
Abstract
Rare or low probability targets are detected more slowly and/ or less accurately than higher probability counterparts. Various proposals have implicated perceptual and response-based processes in this deficit. Recent evidence, however, suggests that it is attentional in nature, with low probability targets requiring more attentional resources than high probability ones to detect. This difference in attentional requirements, in turn, suggests the possibility that low and high probability targets may have different susceptibilities to attention capture, which is also known to be resource-dependent. Supporting this hypothesis, we found that, once attentional resources have begun to be engaged by detection processes, low, but not high, probability targets have a reduced susceptibility to capture. Our findings speak to several issues. First, they indicate that the likelihood of attention capture occurring when a given task-relevant stimulus is being processed is dependent, to some extent, on how said stimulus is represented within mental task sets. Second, they provide added support for the idea that the behavioural deficit associated with low probability targets is attention-based. Finally, the current data point to reduced top-down biasing of target templates as a likely mechanism underlying the attentional locus of the deficit in question.
Similar content being viewed by others

Introduction
It is well-known that rare targets are detected less well than those that occur with higher probability. This deficit has been observed with a variety of experimental paradigms. In tasks like visual search, the deficit typically reveals itself in accuracy measures, with rare targets being missed more often than higher probability counterparts (Mitroff & Biggs, 2013; Wolfe et al., 2007). With paradigms like simple detection that produce high accuracy levels, the rare target effect is observed in speed of response: Rare targets are detected more slowly than more frequently occurring ones (Hon, Yap, & Jabar, 2013; Laberge & Tweedy, 1964). Here, being interested in this latter case, we exclusively utilised the simple detection paradigm, in which stimuli were presented singly and serially.
A topic of central interest relates to the cause of this target probability effect, with earlier proposals suggesting that perceptual (Dykes & Pascal, 1981; Lau & Huang, 2010; Menneer, Donnelly, Godwin, & Cave, 2010) or response processes (Fleck & Mitroff, 2007) might underlie it. However, recent work has suggested that the effect might have an attentional locus, with low probability targets appearing to require more attentional resources (than high probability ones) to detect: Relative to high probability targets, performance to low probability targets was disproportionately worsened by manipulations that reduced the amount of attentional resources available for detection and disproportionately improved by manipulations that increased the availability of such resources (Hon & Tan, 2013). The idea that the target probability effect is attention-based is, in fact, consistent with what has been found in neurophysiological investigations: The effect of target probability is most typically noted in frontoparietal brain regions associated with attention, with low probability targets eliciting greater levels of activity in these regions (Casey et al., 2001; Duncan-Johnson & Donchin, 1977; Hon, Ong, Tan, & Yang, 2012).
It is worth noting though that, in the critical experiment of that earlier study suggesting an attentional locus (Hon & Tan, 2013), the availability of attentional resources was manipulated by the imposition of an additional task to be performed concurrently with the central detection one. A concern, then, relates to how much strategies used to schedule priority of one task over the other influenced those results. A better way of investigating attentional differences between low and high probability targets might be to assess the consequences of detecting these on the processing of other attentionally-demanding events.
The idea that rare targets require greater attentional resources to detect suggests the interesting possibility that high and low probability targets might have different susceptibilities to attention capture. Capture occurs when an irrelevant stimulus draws attention to itself, away from a more central task or preoccupation. Whether or not an irrelevant stimulus can capture attention is dependent on several factors. For example, capture is most likely when the irrelevant stimulus is physically salient (Joseph & Optican, 1996; Pashler, 1988; Theeuwes, 1991), extremely surprising (Asplund, Todd, Snyder, Gilbert, & Marois, 2010; Horstmann, 2002), signals the presence of a new perceptual object (Hillstrom & Yantis, 1994; Yantis & Hillstrom, 1994), or if it enjoys some level of top-down prioritization (Folk, Remington, & Johnston, 1992; Wolfe, 1994). However, of greatest relevance to the current study, there is also evidence that capture depends on the availability of attentional resources, with it being most likely if some amount of resources remains untapped by one’s central preoccupation (Du, Yang, Yin, Zhang, & Abrams, 2013; Santangelo & Spence, 2008). For example, capture can be eliminated if spare resources are occupied with a secondary task (Boot, Brockmole, & Simons, 2005; Santangelo, Olivetti Belardinelli, & Spence, 2007) or if attentional resources are fully “frozen” within the confines of an attentional blink (Du & Abrams, 2009). Capture is, therefore, less likely in situations in which one’s central preoccupation makes high demands on attention, since, in such cases, there would be little resources left over to be captured by irrelevant stimuli.
