
Abstract
Fish learning and cognition are usually approached by testing single individuals in various devices such as mazes that have serious drawbacks, especially in gregarious species, including the stress induced by the test procedure. This might impair the results and lead to misinterpretation about the learning abilities of the targeted species. In order to provide an alternative to the individual-based tests, we investigated for the first time the operant conditioning of four similar groups (50 individuals per tank) of sea bass. We used two computerized self-feeder devices per tank, each coupled with individual electronic identification and that were alternately activated during varying positive appetitive reinforcement period of time (7 to 1 day). Learning abilities were examined at both group and individual levels. At the group level, the operant conditioning was demonstrated as the triggering activity significantly decreased when the device was turned off and increased when it was turned on, whatever the reinforcement period duration. The individual level analysis revealed a more complex situation with fish showing different learning performances that can be best explained through the producer-scrounger game theory.
Similar content being viewed by others

Most of the learning experiments are still based on mammal and bird models (Shettleworth, 2009), but during the past decade an increasing number of studies have used different fish species to study cognition, which includes perception, attention, memory formation, and executive functions related to information processing such as learning and problem solving. Indeed, fishes like the rest of vertebrates exhibit a rich array of sophisticated behaviours and learning plays a pivotal role in their behavioural development (Brown, Laland, & Krause, 2007).
Among other forms of learning, operant conditioning techniques are those in which a consequence is contingent upon the behaviour of the organism. This is carried out with either positive reinforcement techniques, in which the animal must make a choice or response in order to gain an appetitive stimulus, or negative reinforcement techniques, in which the animal must produce a response in order to avoid an aversive stimulus (Blaser & Vira, 2014). Appetitive reinforcers include food (Bilotta, Risner, Davis, & Haggbloom, 2005; Colwill, Raymond, Ferreira, & Escudero, 2005; Grossman et al., 2011; Mueller & Neuhauss, 2012; Parker et al., 2012; Sison & Gerlai, 2010; Williams, White, & Messer, 2002) and social stimuli (Al-Imari & Gerlai, 2008; Karnik & Gerlai, 2012; Pather & Gerlai, 2009). Operant conditioning is usually investigated on single individuals placed in diverse devices such as shuttle box, T maze, or plus maze, using either simple response discrimination (right vs left) or a visual cue (e.g. red card) marking the correct choice (Blaser & Vira, 2014).
All these procedures have serious drawbacks that can impair fish behaviour and especially its learning abilities. For example, food reinforcers for fish are often administered by hand, resulting in variability of quantity and timing in addition to requiring the constant attention of an experimenter which reduces efficiency (Blaser & Vira, 2014). The main concern is, however, that learning is likely to be impaired by acute stress responses induced by experimental conditions. When placed in a maze, the fish will be isolated and often handled between each trial as shown by previous studies on Zebrafish (Danio rerio) (Blaser & Vira, 2014) and sea bass (Dicentrarchus labrax) (Benhaïm, Bégout, Lucas, & Chatain, 2013; Benhaïm et al., 2013). Even though a plus or T maze can provide an objective measure of both acquisition (number of errors, latency to choose) and response to test (preference test, extinction) that can be observed with video tracking, there is, however, so far very little standardization in the way behaviours are measured or reported and this results in considerable variability in the robustness of effects. Measured behaviours include latency to choose, proportion of correct choice, preference for the reinforcer (e.g. duration of time spent near shoal), preference for the discriminative stimulus (e.g. duration of time spent in blue arm). Acquisition measures, including probability of correct choices and latency to choose during training trials, were reported in only a few instances (Bilotta et al., 2005; Colwill et al., 2005; Parker et al., 2012; Williams et al., 2002) with the more common measure being response to the discriminative stimulus (i.e. the correct arm) in a single probe test. Appetitive operant techniques reported so far require considerably more investigation before they will be amenable to large-scale screening endeavours. As suggested by Blaser and Vira (2014), the use of maze-type techniques in which experimenter intervention is required between each trial is far less efficient than automated free-operant techniques, especially when large numbers of trials are required.
Previous studies have shown that fish employ multiple spatial strategies that closely parallel those described in mammals and birds (Lopez, Broglio, Rodriguez, Thimus-Blanc, & Salas, 1999). Animals can solve a spatial task by learning an association between a response and a reward (response strategy) or they can learn an association between a given place and a reward (place strategy) (Dudchenko, 2001; Gibson & Shettleworth, 2005). Recently, a couple of studies on European sea bass (Dicentrarchus labrax) under maze conditions provided a first insight into the ability of this species to learn to discriminate between two 2-D objects associated with a simple spatial task that enabled the tested individual to visually interact with an unfamiliar congener (the reward) located behind a transparent wall at the end of one of the two arms of a maze (Benhaïm, Bégout, Lucas, & Chatain, 2013; Benhaïm et al., 2013). These studies concluded that it would be useful to design experiments minimizing the stress and increasing the number of learning sessions and/or individuals.
Feeding behaviour is a complex and flexible behaviour that encompasses several behavioural responses associated with eating, including modes of feeding and feeding habits, mechanisms of food detection, frequency of feeding, and food preferences (Volkoff & Peter, 2006). Feeding behaviour of fish can be modified by both Pavlovian and operant learning (to catch or manipulate the food; Warburton, 2007). For obvious reasons, feeding motivation and its links with growth variation has been extensively studied in cultured fish species (Martins, Schrama, & Verreth, 2005).
