
Abstract
According to the mixed memory model (Penney, Gibbon, & Meck, Journal of Experimental Psychology: Human Perception and Performance, 26, 1770–1787, 2000), different clock rates for stimuli with different nontemporal properties must be stored within a single reference memory distribution in order to detect a difference between the clock rates of the different signals. In Experiment 1, pigeons were trained in a between-subjects design to discriminate empty intervals (bound by two 1-s visual markers) and filled intervals (a continuous visual signal). The intervals were signaled by different visual stimuli, and they required responses to different sets of comparison stimuli. Empty intervals were judged as being longer than filled intervals. The difference between the point of subjective equality (PSE) for the empty intervals and the PSE for the filled intervals increased proportionally as the magnitudes of the anchor duration pairs were increased from 2 and 8 s to 4 and 16 s. In Experiment 2, the pigeons were trained to discriminate intervals signaled by the absence of houselight illumination (Group Empty) or the presence of houselight illumination (Group Filled). The psychophysical timing functions for these intervals were identical to each other. The results of Experiment 1 indicate that memory mixing is not necessary for detecting a timing difference between empty and filled intervals in pigeons. The results of Experiment 2 suggest that the nature of the stimuli that signal the empty and filled intervals impacts how pigeons judge the durations of empty and filled intervals.
Similar content being viewed by others

Interval timing has been shown to inform essential processes in many species (Gallistel, 1990). The temporal bisection procedure has often been used to study interval timing in both human and nonhuman animals (Penney, Gibbon, & Meck, 2008). Pigeons trained in a bisection procedure are required to respond to one comparison stimulus (e.g., a red hue) after a short-duration signal (e.g., 2 s), and to respond to the other comparison stimulus (e.g., a green hue) after a long-duration signal (e.g., 8 s). Once a high level of accuracy is achieved, test trials of various intermediate signal durations are presented, but responses on these trials are not reinforced. From this procedure, a psychophysical function can be generated that plots the proportion of “long” responses as a function of signal duration and allows for the calculation of the point of subjective equality (PSE). The PSE is the intermediate signal value at which the subject equally often makes a “short” response or a “long” response. Studies employing bisection procedures with animals (e.g., Church & Deluty, 1977) have reliably found that the PSE falls at the geometric mean (GM) of the two anchor durations (Gibbon, Church, & Meck, 1984; Miki & Santi, 2005; Santi, Keough, Gagne, & Van Rooyen, 2007; Santi, Miki, Hornyak, & Eidse, 2005), whereas studies with humans have shown that the PSE falls at either the GM (Allan, 1992; Allan & Gibbon, 1991) or the arithmetic mean (AM; Wearden, 1991; Wearden & Ferrara, 1995, 1996; Wearden, Rogers, & Thomas, 1997). The location of the PSE relative to the GM or the AM has been used to infer the nature of the decision rule used by subjects trained in a bisection procedure. A decision rule based on a ratio comparison of the similarity of the signal duration on the current trial to reference memories of the short and long anchor durations results in bisection at the GM (see Allan & Gibbon, 1991), whereas a decision rule based on the simple difference of the current signal duration from a memory of the short and the long anchor durations results in bisection at the AM (see Wearden, 1991). In addition, rightward and leftward shifts of the PSE have been used to determine whether a given duration is experienced as objectively shorter or longer as a function of a variety of experimental manipulations, such as signal modality (e.g., Penney, Gibbon, & Meck, 2000), the behavior required while timing (e.g., Zentall & Singer, 2008), the effect of a secondary cognitive task during timing (e.g., Molet, Alessandri, & Zentall, 2011), and drug conditions (e.g., Meck, 1983).
Previous research on time perception in humans has indicated that nontemporal properties of the time signals play an important role in the perception and discrimination of time (Allan, 1979; Goldstone & Lhamon, 1974; Grondin, 1993; Poynter, 1989). Temporal discrimination accuracy in animals is affected by stimulus intensity (Kraemer, Brown, & Randall, 1995; Wilkie, 1987) as well as stimulus modality (Meck, 1984; Roberts, Cheng, & Cohen, 1989; Stubbs, Dreyfus, & Fetterman, 1984). For example, pigeons time visual signals more accurately than auditory signals (Roberts et al., 1989), whereas the reverse is found for rats (Meck, 1984). Rats also appear to judge auditory signals as being longer than visual signals of equal duration (Meck, 1991).
