, 2010; Lucas, Brodin, de Kort, & Clayton, 2004). Comparing magpies to nutcrackers allows us to determine if these general ecological traits support rapid abstract-concept learning.
We used the popular same/different task where subjects classify pairs of pictures as “same” or “different.” The gold standard for abstract-concept learning is transfer to novel stimuli: following learning with a stimulus set, we tested for transfer on 10 % of the trials with novel (never seen before, never repeated) stimuli (Katz & Wright, 2006). If subjects were responding on the basis of the abstract relationship (same vs. different) between stimuli that make up each pair (as opposed to specific features of the stimuli), then transfer performance to novel stimuli should be statistically equivalent to performance with trained stimuli (i.e., full concept learning).
Previous transfer tests in this same/different task with rhesus monkeys (Katz, Wright, & Bachevalier, 2002), capuchin monkeys (Wright, Rivera, Katz, & Bachevalier, 2003), and pigeons (Katz & Wright, 2006) showed no significant transfer, and hence no abstract-concept learning, following training with the initial set of these same 8 picture stimuli. Other than the previously mentioned nutcracker study (Magnotti et al., 2015), only human participants had shown transfer of same/different abstract-concept learning following training with this set of eight stimuli (Schmidtke, Magnotti, Wright, & Katz, 2013). All other species we have tested eventually show full concept learning, once they are trained with a sufficiently large number of exemplars (between 1,000 and 66,000). Here we focus on transfer performance after training with only 64 total exemplars (eight same, 56 different) as in our previous study with Clark’s nutcrackers (Magnotti et al., 2015).
Methods
Apparatus and stimuli
Magpies were tested in identical 61 cm wide × 31 cm deep × 56 cm high custom-built wooden chambers as in our previous studies with Clark’s nutcrackers (Magnotti et al., 2015; Wright, Magnotti, Katz, Leonard, & Kelly, 2016). An LCD monitor displayed the experimental stimuli (total visual angle of approximately 73° × 69°, width × height, measured from the bird’s perch); pecks to the stimuli were recorded via an infrared system. Stimuli were color travel slides (cf. Fig. 2 of Wright & Katz, 2006). Experimental events were controlled by custom software via a digital I/O panel (Phidget) connected to the food wheel and the chamber lights. A chamber light mounted centrally in the ceiling illuminated the chamber, but was extinguished during timeouts.
Procedures
All procedures were approved by the University of Manitoba’s Animal Care Committee and were in accordance with the Canadian Council on Animal Care.
Same/different training
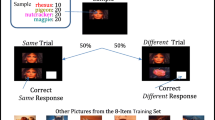
Experimental subjects were ten black-billed magpies (n = 4 females). They were trained in a two-item same/different task using the same eight stimuli (yielding eight possible same trials, 56 possible different trials), procedures, and acquisition/testing criteria previously used with Clark’s nutcrackers, rhesus and capuchin monkeys, and pigeons (Katz & Wright, 2006; Katz et al., 2002; Magnotti et al., 2015; Wright & Katz, 2006; Wright et al., 2003). Trials began with the presentation of a sample picture, to which the magpies were required to respond (peck) 20 times. At the beginning of acquisition only one response was required, but this response requirement was gradually increased to 20 as acquisition progressed. Following the sample response requirement, the comparison picture was presented beneath the sample picture along with a white rectangle to the right of the comparison picture. When the sample picture and comparison picture were the same, responses to the comparison picture were rewarded. When the sample picture and comparison picture were different, responses to the white rectangle were rewarded. Initially, a correction procedure was implemented: incorrect responses produced a 15-s blank interval and the trial was repeated until the bird made the correct choice. Only the first response for a given trial was used in the reported analyses. After the bird reached 80 % for three consecutive sessions, this correction procedure was removed, and the bird remained in acquisition until reaching 80 % accuracy on three consecutive sessions without the correction procedure (average acquisition: 35 sessions; range: 20–60 sessions). As with the other species, magpies were trained on 100-trial sessions, comprised of 50 same and 50 different trials.