Self-feeders are a good example of operant learning in fish with direct applications for aquaculture. These devices deliver food on demand by employing the learning ability of fish. They have been particularly successful in European sea bass, a leading species of Mediterranean aquaculture that was domesticated in the 1980s. This species demonstrated a great ability to use such a system and a high plasticity when facing the triggering device (pushing, pulling, biting) (Covès et al., 2006). Experiments with a self-feeder coupled with a PIT-tag detection antenna, gives the possibility to reveal individual and group demand of feeding. Such combination of techniques was successfully developed in sea bass (Covès et al., 2006; Covès, Gasset, Lemarié, & Dutto, 1998; Rubio et al., 2004). In this species, an intriguing individual specialization exists when using self-feeders with three triggering categories coexisting: high triggering (HT), low triggering (LT), and zero-triggering (ZT) fish (Covès et al., 2006; Di-Poï et al., 2007; Millot & Bégout, 2009; Millot et al., 2008). In small populations composed of 50 to 100 fish, whatever the experimental conditions, the same pattern is always observed: HT fish are very few, with only one or two animals being responsible for most of the triggering activity under a reward regime of one or two pellets per individual given after each actuation (Covès et al. 2006). On the other hand, the ZT status would be attributed to fish that never actuate the device, while the rest of the population is being composed of individuals that seldom actuated the trigger (LT) (Covès et al. 2006; Millot et al. 2008). The percentage of individuals in LT and ZT categories is extremely variable according to authors, but the proportion of HT remains the same (Ferrari, Benhaïm, Colchen, Chatain, & Bégout, 2014). It is important to note that when an individual actuates the trigger, it delivers food for all the fish in the tank. In most cases, the three categories exhibit no difference in mean initial or final weights or in mean specific growth rate (Covès et al., 2006; Di-Poï et al., 2007; Di-Poï et al., 2008); that is, HT does not take advantage of its status. This social structure is better understood in light of recent studies showing that the fish triggering activity is linked to personality traits (Benhaïm et al., 2012; Ferrari, Benhaïm, et al., 2014). As already mentioned by Di-Poï et al. (2008), the observed social pattern could be based on the producer-scrounger model firstly described by Barnard and Sibly (1981). HT may play the role of the producers that feed the entire group, whereas other fish (especially ZT) may play the role of scroungers, individuals that parasitically exploit the food made available by the producers.
Here we investigated for the first time the operant conditioning flexibility of several groups of sea bass using two self-feeder devices that were alternately activated, and we tested the effect of reducing the positive appetitive reinforcement duration. The aim of this study was to propose a new approach to investigate fish learning in groups as opposed to experiments performed in isolated conditions. This approach dramatically changes the way learning experiments are conducted in fish. It avoids the main drawbacks usually met in isolation situations and provides the fish with more realistic/natural conditions. Finally, it provides the opportunity to study potential links between social structure and foraging strategies.
In the current experiment, fish were tested under group conditions, but with a design allowing them to be tracked individually without stress induced by handling and without any limitation in the number of learning trials and sessions. This design ensured a consistent level of motivation over time. We specifically addressed the following questions:
-
On the group scale, can fish learn to switch from an inactive device to an active one?
-
Does a decrease of the positive reinforcement duration alter the learning process?
-
How does the individual level trigger the group learning response, and how does it impact the individual specialization based on the triggering activity?
Material and method
Fish and experimental conditions
Fish were hatched and reared at the experimental research station of Ifremer (Palavas-les-Flots, France) according to sea bass rearing standards (Chatain, 1994). A sample of 600 fish was transported at 86 days post hatching (dph) to the Fish Ecophysiology Platform of La Rochelle (PEP; , France—http://wwz.ifremer.fr/pep). A subsample of 200 fish was then distributed in four 400 L tanks (T1 to T4) located in a dedicated room. At 257 dph, the fish, weighing 15.32 ± 4.96 g, were tagged with a conventional PIT tag to monitor each fish individually using a self-feeder equipped with PIT-tag detection antenna. The four 400 L tanks were supplied with sand-filtered seawater in a recirculated system (flow rate of 4 m3 h−1 in each tank, and 20% water renewal per day). Tanks were surrounded by an opaque black curtain to avoid any disturbance to the fish. A white light (Philips, 80 W) was suspended above each tank. The light cycle was controlled (14 hours day/10 hours night) throughout the experiment. The physicochemical properties of the water were monitored daily to guarantee optimum conditions. Water temperature was maintained at 21.4 ± 1.3 °C, O2 saturation at 72.4 ± 11.2% and salinity at 26.4 ± 1.2. Ammonia, nitrite, and nitrate concentrations were 0.05 ± 0.05, 0.11 ± 0.06, and 0.37 ± 0.11 mg L -1, respectively. Fish were fed with commercial food (Le Gouessant®, NEO START-coul4, 4 mm diameter, 52% proteins, France) using self-feeders.
Evaluation of biological performances
Growth was measured at the beginning and the end of the experiment, and no intermediate biometrics measurements were done to avoid human disturbances on feed demand structure and associated learning processes. The variable chosen to evaluate the biological performances were the following: specific growth rate (SGR; (% of mass per day) = 100 (Ln BMf - Ln BMi) / t), with BMf and BMi being the final and initial body mass (g) respectively, and t the total number of days; feed demand (FD; g kg−1 of fish biomass).
Feed demand behaviour and self-feeder apparatus
The device to operate the feeder comprised a screened type sensor (a metal rod protected in a PVC cylinder surrounded by the tag detection antenna (Covès et al. 2006) and a control box linked to a computer (see Fig. 1; Imetronic, Pessac, France). After each actuation, fish were rewarded with pellets (at least one per fish), and feed dispensers were regulated to distribute always the same quantity of food, which corresponded to a mean of 1.75 ± 0.19 g. The reward level was a compromise between minimizing wastage and optimizing feed allocation to the group. Such a set up allowed us to monitor on a daily basis the individual feed demand behaviour. Triggering activity recordings were done continuously. Feed demand behaviour was followed over 105 days from 500 dph to 605 dph.