Whether an interval is filled or empty affects the accuracy and variability with which it is timed (Abel, 1972a, 1972b; Rammsayer & Lima, 1991; Rammsayer & Skrandies, 1998). A filled interval is the duration of a continuously presented signal such as a light or tone, whereas an empty interval is the duration between the offset of one time marker and the onset of a second time marker. Initial findings suggested that humans judge filled intervals as being longer than empty intervals of the same duration (Craig, 1973; Goldfarb & Goldstone, 1963; Goldstone & Goldfarb, 1963), however as noted by Rammsayer (2010) differences in duration discrimination with filled and empty intervals depends on various factors, such as the duration of the intervals to be compared, the sensory modality in which the intervals are presented, the physical characteristics of the markers defining the empty intervals, or the psychophysical procedure applied (Allan, 1979; Fraisse, 1978; Grondin, 2001, 2003, 2008). Miki and Santi (2005) trained pigeons in a within-subjects design to discriminate filled intervals (short and long durations of light) and empty intervals (short and long durations signaled by two 500 ms light markers). The duration of the empty interval was the interval of time between the offset of the first marker and the onset of the second marker. Three different sets of anchor durations were used in training: 1 versus 4 s, 2 versus 8 s, and 4 versus 16 s. Unlike previous findings reported in humans, pigeons judged the empty intervals to be longer than the filled intervals. Importantly, the difference between the PSE for empty intervals and the PSE for filled intervals got larger as the magnitude of the anchor durations used in training increased. This indicates that the difference in the PSE for empty and filled intervals could not have been due to either a difference in the latency to initiate the timing or to the timing of the markers themselves on empty-interval trials, because the effect of these two factors would have been constant across the anchor duration pairs used in training. Additional evidence, supporting the claim that pigeons do not time the markers on empty-interval trials, has been obtained by conducting psychophysical tests with different marker durations, and showing that no significant increase in the percentage of “long” responding occurs as marker duration is increased (Grant & Talarico, 2004; Santi et al., 2007).
Previous research conducted on humans (Block & Zakay, 1997; Chaston & Kingstone, 2004; Zakay & Block, 1996, 1997) and on animals (Buhusi & Meck, 2006a, 2006b; Buhusi, Pascalis, & Cerutti, 2006; Lejeune, Macar, & Zakay, 1999; Sutton & Roberts, 2002) has shown that the duration of temporal intervals can be underestimated if attentional resources are diverted away from timing in order to engage in nontemporal information processing. This attention-sharing concept was extended by Santi et al. (2007) to explain the empty–filled timing difference in pigeons. They suggested that during empty intervals of darkness, pigeons could devote more attention to timing signal duration, because there would be little distracting visual information to attend to in the chamber. However, during filled intervals of a continuously illuminated signal, pigeons may have devoted less attention to temporal processing because of the availability of other visual stimuli in the chamber (opening to the food hopper, pieces of grain below the grid floor of the chamber, etc.) capturing attention. Santi et al. (2007) tested this attention-sharing explanation by manipulating the ambient illumination conditions during psychophysical test sessions. They found that empty intervals were judged to be longer than filled intervals when testing occurred in a darkened test room, but not when the test room was illuminated. Presumably this occurred because when ambient illumination was continually present during test sessions, the nontemporal visual features of the test environment would be available to divert attention from temporal processing on both filled-interval and empty-interval trials. As a result, no timing difference between filled and empty intervals would be found. Additionally, Santi et al. (2007) showed that if different tonal frequencies were used to mark empty intervals and to signal filled intervals, no evidence was apparent of an overestimation of empty relative to filled intervals. This result is also consistent with the idea that competition for attention by visual features is responsible for the empty–filled timing difference exhibited by pigeons.