Transfer testing
After fulfilling the acquisition criteria, we tested for abstract-concept learning in six transfer sessions. Transfer sessions contained 90 baseline trials and ten novel transfer trials (five same, five different). The transfer stimuli were never repeated within or across transfer sessions to ensure novelty and that item-specific learning could not contribute to transfer performance. To reduce the chance the magpies would notice outcome differences between the training and novel trials, choice responses on transfer trials were reinforced identically to those on baseline trials (Katz & Wright, 2006; Katz et al., 2002; Magnotti et al., 2015; Wright & Katz, 2006; Wright et al., 2003). As in previous studies, we used paired t-tests to look for significant differences in mean percent correct (across sessions) between baseline and transfer trials for each subject. If the t-test rejected the hypothesis of equality between the means, we next tested transfer accuracy (across sessions) against chance using a one-sample t-test to assess for partial concept learning.
Results
Transfer test
Figure 1 shows mean accuracy for each bird for baseline and transfer trials, grouped by transfer performance. Across birds, there was a significant difference between mean transfer accuracy (65.7 %) and chance performance [50 %; t(9) = 4.83, p = 0.0009]. Three magpies (BLA, RAB, BA0) showed full abstract-concept learning, that is there was no significant difference between transfer accuracy and baseline accuracy [all ts(5) ≤ 2.49; ps > 0.055]. Four magpies (RD0, RD4, BA3, YVC) showed transfer significantly below baseline accuracy, but above 50 % chance accuracy [all ts(5) ≥ 2.7, ps < 0.043], suggesting partial abstract-concept learning. In the context of a single transfer test, partial abstract-concept learning is an unclear result, as a mixture of item-specific and relational rules may control performance. Our previous work suggests that as the training set size is increased, all species eventually show full abstract-concept learning. The final three magpies (BL0, RX1, YLW) showed transfer accuracies that were not significantly different from chance accuracy [all ts(5) < 2.08, ps > 0.09].

Comparison of mean percent correct between baseline trials (gray) and transfer trials (black) for each magpie. Birds are arranged according to the results of the abstract-concept learning test. Full transfer: paired t-test between baseline and transfer showed no significant difference (p > 0.05). Partial transfer: failed full transfer, but t-test of transfer accuracy against chance (50 %) was rejected (p < 0.05). Transfer to chance: failed full and partial transfer criteria. Error bars are 1 standard error of the mean. Solid line indicates the accuracy criterion that was met before transfer (80 % correct). Dashed line indicates chance performance (50 % correct)
Control analyses
We also looked for the presence of learning or consistent biases in responding during transfer. We used a 3-factor repeated-measures analysis of variance (RM ANOVA) to assess effects of Session (1 to 6), Trial Type (same vs. different), and Stimulus Type (Baseline vs. Transfer) on accuracy across birds. We found significant main effects of Stimulus Type [mean baseline accuracy = 87 %, transfer = 66 %; F(1, 9) = 64.7, p = 10-5; η2 g = 0.23] and Trial Type [mean same accuracy = 67 %, different = 86 %; F(1, 9) = 11.4, p = 0.008; η2 g = 0.19), but no main effect of Session [F(5, 45) = 2.0, p = 0.10; η2 g = 0.02]. There was also an interaction between Trial Type and Stimulus Type, with similar accuracy for Different trials between Baseline and Transfer (mean accuracies of 85 % and 86 %, respectively), but large differences for same trials between baseline and transfer (88 % and 45 %, respectively).
As a follow-up, we used separate RM ANOVA for the three magpies that showed full transfer to test trials for a similar different-trial advantage. None of the three showed a significant effect of Stimulus Type [baseline vs. transfer; all Fs(1, 5) ≤ 6.2, ps > 0.054] or Trial Type [same vs. different; all Fs(1, 5) ≤ 3.7, ps > 0.11], but one bird (RAB) showed a significant interaction between stimulus type and trial type [F(1,5) = 7.5, p = 0.04, η2 g = 0.28], with higher performance on different trials than same trials for transfer stimuli (93 % vs. 57 %, respectively), but similar performance on different trials and same trials for baseline stimuli (86 % vs. 89 %, respectively). We note that RAB’s performance cannot be described as a total bias (i.e., choose different for all novel stimulus pairs), because performance on same trials was not below chance, nor was performance on different trials 100 %. Nonetheless, it is clear that the transfer for RAB was not as strong as was for BLA and BA0.