Demand feeding system; fish activated the food dispenser, pushing the trigger located inside the PVC cylinder surrounded by a PIT tag detection antenna connected to a computer, allowing registration of the PIT tag number of the individual as well as the date and time of actuation (after Covès et al., 2006)
Characterization of triggering categories
Fish were characterized according to their triggering activity. They were classified into three categories, taking into account their individual mean contribution to the total number of trigger actuations within the tank (%) for each self-feeder device (SF1 and SF2) and each situation (Code: On and Off) over the whole duration of the experiment (105 days) following Ferrari, Benhaïm, Colchen, et al. (2014): high triggering HT (≥8% of total actuations), low triggering LT (<8%), and zero triggering ZT (<2%).
Learning process
Once the social structure based on self-feeding activity was well established (regular feed demand activity), the first feeder (SF1) was switched off and a second feeder (SF2), placed in each tank at the opposite side of SF1, was immediately activated (see Fig. 2). The activation of SF2 corresponded to the beginning of the experiment (500 dph). For each day, the triggering activity was recorded, and the quantity of food distributed by each self-feeder and in each tank was calculated. For each tank, the success on the active feeder was assessed as the number of trigger actuations on the active feeder * 100 / total number of actuations on both active and inactive feeders, in percentages. The experiment was carried out from 500 dph to 605 dph and was structured in 10 successive phases, each of them corresponding to a learning phase (time to reach 90% of success on the active feeder at the tank level), followed by a reinforcement phase of decreasing duration (see Table 1). For example, in Phase 2 (Table 1), SF2 was turned off after the fish in the tank performed at least 90% of success, then SF1 was activated until the fish reached at least 90% and during an additional 7-day reinforcement phase.

Computerized self-feeder device used in this experiment in one of the four tanks. SF1 and SF2 First and second self-feeder, the triggers being inside both PVC cylinders. A PIT tag detection antenna. D1 and D2 Feed dispensers
At the end of experiment (605 dph), all the fish were killed with an anaesthetic overdose and their sex determined.
Data analyses
All variables were checked for normality and homoscedasticity.
Modelling process
All mixed models presented below were performed in R Version 3.2.2 (R Core Team, 2015) as follows. A selection based on AICs (Akaike information criterion) was applied to determine whether the random effects affected the model intercept only or both model intercept and slopes. Finally, the significance of the random and fixed effects at a 5% alpha risk threshold was tested by likelihood ratio test (LRT) between nested models respecting marginality of the effects that are supposed to follow a \( {\chi}^2 \) distribution under the null hypothesis (Type II tests; Fox & Weisberg, 2011). Diagnostics based on residuals were performed to assess the adequacy of the model and compliance to the underlying assumptions. Then analyses were followed by post hoc multiple comparison tests (Hothorn, Bretz, & Westfall, 2008).
Biological performances
FD was compared between tanks using a three-way ANOVA with self (SF1 and SF2) and sex as fixed factors and tank (four tanks) as a random factor.
A chi-square test was used to compare the sex ratio between tanks. SGR was analysed using a generalized linear mixed-effects model (GLMM), with individual triggering activity as an explanatory variable and sex (fixed) and tank (random) as categorical predictors.
Social structure
Individual triggering activity (mean contribution) was analysed using a GLMM with gamma error distribution and inverse link function. Two nested random effects (individuals nested within tanks) were used to account for variability due to experimental tanks and individuals. The fixed part of the model included Code (Off, On) and Self (SF1, SF2) effects to assess for individual triggering contribution on both self-feeders.
Chi-square tests were used to compare the proportion of individuals according to the triggering category (HT, LT, or ZT) in each tank for each situation (SF1 On, SF1 Off, SF2 On, SF2 Off).
Contribution differences between triggering categories according to the self whether active or inactive were analysed using a GLMM with gamma error distribution and inverse link function. The fixed part of the model included Code (Off, On), Self (SF1, SF2) and Triggering category (HT, LT, ZT) effects, and Tank was used as a random factor.
Learning performances
At the group level, the learning durations (i.e. time to reach 90% of success on the active feeder, in days) were compared using a Kruskal–Wallis test with phase (Phases 7, 5, 3 and 1), Self (SF1 and SF2), or Tank (four tanks) as factors. Significant tests were followed by a rank-based multiple comparisons (Zar, 1984). Individual contributions were calculated for each day, and mean contribution per phase were also calculated on both active and inactive feeders.
At the individual level, learning performances on both self-feeders in both situations (when inactive and when active) were assessed for each tank, comparing the triggering activity between the beginning and the end of each learning phase. To improve the models robustness, only individuals with a significant triggering contribution were included (see previous social structure section). The built data set was analysed using a GLMM, with Poisson error distribution and log link function. In this analysis, the fixed part of the model included a Period_Self (Start-SF1, Start-SF2, End-SF1, End-SF2) and a Code (Off, On) effect to assess for individual learning performances on both self-feeders (an increase or a decrease of the trigger actuations number were expected depending on whether self-feeders were active or inactive).
For all tests, the significant threshold was p < .05 unless otherwise stated. GLMMs were fitted using the lme4 package (Bates, Mächler, Bolker, & Walker, 2014).