Given the proposal that detection of low probability targets makes higher demands on attentional resources, we hypothesized that, once detection processes are underway, such targets would be more resistant to attention capture than their less attentionally-demanding high probability counterparts.
Experiment 1
As mentioned earlier, the detection of low probability targets, being more attentionally demanding, would leave fewer resources untapped than the detection of high probability targets. Importantly, this attentional difference should emerge only after detection processes have begun, since attentional resources would be deployed on the basis of some initial match between an incoming stimulus and an internally-held target template. (Detection is confirmed when sufficient evidence relating to this match is accumulated.) Accordingly, we hypothesized that low probability targets will be less susceptible to capture if the capturing stimulus appears after detection has been initiated and resource deployment has begun. Conversely, because they would leave more resources unconsumed, high probability targets would be much more susceptible to capture under these same circumstances.
To test this hypothesis, we presented our capturing stimulus shortly after the onset of the target. This short delay would allow for preliminary matches between the incoming target stimulus and the internal target template to occur, triggering the deployment of resources, and allowing for the attentional differences between targets described above to manifest.
Methods
Participants
28 participants from the National University of Singapore participated in this study. All participants had normal or corrected-to-normal vision.
Stimuli
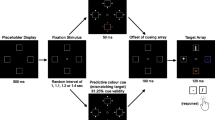
Letter stimuli were used in this study, with these being presented in black against a white background. The letters were presented in Courier New font, which, when viewed from a distance of 50 cm, subtended approximately 1.4° of visual angle both vertically and horizontally. All stimuli were presented in the centre of the screen. The capturing stimulus was a red dot that appeared either to the left of, right of, above or below the letter. The dot occurred equally often across the 4 possible locations. The dot measured .58° of visual angle vertically and horizontally, and appeared at a centre-to-centre distance of 1.60° of visual angle from the letter.
Procedure
In the experiment proper, participants observed a single 240-trial block of serially-presented letter stimuli, with the objective of detecting occurrences of two letters that were designated as targets. The targets were always the letters “C” and “W. Together, the two targets accounted for 50 % of all trials within the block. Critically, though, one target letter was presented on 10 % of all trials within the block (low probability target), while the other was presented on 40 % (high probability target). Letter-probability assignment was counterbalanced across the two letters used as the targets, with preliminary analysis confirming that this did not interact with the main capture stimulus and target probability variables. Distractors accounted for the remaining 50 % of trials in a block, with the distractor set comprising all letters of the alphabet not designated as targets. Regardless of whether a letter was a target or distractor, it was presented for 1000 ms, followed by a blank frame presented for 1000 ms and, subsequently, the presentation of the next stimulus. To indicate detection of a target, participants pressed the “1” button with the index finger of their master hand, with this same response being made to both targets. This minimised the likelihood that any effects of probability would be due to differences in response processes. Trial order was randomised for each participant.
On a small number of trials, a capturing stimulus (a red dot) was presented 30 ms after the onset of the letter stimulus, remaining onscreen for 50 ms before being extinguished. The capturing stimulus was presented 8 times each (twice in each of the four positions mentioned above) on low probability target, high probability target and distractor trials. Participants were not informed of the occurrence of the capturing stimulus beforehand.