Although we had no a priori hypothesis about a benefit for same versus different trials during transfer, we explored the acquisition data to look for possible explanations. We compared the mean accuracy for same and different trials across the first 6 days of acquisition for the ten magpies, finding no significant performance difference [same = 44 %, different = 56 %; t(9) = -0.895, p = 0.39]. Underscoring this result, four birds showed higher same-trial performance than different-trial performance and six birds showed the opposite result. If we consider only the magnitude of the difference between same and different trials, on average birds showed a large preference for a single trial type early on in acquisition (mean absolute difference between same and different = 41 %). This difference disappeared by the end of acquisition (last three sessions of acquisition, mean absolute difference between same and different = 7 %), showing the preferences had attenuated during acquisition.
Comparison to other species’ same/different transfer results
Black-billed magpies are the second corvid species tested in our eight-item same/different abstract concept task, and like the first corvid species, nutcrackers, some magpies showed considerable transfer following training with this initial small eight-item set. We compared group mean transfer accuracy across five different species (Fig. 2) using a one-way ANOVA and found a significant species effect [F(4, 30) = 7.80, p = 0.0002, η2 g = 0.51]. Comparisons of the magpies’ transfer to the other species (unequal variance t-test; p-values shown are corrected for false discovery rate; Benjamini & Hochberg, 1995) showed similar transfer accuracy [t(16.9) = .30, p = 0.77] between magpies (group mean = 66 %) and nutcrackers (67 %), and a similar spread of transfer accuracy from chance to baseline performance. Like nutcrackers (Magnotti et al., 2015), magpies showed significantly more transfer than rhesus monkeys [mean = 51 %; t(7.7) = 2.91, p = 0.03], capuchin monkeys [mean = 52 %; t(10.6) = 3.7, p = 0.007], and pigeons [mean = 50 %; t(12.0) = 4.4, p = 0.003].

Comparison of mean accuracy on transfer trials across species: black-billed magpie (black, n = 10), Clark’s nutcracker (blue, n = 9), rhesus monkey (red, n = 4), capuchin monkey (orange, n = 3), and pigeon (green, n = 9). Error bars are 1 standard error of the mean. Solid horizontal bars are group means. Gray dashed lines indicate the acquisition criterion required before transfer testing (80 % correct) and the chance performance (50 % correct)
Discussion
Among the ten Black-billed magpies, we found a wide range of transfer to novel stimuli (from 48 % to 80 %). On average, magpies showed greater transfer than two monkey species (rhesus and capuchin) or pigeons, but were similar to Clark’s nutcrackers. How can the transfer performance of the magpies be explained? Magpies and nutcrackers share a number of relevant qualities which make them good subjects for comparison. Neither bird shows a lateralization, or bias, of visuospatial attention (Clary, Cheys, & Kelly, 2014), and both magpies and nutcrackers cache food items readily, although nutcrackers have a more specialized diet (Ducatex, Clavel, & Lefebvre, 2015) and show a greater reliance on food caches in the wild over longer periods of time than do magpies (Bond et al., 2010; Trost, 1999). However, despite this difference of diet specialization and reliance on cached food, both magpies (Zinkivskay, Nazir, & Smulders, 2009) and nutcrackers (Gould, Ort, & Kamil, 2012) show a form of what-where-when memory (relational memory) for cached food, wherein they are able to remember the locations of food cached based on its likelihood to become inedible over time. Furthermore, although a different species, azure-winged magpies (Cyanopica cyanus) and nutcrackers both show exceptional performance in an operant test of transitive inference, indicative of the ability to categorize stimuli according to relational representations (Bond et al., 2010).
Magpies and nutcrackers also differ in ways that are potentially relevant to their ability to learn abstract concepts. Magpies tend to form large flocks, with complex social hierarchies (Eden, 1987), whereas Clark’s nutcrackers most often group as mated pairs (Mewaldt, 1956). The Social Brain Hypothesis (Dunbar, 1998) suggests that social group size is the primary determinant of complex cognitive abilities shown in primate and non-primate species. This is believed to arise from the requirement to remember multiple individuals and one’s own relationship with them. Magpies rely on social interactions for flocking and processing dominance hierarchies, which requires relational processing (Dhindsa & Boag, 1992; Komers, 1989; Trost & Webb, 1997). Additionally, magpies, but not nutcrackers, are common victims of brood parasites, but have excellent ability to recognize their own eggs from the eggs of another species (Bolen, Rothstein, & Trost, 2000).