Results
Biological performances
No fish died over the experiment duration. Initial mean body mass was 96.4 ± 32.0 g at 488 dph; N = 200 and final body mass was 164.5 ± 64.2 g at 605 dph. There was a significant interaction between Self and Tank for FD, F(3, 361) = 2,7, p = .04. Further Newman-Keuls tests showed that FD in tank 4/SF2 was significantly higher compared to all other conditions (p < .01 in all cases). No sex effect was recorded for FD.
The difference in sex ratio between the tanks was significant (72% to 75% of males in Tanks 1, 2 and 3 but only 54% in Tank 4, χ2 = 13.7, df = 3, p < .05). SGR during the experiment duration was 0.43 ± 0.17% day-1. SGR was significantly different between Sex and Tanks (LRT, df = 1, χ2 = 12.2, p < .001; LRT, df = 1, χ2 = 226.5, p < .001, respectively). SGR was not explained by individual contributions (LRT, df = 1, χ2 = 0.43, p = .5). The following model was used: lmm(SGR~Sex + (1|Tank). Post hoc tests showed that SGR was significantly higher in females (z = -3.6, p < .001).
Social structure
The proportion of individuals according to the triggering category was not significantly different between tanks in each situation (SF1 On, SF1 Off, SF2 On, SF2 Off). Table 2 reports the global proportion of individuals in each triggering category. The four tanks mean proportions were 4.7±1.8% of HT, 8.6 ± 4.9% of LT, 86.7 ± 3.7% of ZT.
At the individual level, GLMM showed a significant interaction between Self and Code (LRT, df = 1, χ2 = 4.5, p = .03) and a significant effect of Individuals (LRT, df = 3, χ2 = 2492.5, p < .001), then the following model was used to analyse the contribution:
glmer(Contribution + 1~Self + (Self|ID), family = Gamma(link = "inverse")) with Contribution (Individual mean triggering activity in %), Self (SF1, SF2), and ID (Individual identity). Fifteen individuals showed significantly higher contributions compared to all other individuals. These individuals (reported in Table 3) were later used to assess for learning performances.
There was a significant interaction for percentage of triggering activity between Triggering category (HT, LT, ZT) and Self (SF1, SF2) (LRT, df = 5, χ2 = 1403.1, p < .001) and a Tank significant effect (LRT, df = 1, χ2 = 11.4, p < .001). The following model was used:
glmm(Contribution + 1~Self * Category + (1|Tank), family = Gamma(link = "inverse")). Post hoc tests showed that HT contributions did not differ between self-feeders (z = 2.3, p = .18) but LT and ZT contributions on SF2 were higher than on SF1 (z = -4.9, p < .001; z = -2.8, p = .04, respectively).
Learning process at the group level
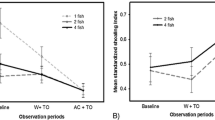
The criteria of 90% of success was obtained in all tanks on both self-feeders when they were active whatever the duration of the reinforcement phase (see Fig. 3). At the end of each reinforcement period, this percentage was always higher than 90% in each tank.

Learning and reinforcement processes at the group level in each tank. The x-axis is expressed in days whereas the y-axis is the percentage of successful triggering activity either on Self-feeder 1 (S1) or Self-feeder 2 (S2). The different reinforcement periods are indicated under x-axis. SF1 off = time when the S1 is turned off after the fish population in the tank reached 90% of success. In the same time S2 is turned on; S2 off = time when the S2 is turned off after the fish population in the tank reached 90% of success. In the same time, S1 is turned on; T1 to T4 = Tank 1 to Tank 4 (Colour figure online)
The learning process duration tended to decrease from the first to the last phase of the experiment (see Fig. 4); that is, there was a significant phase effect, H(3, 44) = 15,0, p = .002. the 1-day phase being shorter than the 5-day and the 7-day phases (p = .03 and p = .01, respectively). There were no significant effects of self-feeder or tank.

Learning duration (mean ± SD) at the group level. This was the time (days) required to reach 90% of success on the active self-feeder according to the different reinforcement phases durations applied (in days): 7, 5, 3, and 1
At the group scale, taking into account the individual variability, there was no significant increase or decrease of the triggering activity between the beginning and the end of the learning period on SF2 (z = -0.09, p = 1) when it was active while there were significant learning performances in all other conditions (at 10% alpha risk, SF2-Start Off vs SF2-End Off: z = - 4.5, p < .001; SF1-Start Off vs SF1-End Off : z = -3.7, p < .001; SF1-Start On vs SF1-End On: z = 2.0, p = .098).
Learning process at the individual level
GLMM showed a significant interaction between Period_Self and Code (LRT, df = 3, χ2 = 1095.2, p < .001) and a significant effect of Individuals (LRT, df =10, χ2 = 4996.9, p < .001). We used the following model to analyse the learning process: glmm (TA~Period_Self * Code + (Period_Self|ID), family = Poisson(link = "log")), with TA (Individual triggering activity: number of trigger actuations), Period_Self (Start-SF1, Start-SF2, End-SF1, End-SF2), Code (Off, On), and ID (Individual identity).
The following numbers in brackets corresponds to fish identity. There was only one individual showing some triggering activity on both self-feeders in Tanks1 (363), 2 (388), and 4 (869) and two individuals in Tank 3 (779, 699; see Table 3). The learning performances of all tested individuals were not similar. Three individuals (363, 779, and 869) showed significant learning performances on both self-feeders. Five individuals (593, 388, 371, 112, 699) showed significant learning performances only on SF1. All other individuals showed some triggering activity on one self-feeder only, associated or not with significant learning performances (no significant decrease or increase of the triggering activity between the beginning and the end of the learning period; see Table 3).