The experiment proper was preceded by a 100-trial long practice that utilised the same target identities and stimulus probabilities as the experiment. The practice was performed to familiarize participants with the paradigm and, in particular, with the probabilities associated with the specific targets. No capturing stimuli were presented during the practice.
The stimuli were presented on a 24-in LCD monitor, with the experiment being implemented on a PC running the E-Prime software.
Results
Given our use of high visibility stimuli that were presented serially and at a slow pace, detection accuracy was unsurprisingly high (Table 1). The number of false alarms to distractor trials on which the dot appeared was exceedingly few (M = .25) and, in fact, less than that to distractor trials on which the dot was absent (M = .71), t(27) = 2.79, p = .01. This suggests that the appearance of the dot per se did not dictate responding.
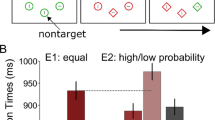
Our main interest was in response times (RT), which we now turn to. These data are depicted in Fig. 1. A fully-within probability (high, low) x capture stimulus (present, absent) ANOVA revealed significant main effects of probability [F(1, 27) = 38.25, p < .001] and capture stimulus [F(1, 27) = 27.26, p< .001], as well as the interaction of the two [F(1, 27) = 12.09, p = .001]. Simple effects tests revealed that the appearance of the capturing stimulus affected performance to high [t(27) = 5.72, p < .001] but not low probability targets [t(27) = 1.03, p = .310]. These data support the hypothesis that, once detection processes have begun, the greater consumption of attentional resources by low probability targets reduce their susceptibility to attention capture.Footnote 1 And this is so even when the capturing stimulus is a putatively powerful one, as it was here (a sudden-onset, colour singleton that appeared on a very small number of trials).

Response times to low and high probability targets in Experiment 1 as a function of whether a delayed-onset capturing stimulus (dot) was presented or not. Error bars indicate 1 SEM
Experiment 2
In Experiment 1, we found that, once detection processes have started, low probability targets are largely resistant to attention capture. In Experiment 2, we examined what would happen when the capturing stimulus is presented simultaneously with the target. Under such conditions, it is unlikely that detection processes will be triggered before apprehension of the capturing stimulus, particularly given its greater salience relative to the letter stimuli. Prior to the initiation of detection processes, attentional resources would be “free-floating” and available to be captured. Accordingly, we predicted that, here, both low and high probability target detection would be interfered with by the appearance of the capturing stimulus. Such a finding would support the proposal that the resistance to capture observed in Experiment 1 reflects differences in the amount of attentional resources specifically required for detection by low and high probability targets.
Methods
Participants
31 participants from the National University of Singapore participated in this study. All participants had normal or corrected-to-normal vision.
Stimuli and Procedure
With the exception of the onset of the capturing stimulus, which now coincided with the onset of the letter stimulus, all other aspects of the experiment were identical to Experiment 1.
Results
The accuracy data from this experiment are presented in Table 2. As before, we found that the number of false alarms to distractor trials on which the dot appeared (M = .29) was less than that to distractor trials on which the dot was absent (M = 1.35), t(30) = 2.77, p = .01. Again, this indicates that participants did not use the appearance of the capturing stimulus as a cue for responding.
Figure 2 depicts the RT data from this experiment. A fully-within probability (high, low) x capture stimulus (present, absent) ANOVA performed on these data revealed significant main effects of probability [F(1, 30) = 45.67, p = .001,] and capture stimulus [F(1, 30) = 19.77, p < .001]. Critically, though, the interaction between the two was not significant [F < 1, n.s.]. Simple effects tests confirmed that the appearance of a capturing stimulus adversely affected RTs to both low [t(30) = 2.55, p = .015] and high probability targets [t(30) = 3.28, p = .003]. These results support the idea that both low and high probability targets are susceptible to attention capture if conditions allow for the irrelevant stimulus to be apprehended before detection processes can be initiated, when attentional resources are still un-deployed.