A possible perceptual explanation for novel transfer
Accurate transfer of a learned rule to novel stimuli is the gold standard of abstract-concept learning. However, perceptual-based strategies have also been proposed to account for transfer to novel stimuli. A prominent hypothesis, known as translational symmetry, suggests that transfer performance is controlled by the perceptual symmetry that exists on same trials, but is not present on different trials. When this symmetry is present, subjects respond to the lower picture, otherwise they default to the different choice. Although not directly addressed by the current data, we have previously explored this theory in detail using subjects trained in the identical procedure. Katz, Sturz, and Wright (2010) trained pigeons using the current same/different procedure and, once the subjects showed full concept learning, they were transferred to a delayed same/different task. In this task, the sample stimulus is removed following the observing response, and then comparison stimulus and white box (different choice) are presented after a variable delay (from 0 s to 6 s). If behavior was controlled by a perceptual feature, the translational symmetry hypothesis predicts that performance on same trials should decrease with increasing delay as the perceptual feature is made more difficult to extract, but performance on different trials should remain unchanged across delays—a statistical interaction between trial type and delay. Counter to this prediction, performance on same and different trials showed a similar pattern across delay. Translational-symmetry has also been experimentally rejected as an explanation for the nonhuman primate results in similar procedures (Katz et al., 2010; Wright & Katz, 2007; Wright et al., 2003). In the absence of evidence to the contrary, we suggest that the species tested here, Black-billed magpies (Pica hudsonia), learn the same/different procedure using a similar strategy to the pigeons previously tested.
Future directions
Future directions include determining how the magpies’ abstract-concept learning might change with expansion of the training stimulus set. Our expectation is that all magpies will eventually show full concept learning, as did monkeys, pigeons, and nutcrackers (Wright et al., 2016). Such a result would demonstrate qualitative similarity across these species. Also of importance are quantitative differences in transfer increases and the set size that produces full concept learning, particularly in comparison to nutcrackers because both these species come from the same branch (family Corvidae) of the evolutionary tree. A quantitative difference might reflect differences in ecological demands, evolutionary differences, and/or neural-architectural differences, whereas quantitative similarity would point to the common fundamental aspect of relational concept learning—to fully generalize a fundamental abstract concept such as same versus different.
References
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery rate: A practical and powerful approach to multiple testing. Journal of the Royal Statistical Society, Series B, 57, 289–300.
Bolen, G. M., Rothstein, S. I., & Trost, C. H. (2000). Egg recognition in yellow-billed and black-billed magpies in the absence of interspecific parasitism: Implications for parasite-host coevolution. The Condor, 102(2), 432–438.
Bond, A. B., Wei, C. A., & Kamil, A. C. (2010). Cognitive representation in transitive inference: A comparison of four corvid species. Behavioural Processes, 85(3), 283–292.
Clary, D., Cheys, A., & Kelly, D. M. (2014). Pattern of visuospatial lateralization in two corvid species, black-billed magpies and Clark’s nutcrackers. Behavioural Processes, 107, 94–98.
Dhindsa, M. S., & Boag, D. A. (1992). Patterns of nest site, territory and mate switching in Black-billed Magpies. Canadian Journal of Zoology, 70, 633–640. doi:10.1139/z92-095
Ducatex, S., Clavel, J., & Lefebvre, L. (2015). Ecological generalism and behavioural innovation in birds: Technical intelligence or the simple incorporation of new foods? Journal of Animal Ecology, 84, 79–89.
Dunbar, R. I. M. (1998). The social brain hypothesis. Evolutionary Anthropology, 6, 178–190.
Eden, S. F. (1987). The social organization of non-breeding Magpies Pica pica. Ibis, 131, 141–153.