Discussion
The aim of this study was to assess whether groups of sea bass were able to learn switching from one self-feeder device to another, depending on whether they were active or not. This learning ability was examined at both group and individual levels. At the group level, operant conditioning was demonstrated, and all tanks reached the learning criteria in a few days, whatever the reinforcement period duration. The individual-level analysis revealed a more complex situation, with fish showing different learning performances that could be best explained through the producer-scrounger game theory and allows a better understanding of the individual specialization built around the self-feeder devices.
Biological performances
Growth performances recorded in the present study were in the same range as those from studies on the same species at a similar age (Benhaïm et al., 2011; Covès et al., 2006; Millot et al., 2008). There was between-tank variation that can be explained by learning differences between tanks, as already shown in the same studies. According to previous studies (Saillant, Fostier, Menu, Haffray & Chatain, 2001; Ferrari, Chatain, et al., 2014) females were found to grow better, which we also observed in the present study. We did not find sex or growth to be linked with triggering activity, which is in accordance with most studies on sea bass under self-feeding conditions (Benhaïm et al., 2012; Benhaïm et al., 2011; Covès et al., 2006; Covès, Gasset, Lemarié, & Dutto, 1998; Ferrari, Benhaïm, et al., 2014; but see Millot et al. (2008). However, our study is the first to take the individual triggering activity rather than the individual triggering category (HT, LT, and ZT) into account in the analysis of sex differential growth patterns.
Learning at the group level
All groups from all tanks reached the learning criteria between 1.9 and 6.4 days, depending on the tank and the reinforcement period duration. Previous studies have already shown that sea bass were able to operate self-feeding systems under a variety of experimental conditions (Covès et al., 2006). The present work is, however, the first one showing that at the group level, sea bass is able to switch from one device to another when the first one is turned off and the second turned on. This means that sea bass groups are not only able to be operant conditioned (i.e. linking triggering activity to the food reward delivered for all the fish in the tank) but also to recognize the difference between an inactive and an active device. This ability was not altered by the decrease of the positive reinforcement period duration, the learning criteria being reached even faster at the end of the experiment. Interestingly, better learning performances were observed when self-feeder devices were inactive. This is similar to the faster and more reliable learning obtained with negative reinforcement as compared to the positive one (Mills et al., 2010).
This confirms the high learning ability of this species that is able to deal with new situations, which is an important driving force of evolution (Kotrschal & Taborsky, 2010). As already explained in a previous study (Lieberman, 1990), learning and motivation can enhance performance, and much of the improvement is the result of skill development or increased speed, and the period when associations are formed is likely to be much shorter. This group-level approach is clearly a good alternative to the individual-based experiments performed in maze because we were able to successfully record sea bass highly objective computerized learning data during 105 days, minimizing at the same time experimenter bias and other experimental conditions on the animal’s behaviour. This improves, therefore, the reliability and validity of the results obtained in such fish learning experiments. As developed below, the design of our experiment also allowed us to track the fish individually, which was necessary to better understand how cognition at the individual level triggers the group response.
Learning at the individual level
First of all, a second self-feeder device in each tank did not modify the individual specialization usually observed with one self-feeder only. Specifically, a few individuals were responsible for most of the triggering activity in the tank (Benhaïm et al., 2012; Benhaïm et al., 2011; Covès et al., 2006; Ferrari, Benhaïm, et al., 2014; Millot et al., 2008).
High triggering fish were, however, not always the same on both self-feeders. Interestingly, there was only one individual showing a triggering activity consistently on both self-feeders in each tank, but at the same time some other individuals focused their activity on one self-feeder (either SF1 or SF2). In one tank, there was only one high triggering fish responsible for most of the activity on both self-feeders. In another tank, one individual showed the highest triggering activity on both self-feeders, but there were also a couple of individuals showing a triggering activity either on SF1 or SF2. Finally, in the last two tanks, some individuals focused their activity either on SF1 or SF2, and their activity were higher than those recorded for the individual showing a triggering activity on both self-feeders.
When looking at the learning process at the individual level in each tank, the best performances were observed on single fish showing a triggering activity on both self-feeders. These individuals decreased their activity when SF1 or SF2 were turned off and increased it when it was turned on. However, the differences between the beginning and the end of the learning period were not always significant when the self-feeder was active. In most cases, it does not mean that the high triggering fish was not able to learn the task but that the triggering activity was high as soon as the self-feeder was turned on, making the difference between the first days and the last days not significant. Therefore, these individuals demonstrated higher flexibility which is in accordance with our previous results showing that high triggering fish were characterized by shy personality trait (Ferrari, Benhaïm, et al., 2014). Shyness is indeed associated with a reactive strategy; individuals showing a higher behavioural flexibility (Bolhuis et al., 2004) and higher responsiveness to their environment (Verbeek, Drent, & Wiepkema, 1994). In one tank, however, the triggering activity remained almost the same when one self-feeder was turned off, but the triggering activity on the active one was much higher. In other words, some individuals showed a triggering activity on both inactive and active devices. Other individuals that focused their activity on one self-feeder performed variously according to the tank, but in all cases there was at least one individual showing good learning performances.
These results reveal how the behaviour of individual components scales to that at the group level which is a fundamental problem in a wide range of biological disciplines including the cognitive sciences (Kitano, 2002). Below we attempt to explain the link between individual and group behaviour through the producer-scrounger game theory.
Link between individual and group levels
How can we explain why individuals among the group adopt different strategies when facing two self-feeders alternatively activated? The first important point is that learning rules are not strict causal models of learning based on psychological mechanisms (Beauchamp, 2000).