Response times to low and high probability targets in Experiment 2 as a function of whether a simultaneous-onset capturing stimulus (dot) was presented or not. Error bars indicate 1 SEM
In this experiment, the appearance of the capturing stimulus had an equivalent effect on both low and high probability targets. This is different to what was found in Experiment 1, in which the capturing stimulus affected high, but not low, probability targets. To test this pattern formally, we compared the difference in size of the capture effects between high and low probability targets (i.e., [High ProbabilityCaptureStimulusPresent - High ProbabilityCaptureStimulusAbsent] - [Low ProbabilityCaptureStimulusPresent - Low ProbabilityCaptureStimulusAbsent]) from the two experiments (Fig. 3). A large positive score (as in the case of the Experiment 1 bar in Fig. 3) indicates that the capturing stimulus had a larger effect on high probability targets than on low probability ones.Footnote 2 A score not significantly different from zero (as in the case of the Experiment 2 bar) indicates that the appearance of the capturing stimulus affected high and low probability targets to a similar extent.Footnote 3 A one-way ANOVA performed on these data confirmed that the two experiments produced different patterns of results, F(1, 57) = 4.24, p = .044.

Difference between high and low probability target capture effects (see Main Text) from the two experiments. Error bars indicate within-group SEMs
Discussion
Our findings speak to two issues. First, they provide added support to the idea that the target probability effect is attentional in nature; in particular, that low probability targets require more attentional resources than high probability ones to detect (Hon & Tan, 2013). A consequence of this attentional difference is that low probability targets are less susceptible to capture once detection processes have begun and attentional resources are deployed (Experiment 1). This likely reflects the fact that detection of low probability targets, being more attentionally demanding, leaves fewer untapped resources available to be captured by the irrelevant stimuli. This interpretation is buttressed by the finding that capture was observed on both low and high probability target trials when the capturing stimulus appeared before detection could begin, when resources were still “free-floating” (Experiment 2).
Second, our data suggest that whether or not capture occurs is determined, in part, by how targets are represented within higher-order task sets. As mentioned earlier, other studies have demonstrated failures of capture when untapped resources are limited or depleted in some way. A typical way by which this is accomplished is through the manipulation of task conditions (e.g., depleting spare resources via the imposition of a secondary task). Here, however, we demonstrate a reduced susceptibility to capture specific to a particular stimulus, even when the task conditions were held constant for our task relevant stimuli (i.e., targets). This suggests the more general theoretical point that a given task-relevant stimulus can be rendered resistant to capture simply by virtue of the way it is represented within the mental task sets that guide behaviour. Here, our low and high probability targets were equivalent in the sense that they were equally relevant to the task (the objective was to detect all occurrences of both targets), shared the same response and were even presented within the same block of trials. Nonetheless, the representations of the two within the task set would have differed in terms of what was encoded with respect to probability of occurrence. We propose that this representational difference, and its attentional consequences, is what grants low probability targets their resistance to capture once detection is underway.
Taken together, the points above hint at the specific manner in which attentional differences between low and high probability targets arise. As mentioned previously, detection of a target is accomplished when an incoming stimulus matches an internally-held template of the target. This target template is instantiated and maintained in the appropriate peripheral (e.g., perceptual) system through the action of top-down (biasing) signals from more central systems that encode the “task set” or higher-order mental representation encompassing the various task-relevant elements (Corbetta & Shulman, 2002; Kastner & Ungerleider, 2000). The fact that attentional differences between low and high probability targets become apparent only after detection processing has begun suggests that something makes stimulus-template matching for low probability targets more attentionally demanding. We propose that this occurs because, as one learns that a given target only rarely appears, adjustments are made to this target’s representation within higher-order tasks sets.Footnote 4 The effect of which is that a reduced level of top-down biasing is sent to the target template coding for this specific target; that is, the template is maintained at a lower level of “readiness”. This reduced level of readiness results in an increased amount of attentional resources being required for determination of matches between an incoming stimulus and the target template (e.g., greater attentional guidance may be necessary for the accumulation of evidence that determines a match), the behavioural consequence of which is that it would take longer and be more effortful to detect low probability targets. It is also worth noting that the preceding framework allows for a simple explanation of why perceptual manipulations have been found to affect low probability targets more than high. Targets whose templates are maintained at an impoverished level (as with low probability targets) would likely be affected more by manipulations that limit visibility or discriminability, since these would compound the original disadvantage produced by reduced levels of top-down biasing.