Gould, K. L., Ort, A. J., & Kamil, A. C. (2012). Do Clark’s nutcrackers demonstrate what-where-when memory on a cache-recovery task? Animal Cognition, 15, 37–44.
Katz, J. S., Sturz, B. R., & Wright, A. A. (2010). Testing the translational-symmetry hypothesis of abstract-concept learning. Learning & Behavior, 38, 35–41.
Katz, J. S., & Wright, A. A. (2006). Same/different abstract-concept learning by pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 32(1), 80–86. doi:10.1037/0097-7403.32.1.80
Katz, J. S., Wright, A. A., & Bachevalier, J. (2002). Mechanisms of same-different abstract-concept learning by rhesus monkeys (Macaca mulatta). Journal of Experimental Psychology: Animal Behavior Processes, 28(4), 358.
Katz, J. S., Wright, A. A., & Bodily, K. D. (2007). Issues in the comparative cognition of abstract-concept learning. Comparative Cognition and Behavior Reviews, 2, 79–92. doi:10.3819/ccbr.2008.20005
Komers, P. E. (1989). Dominance relationships between juvenile and adult Black-billed Magpies. Animal Behaviour, 37, 256–265. doi:10.1016/0003-3472(89)90114-0
Lucas, J. R., Brodin, A., de Kort, S. R., & Clayton, N. S. (2004). Does hippocampal size correlate with the degree of caching specialization? Proceedings of the Royal Society, 271(1556), 2423–2429.
Magnotti, J. F., Katz, J. S., Wright, A. A., & Kelly, D. M. (2015). Superior abstract-concept learning by Clark’s nutcrackers (Nucifraga columbiana). Biology Letters, 11, 20150148. doi:10.1098/rsbl.2015.0148
Mewaldt, L. R. (1956). Nesting behaviour of the Clark nutcracker. The Condor, 58(1), 3–23.
Schmidtke, K., Magnotti, J. F., Wright, A. A., & Katz, J. S. (2013). The evolution of comparative cognition. In J. Bradley & J. Lamar (Eds.), Charles Darwin: A celebration of his life and legacy (pp. 141–156). Montgomery, AL: NewSouth Books.
Trost, C. H. (1999). In A. Poole (Ed.), Black-billed Magpie (Pica hudsonia), The Birds of North America Online. Ithaca: Cornell Lab of Ornithology. Retrieved from the Birds of North America Online: http://bna.birds.cornell.edu/bna/species/389 doi: 10.2173/bna.389
Trost, C. H., & Webb, C. L. (1997). The effect of sibling competition on the subsequent social status of juvenile North American Black-billed Magpies (Pica pica hudsonia). Acta Ornithologica, 32, 111–119.
Wright, A. A., & Katz, J. S. (2006). Mechanisms of same/different concept learning in primates and avians. Behavioural Processes, 72, 234–254.
Wright, A. A., & Katz, J. S. (2007). The generalization hypothesis of abstract-concept learning: learning strategies and related issues in Macaca mulatta, Cebus apella, and Columba livia. Journal of Comparative Psychology, 121, 387–397.
Wright, A. A., Magnotti, J. F., Katz, J. S., Leonard, K., & Kelly, D. M. (2016). Concept learning set-size functions for Clark’s nutcrackers. Journal of the Experimental Analysis of Behavior, 105, 76–84. doi:10.1002/jeab.174
Wright, A. A., Rivera, J. J., Katz, J. S., & Bachevalier, J. (2003). Abstract-concept learning and list-memory processing by capuchin and rhesus monkeys. Journal of Experimental Psychology: Animal Behavior Processes, 29(3), 184–198.
Zinkivskay, A., Nazir, F., & Smulders, T. (2009). What-Where-When memory in magpies (Pica pica). Animal Cognition, 12, 119–125.
Acknowledgments
The authors are grateful Danial Peirson, Dawson Clary, and Eric Platt for their help in data collection. This research was supported by a Natural Sciences and Engineering Research Council of Canada (NSERC) grant to DMK.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Magnotti, J.F., Wright, A.A., Leonard, K. et al. Abstract-concept learning in Black-billed magpies (Pica hudsonia). Psychon Bull Rev 24, 431–435 (2017). https://doi.org/10.3758/s13423-016-1132-4
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-016-1132-4