Previous studies on social information transfer in schooling fish showed that an informed individual that has been trained to perform a task (associating a spatial target with a food reward) behaved differently within a group of uninformed individuals (Ioannou, Singh, & Couzin, 2015). The informed individual efficiency based distribution then turned as a normal distribution that can be explained by a trade-off between reaching the goal in the most effective way and the cost of isolation by leaving followers behind. Interestingly, in our study, a few individuals only performed the task (activating the trigger efficiently, i.e. HT fish) and the social structure based on triggering activity remained consistent over time. According to Ioannou, Singh, and Couzin (2015), a modification of the social structure (a decrease of high triggering fish activity and an increase of low triggering fish activity) could have been expected, leading to a normal distribution of activity between individuals. This can be explained by our experimental conditions where the social structure is regulated by a different trade-off linked to food resources availability and acquisition. According to the producer-scrounger game theory, individuals in many species search for resources in groups, and foraging payoff strongly depends on the behaviour of other congeners and available resources. In the present work, the high triggering fish that was able to switch from one self to the other could play the role of the producer actively searching and finding food sources that may be shared with other group members. This strategy could be used because it provides them with the greatest payoff for their phenotype (shy personality trait) (Giraldeau & Beauchamp, 1999; Ferrari, Benhaïm, et al., 2014). On the contrary, other individuals that never or seldom actuate the trigger clearly seem to play the role of scroungers. The high proportion of scroungers always observed in self-feeding conditions can easily be explained by the fact that there is no food limitation. It should be remembered here that when the high triggering fish actuates the trigger, it delivers food for all the fish in the tank. Interestingly, in our experiment, some individuals adopted different strategies, and some new triggering fish appeared at the switch between feeders. This can be certainly linked to the results obtained by Di-Poï et al. (2008), who investigated the effects of removal of the HT fish on the social and feeding behaviours in groups of juvenile sea bass. These authors showed that following removal, new fish (coming from the group of fish having a low number of triggering actuations) displaying a dominant self-feeding activity quickly appear. In the present work, when the first self-feeder was turned off, the HT fish continued to trigger it during the learning process before switching to the active one. In addition, learning performances were shown to be lower on the self-feeder two than the self-feeder one, leading to a decrease of high triggering fish activity on self-feeder two. As a consequence, less food was delivered during this period in the tank, which induced some other individuals showing previously a low triggering activity to become more active. As assumed by the producer-scrounger game theory, the payoffs obtained by scroungers are negatively frequency dependent. Increasing the cost of scrounging leads to a decrease in the observed proportion of scroungers, whereas raising the cost of producing increases the proportion of scroungers (Morand-Ferron, Giraldeau, & Lefebvre, 2007). Individuals that specialized on one single self-feeder either active or inactive that we could call ‘bad learners’ from a strict causal model of learning could also use a tactic providing them with the greatest payoff for their phenotype.
The individual decision to switch from a scrounger to a producer tactic involves that the fish is able to learn the task, which could be facilitated by the group situation. Among its very well-known adaptive advantages, schooling could indeed improve the flow of information through the group (Couzin, Krause, Franks, & Levin, 2005). Social interactions could greatly improve the absolute ability to efficiently complete a task by social transmission, as already suggested in previous studies on fish (Ioannou, Couzin, James, Croft, & Krause, 2011) and on sea bass in particular under self-feeding conditions (Anthouard, 1987; Di-Poï et al., 2008).
Conclusion
In conclusion this experiment is the first attempt to study sea bass cognition under group conditions, with a design allowing at the same time to follow the performances of each individual, providing therefore a way to avoid the serious drawbacks usually met in isolated conditions. We argue that studying fish cognition could be much more relevant and meaningful in such group conditions because it corresponds to a more realistic situation. The study of animal groups presents an opportunity to reveal how the behaviour of individual components scales to that at higher organizational levels which is a fundamental problem in a range of biological disciplines, not least the cognitive sciences (Kitano, 2002). Cognition is just one of several factors that can explain how animals adapt to their environment. Further research is needed to better understand the link between individual and group cognition in fish. The computerized self-feeder device opens new perspectives in this way. It could help to link different concepts such as cognition, personality, individual specialization, and information transfer in fish groups. We suggest, for example, to build specific populations and/or to modify the amount of food delivered in the tank in order to understand the decision-making process and the role of each individual for the group.
Change history
05 June 2017
An erratum to this article has been published.
References
Al-Imari, L., & Gerlai, R. (2008). Sight of conspecifics as reward in associative learning in zebrafish (Danio rerio). Behavioural Brain Research, 189, 216-219.
Anthouard, M. (1987). A study of social transmission in juvenile Dicentrarchus labrax (Pisces, Serranidae), in an operant conditioning situation. Behaviour, 103, 266–275.
Barnard, C. J., & Sibly, R. M. (1981). Producers and scroungers: A general model and its application to captive flocks of house sparrows. Animal Behaviour, 29, 543–550.
Bates, D., Mächler, M., Bolker, B., & Walker, S. (2014). Fitting linear mixed-effects models using lme4. ArXiv14065823 Stat. doi:10.18637/jss.v067.i01
Beauchamp, G. (2000). Learning rules for social foragers: Implications for the producer-scrounger game and ideal free distribution theory. Journal of Theoretical Biology, 207, 21–35.
Benhaïm, D., Bégout, M.-L., Lucas, G., & Chatain, B. (2013). First insight into exploration and cognition in wild caught and domesticated sea bass (Dicentrarchus labrax) in a maze. PLoS ONE, 8(6), e65872.
Benhaïm, D., Bégout, M.-L., Péan, S., Brisset, B., Leguay, D., & Chatain, B. (2012). Effect of fasting on self-feeding activity in juvenile sea bass (Dicentrarchus labrax). Applied Animal Behaviour Science, 136(1), 63–73.