We can rule out several alternative explanations for our findings. Our results are unlikely to have been caused by differences in perceptual or cognitive load, which have been proposed to be critical in determining capture – capture is most likely when perceptual load is low or when cognitive load is high (Lavie, 2005). In our experiments, perceptual and cognitive loads were the same for both probability targets; however, failure to capture was only found with low probability targets once detection has begun. Finally, the failure to capture in Experiment 1 is unlikely to have been due to a floor effect associated with the low probability targets. The standard deviation for the capture stimulus-absent low probability targets (62 ms) was larger than the effect of capture in that experiment (34 ms), indicating that there was ample room for low probability target performance to vary.
In conclusion, although low probability targets are generally associated with a performance deficit, here, we demonstrate that the mechanisms that underlie this deficit may produce an unexpected benefit for such targets. Specifically, the attentional mechanisms underpinning that deficit may grant low probability targets some level of resistance to attention capture, at least once those mechanisms have begun to operate.
Notes
We replicated this pattern of results in another experiment in which the delayed-onset capturing stimulus was a red box that surrounded the letter stimulus (as opposed to the red dot used in Experiment 1 above). As in Experiment 1, we found, with the box version, that probability (high, low) interacted with capture stimulus (present, absent), F(1, 29) = 5.69, p = .024. Simple effects tests confirmed that the appearance of the box adversely affected performance to high [t(29) = 5.27, p < .001] but not low probability targets [t(29) = 1.53, p = .14]. Furthermore, a cross-experiment analysis revealed that there was no difference in the pattern of capture effects found in Experiment 1 and the box version (F < 1, n.s.).
The difference score indicated by the Experiment 1 bar is significantly greater than zero (p = .001, one-sample t-test).
The difference score indicated by the Experiment 2 bar is not significantly different from zero (t < 1, n.s.).
At this time, the exact nature of these adjustments and why they occur is still unclear. Obviously, these issues will benefit from further focused inquiry.
References
Asplund, C. L., Todd, J. J., Snyder, A. P., Gilbert, C. M., & Marois, R. (2010). Surprise-induced blindness: A stimulus-driven attentional limit to conscious perception. Journal of Experimental Psychology: Human Perception & Performance, 36(6), 1372–1381.
Boot, W. R., Brockmole, J. R., & Simons, D. J. (2005). Attention capture is modulated in dual-task situations. Psychonomic Bulletin & Review, 12(4), 662–668.
Casey, B.J., Forman, S.D., Franzen, P., Berkowitz, A., Braver, T.S., Nystrom, L.E., Thomas, K.M. & Noll, D.C. (2001). Sensitivity of prefrontal cortex to changes in target probability: A functional MRI study. Human Brain Mapping, 13(1), 26–33.
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed and stimulus-driven attention in the brain. Nature Reviews Neuroscience, 3(3), 201–215.
Du, F., & Abrams, R. A. (2009). Onset capture requires attention. Psychonomic Bulletin & Review, 16(3), 537–541.
Du, F., Yang, J., Yin, Y., Zhang, K., & Abrams, R. A. (2013). On the automaticity of contingent capture: Disruption caused by the attentional blink. Psychonomic Bulletin & Review, 20(5), 944–950.
Duncan-Johnson, C. C., & Donchin, E. (1977). On quantifying surprise: The variation of event-related potentials with subjective probability. Psychophysiology, 14(5), 456–467.
Dykes, J. R., & Pascal, V. (1981). The effects of stimulus probability on the perceptual processing of letters. Journal of Experimental Psychology: Human Perception & Performance, 7, 528–537.