Benhaïm, D., Bégout, M.-L., Péan, S., Manca, M., Prunet, P., & Chatain, B. (2013). Impact of a plant-based diet on behavioural and physiological traits in sea bass (Dicentrarchus labrax). Aquatic Living Resources, 26(02), 121–131. doi:10.1051/alr/2013049
Benhaïm, D., Péan, S., Bégout, M.-L., Brisset, B., Leguay, D., & Chatain, B. (2011). Effect of size grading on sea bass (Dicentrarchus labrax) juvenile self-feeding behaviour, social structure and culture performance. Aquatic Living Resources, 24, 391–402. doi:10.1051/alr/2011140
Bilotta, J., Risner, M. L., Davis, E. C., & Haggbloom, S. (2005). Assessing appetitive choice discrimination learning in zebrafish. Zebrafish, 2, 259–268.
Blaser, R. E., & Vira, D. G. (2014). Experiments on learning in zebrafish (Danio rerio): A promising model of neurocognitive function. Neuroscience & Biobehavioral Reviews, 42, 224–231. doi:10.1016/j.neubiorev.2014.03.003
Bolhuis, J. E., Schouten, W. G. P., Leeuw, J. A. D., Schrama, J. W., & Wiegant, V. M. (2004). Individual coping characteristics, rearing conditions and behavioural flexibility in pigs. Behavioural Brain Research, 152, 351–360.
Brown, C., Laland, K., & Krause, J. (2007). Fish cognition and behavior. Hoboken: Blackwell.
Chatain, B. (1994). Estimation et amélioration des performances zootechniques de l’élevage larvaire de Dicentrarchus labrax et de Sparus auratus [Assesment and improvement of Dicentrarchus labrax and Sparus aurata growth performances in hatchery] (Doctoral thesis, Université d'Aix Marseille II, France).
Colwill, R. M., Raymond, M. P., Ferreira, L., & Escudero, H. (2005). Visual discrimination learning in zebrafish (Danio rerio). Behavioural Processes, 70(1), 19–31.
Couzin, I. D., Krause, J., Franks, N. R., & Levin, S. A. (2005). Effective leadership and decision-making in animal groups on the move. Nature, 433, 513–516.
Covès, D., Beauchaud, M., Attia, J., Dutto, G., Bouchut, C., & Bégout, M. L. (2006). Long-term monitoring of individual fish triggering activity on a self-feeding system: An example using European sea bass (Dicentrarchus labrax). Aquaculture, 253(1/4), 385–392.
Covès, D., Gasset, E., Lemarié, G., & Dutto, G. (1998). A simple way of avoiding feed wastage in European seabass, Dicentrarchus labrax, under self-feeding conditions. Aquatic Living Resources, 11(6), 395–401.
Di-Poï, C., Attia, J., Bouchut, C., Dutto, G., Covès, D., & Beauchaud, M. (2007). Behavioral and neurophysiological responses of European sea bass groups reared under food constraint. Physiology & Behavior, 90, 559–566. doi:10.1016/j.physbeh.2006.11.005
Di-Poï, C., Beauchaud, M., Bouchut, C., Dutto, G., Covès, D., & Attia, J. (2008). Effects of high food demand fish removal in groups of juvenile sea bass (Dicentrarchus labrax). Canadian Journal of Zoology, 86, 1015–1023. doi:10.1139/Z08-077
Dudchenko, P. A. (2001). How do animals actually solve the maze? Behavioral Neuroscience, 115(4), 850–860.
Ferrari, S., Benhaïm, D., Colchen, T., Chatain, B., & Bégout, M.-L. (2014). First links between self-feeding behaviour and personality traits in European seabass, Dicentrarchus labrax. Applied Animal Behaviour Science, 161, 131–141. doi:10.1016/j.applanim.2014.09.019
Ferrari, S., Chatain, B., Cousin, X., Leguay, D., Vergnet, A., & Vidal, M.-O. (2014). Early individual electronic identification of sea bass using RFID microtags: A first example of early phenotyping of sex-related growth. Aquaculture, 426–427, 165–171. doi:10.1016/j.aquaculture.2014.01.033
Fox, J., & Weisberg, S. (2011). An R companion to applied regression (2nd ed.). Thousand Oaks: SAGE.
Gibson, B. M., & Shettleworth, S. J. (2005). Place versus response learning revisited: Tests of blocking on the radial maze. Behavioral Neuroscience, 119(2), 567–586.
Giraldeau, L.-A., & Beauchamp, G. (1999). Food exploitation: Searching for the optimal joining policy. Trends in Ecology & Evolution, 14, 102–106.
Grossman, L., Stewart, A., Gaikwad, S., Utterback, E., Wu, N., & DiLeo, J. (2011). Effects of piracetam on behavior and memory in adult zebrafish. Brain Research Bulletin, 85, 58–63. doi:10.1016/j.brainresbull.2011.02.008
Hothorn, T., Bretz, F., & Westfall, P. (2008). Simultaneous inference in general parametric models. Biometrical Journal, 50, 346–363.
Ioannou, C. C., Couzin, I. D., James, R., Croft, D. P., & Krause, J. (2011). Social organization and information transfer in schooling fish. In C. Brown, K. Laland, & J. Krause (Eds.), Fish cognition and behaviour (2nd ed.). Oxford: Wiley-Blackwell. doi:10.1002/9781444342536.ch10
Ioannou, C. C., Singh, M., & Couzin, I. D. (2015). Potential leaders trade off goal-oriented and socially oriented behavior in mobile animal groups. The American Naturalist, 186(2), 284–293. doi:10.1086/681988
Karnik, I., & Gerlai, R. (2012). Can zebrafish learn spatial tasks? An empirical analysis of place and single CSUS associative learning. Behavioural Brain Research, 233, 415–421.
Kitano, H. (2002). Systems biology: A brief overview. Science, 295, 1662–1664.
Kotrschal, K., & Taborsky, B. (2010). Environmental change enhances cognitive abilities in fish. PLoS Biology, 8(4), e1000351.
Lieberman, D. A. (1990). Learning: Behavior and cognition. Belmont: Wadsworth.
Lopez, J. C., Broglio, C., Rodriguez, F., Thimus-Blanc, C., & Salas, C. (1999). Multiple spatial learning strategies in goldfish (Carassius auratus). Animal Cognition, 2, 109–120.
Martins, C. I. M., Schrama, J. W., & Verreth, J. A. J. (2005). Inherent variation in growth efficiency of African catfish Clarias gariepinus (Burchell, 1822) juveniles. Aquaculture Research, 36, 868–875.
Millot, S., & Bégout, M.-L. (2009). Individual fish rhythm directs group feeding: A case study with sea bass juveniles (Dicentrarchus labrax) under self-demand feeding conditions. Aquatic Living Resources, 22, 363–370.
Millot, S., Bégout, M.-L., Person-Le Ruyet, J., Breuil, G., Di-Poï, C., & Fievet, J. (2008). Feed demand behavior in sea bass juveniles: Effects on individual specific growth rate variation and health (inter-individual and intergroup variation). Aquaculture, 274, 87–95.
Mills, D. S., Marchant-Forde, J. N., McGreevy, P. D., Morton, D. B., Nicol, C. J., & Phillips, C. J. C. (2010). The encyclopedia of applied animal behaviour and welfare. Wallingford: CAB International.
Morand-Ferron, J., Giraldeau, L.-A., & Lefebvre, L. (2007). Wild grackles play a producer-scrounger game. Behavioral Ecology, 18, 916–921.
Mueller, K. P., & Neuhauss, S. C. (2012). Automated visual choice discrimination learning in zebrafish (Danio rerio). Journal of Integrative Neuroscience, 11, 73–85.
Parker, M. O., Gaviria, J., Haigh, A., Millington, M. E., Brown, V. J., & Combe, F. J. (2012). Discrimination reversal and attentional sets in zebrafish (Danio rerio). Behavioural Brain Research, 232(1), 264–268. doi:10.1016/j.bbr.2012.04.035
Pather, S., & Gerlai, R. (2009). Shuttle box learning in zebrafish (Danio rerio). Behavioural Brain Research, 196, 323–327.
R core Team (2015). R: A langage and environment for statistical computing, R Foundation for statistical computing. Vienna, Austria.
Rubio, V. C., Vivas, M., Sanchez-Mut, A., Sanchez-Vazquez, F. J., Coves, D., & Dutto, G. (2004). Self-feeding of European sea bass (Dicentrarchus labrax, L.) under laboratory and farming conditions using a string sensor. Aquaculture, 233, 393–403. doi:10.1016/j.aquaculture.2003.10.011
Saillant, E., Fostier, A., Menu, B., Haffray, P., & Chatain, B. (2001). Sexual growth dimorphism in sea bass Dicentrarchus labrax. Aquaculture, 202, 371–387.
Shettleworth, S. J. (2009). The evolution of comparative cognition: Is the snark still a boojum? Behavioural Processes, 80(3), 210–217. doi:10.1016/j.beproc.2008.09.001
Sison, M., & Gerlai, R. (2010). Associative learning in zebrafish (Danio rerio) in the plus maze. Behavioural Brain Research, 207(1), 99–104. doi:10.1016/j.bbr.2009.09.043
Verbeek, M. E. M., Drent, P. J., & Wiepkema, P. R. (1994). Consistent individual differences in early exploratory behaviour of male great tits. Animal Behaviour, 48(5), 1113–1121.
Volkoff, H., & Peter, R. E. (2006). Feeding behavior of fish and its control. Zebrafish, 3(2), 131–140.
Warburton, K. (2007). Learning of foraging skills by fishes. Hoboken: Blackwell.
Williams, F. E., White, D., & Messer, W. S., Jr. (2002). A simple spatial alternation task for assessing memory function in zebrafish. Behavioural Processes, 58(3), 125–132. doi:10.1016/S0376-6357(02)00025-6
Zar, J. H. (1984). Biostatistical analysis (2nd ed.). Englewood Cliffs: Prentice Hall.
Acknowledgements
This experiment was made possible through funding provided by the Conseil Régional de Basse-Normandie and the Syndicat Mixte du Cotentin. We are grateful to Pr. Ludovic Dickel and Dr. Christophe Féron for precious help on the design of our experiment. Thanks to Dr. Christos Ioannou for helpful comments on a previous version of this manuscript. We also thank Khaled Horri and Régis Gallon for invaluable statistical help. We finally thank Didier Leguay for technical help.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Ethical standards
This study was conducted under the approval of the Animal Care Committee of France under the official licence of M.L. Bégout (17-010).
Additional information
The original version of this article was revised: The Given Name and Family Name of the second author are inverted. The correct order is Sébastien Ferrari.
David Benhaïm and Sébastien Ferrari contributed equally to this work.
An erratum to this article is available at https://doi.org/10.3758/s13420-017-0276-z.
Rights and permissions
About this article

Cite this article
Benhaïm, D., Ferrari, S., Colchen, T. et al. Relationship between individual and group learning in a marine teleost: A case study with sea bass under self-feeding conditions. Learn Behav 45, 276–286 (2017). https://doi.org/10.3758/s13420-017-0266-1
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13420-017-0266-1