Fleck, M. S., & Mitroff, S. R. (2007). Rare targets are rarely missed in correctable search. Psychological Science, 18(11), 943–947.
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception & Performance, 18(4), 1030–1044.
Hillstrom, A. P., & Yantis, S. (1994). Visual motion and attentional capture. Perception & Psychophysics, 55(4), 399–411.
Hon, N., Ong, J., Tan, R., & Yang, T. H. (2012). Different types of target probability have different prefrontal consequences. NeuroImage, 59, 655–662.
Hon, N., & Tan, C. H. (2013). Why rare targets are slow: Evidence that the target probability effect has an attentional locus. Attention, Perception, & Psychophysics, 75(3), 388–393.
Hon, N., Yap, M. J., & Jabar, S. B. (2013). The trajectory of the target probability effect. Attention, Perception, & Psychophysics, 75, 661–666.
Horstmann, G. (2002). Evidence for attentional capture by a surprising color singleton in visual search. Psychological Science, 13(6), 499–505.
Joseph, J. S., & Optican, L. M. (1996). Involuntary attentional shifts due to orientation differences. Perception & Psychophysics, 58(5), 651–665.
Kastner, S., & Ungerleider, L. G. (2000). Mechanisms of visual attention in the human cortex. Annual Review of Neuroscience, 23, 315–341.
Laberge, D., & Tweedy, J. R. (1964). Presentation Probability and Choice Time. Journal of Experimental Psychology, 68, 477–481.
Lau, J. S., & Huang, L. (2010). The prevalence effect is determined by past experience, not future prospects. Vision Research, 50(15), 1469–1474.
Lavie, N. (2005). Distracted and confused?: Selective attention under load. Trends in Cognitive Sciences, 9, 75–82.
Menneer, T., Donnelly, N., Godwin, H. J., & Cave, K. R. (2010). High or low target prevalence increases the dual-target cost in visual search. Journal of Experimental Psychology: Applied, 16(2), 133–144.
Mitroff, S. R., & Biggs, A. T. (2013). The ultra-rare-item effect: Visual search for exceedingly rare items is highly susceptible to error. Psychological Science, 25(1), 284–289.
Pashler, H. (1988). Cross-dimensional interaction and texture segregation. Perception & Psychophysics, 43(4), 307–318.
Santangelo, V., Olivetti Belardinelli, M., & Spence, C. (2007). The suppression of reflexive visual and auditory orienting when attention is otherwise engaged. Journal of Experimental Psychology: Human Perception & Performance, 33(1), 137–148.
Santangelo, V., & Spence, C. (2008). Is the exogenous orienting of spatial attention truly automatic? Evidence from unimodal and multisensory studies. Consciousness & Cognition, 17(3), 989–1015.
Theeuwes, J. (1991). Cross-dimensional perceptual selectivity. Perception & Psychophysics, 50(2), 184–193.
Wolfe, J. M. (1994). Guided Search 2.0 A revised model of visual search. Psychonomic Bulletin & Review, 1(2), 202–238.
Wolfe, J. M., Horowitz, T. S., Van Wert, M. J., Kenner, N. M., Place, S. S., & Kibbi, N. (2007). Low target prevalence is a stubborn source of errors in visual search tasks. Journal of Experimental Psychology: General, 136(4), 623–638.
Yantis, S., & Hillstrom, A. P. (1994). Stimulus-driven attentional capture: Evidence from equiluminant visual objects. Journal of Experimental Psychology: Human Perception & Performance, 20(1), 95–107.
Acknowledgments
This study was supported by grant FY2015-FRC-1-006 from the National University of Singapore.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Hon, N., Ng, G. & Chan, G. Rare targets are less susceptible to attention capture once detection has begun. Psychon Bull Rev 23, 445–450 (2016). https://doi.org/10.3758/s13423-015-0921-5
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-015-0921-5