Recently, it has been demonstrated that pigeons underestimate the duration of filled intervals if pecking is required during signal presentation, and they overestimate the duration of filled intervals if they are required to refrain from pecking (Zentall & Singer, 2008). In order to determine whether key pecking during filled intervals, but not during empty intervals might be responsible for the empty-filled timing difference, Santi, Adams, and Bassett (2010) trained pigeons in a within-subjects design to discriminate empty intervals (bound by two 1-s visual markers) and filled intervals (a continuous visual signal). Santi et al. (2010) replicated the previously reported empty–filled timing differences and found that although there was more pecking during filled than during empty intervals, no significant correlation emerged between pecking during filled intervals and the value of the PSE. More importantly, when they explicitly required the pigeons to refrain from pecking during filled intervals, they still found that empty intervals were judged to be longer than filled intervals. Thus, the empty–filled timing difference in pigeons does not appear to be due differences in key pecking during filled and empty intervals.
Penney et al. (2000) proposed a mixed memory version of scalar timing theory to account for the effects of different stimulus modalities on timing in humans (auditory vs. visual). According to this model, differences in timing due to modality arise from effects at two processing levels. The first process is a clock process, in which auditory stimuli drive the pacemaker/accumulator component at a higher rate than visual stimuli. The second process is a reference memory process that mixes the clock values for auditory and visual stimuli. Although this model makes no prediction about whether a timing difference will occur for empty and filled intervals, it does provide a possible explanation of this difference in terms of a slower clock rate for filled intervals, and it may provide a basis for understanding the experimental conditions necessary to observe the difference. According to the model, even if the clock accumulated pulses at a slower rate for filled intervals than for empty intervals (because of attention-sharing), when pigeons are trained to time only one type of interval, reference memory would only consist of values for that particular type of interval. As a result, when signal durations timed with a slower clock are compared to memory distributions for a signal timed with an equally slow clock (i.e., comparing the filled interval duration on the current trial to the reference memory of filled interval durations), and signals timed with a faster clock are compared to memory distributions for a signal timed with an equally fast clock (i.e., comparing the empty interval duration on the current trial to the reference memory of empty interval durations), no timing differences would be expected. In order to observe a timing difference, the model claims that the accumulator values for the empty and filled intervals must be stored within a single reference memory distribution representing the short values for the two intervals and within a single reference memory distribution representing long values for the two intervals. Consequently, an empty–filled timing difference could be detected if the empty and filled intervals were studied within subjects, as in our previous studies (Miki & Santi, 2005; Santi et al., 2010; Santi et al., 2007), but not if they were studied in a between-subjects design. The purpose of the present experiment was to determine whether the empty–filled timing difference would be eliminated if pigeons were trained to only time one type of interval in each session (between-subjects design).
Experiment 1
Method
Subjects
A group of 16 adult male homing pigeons, maintained at approximately 80 % of their free-feeding weights, served as the subjects. The birds were housed individually with continual access to water and grit throughout the duration of the experiment, in a colony room that was illuminated on a 12:12 h light:dark cycle, with lights turning on at approximately 7:00 a.m. each day. Testing was conducted five days per week between 9:00 a.m. and 4:00 p.m. The birds had not previously served in experiments involving timing of empty and filled intervals.
Apparatus
Four Coulbourn modular operant test cages (Model E10-10) were used. Each test cage was housed within isolation cubicles (Model 10-20), and each cubicle used baffled air intake exhaust systems and ventilation fans. Each test cage contained three horizontally aligned, translucent plastic keys positioned approximately at a pigeon’s standing sightline. Behind each key a projector displayed the experimental stimuli onto a frosted rear projection screen (Coulbourn Model E21-18). The stimuli included hues (red and green), line orientations (vertical and horizontal), and shapes (circle and triangle). All line orientations and shapes were white and presented against a black background. Directly below the center key a 5.75-cm opening provided access to a hopper containing mixed grain. Within the opening a lamp (Coulbourn Model E14-10 with bulb S11819X) was illuminated only during reinforcement. A houselight (Coulbourn Model 14-10) that permitted light to be directed upward to the top of the chamber was located 6.5 cm above the center key. The houselight was not used in Experiment 1, but it was used in Experiment 2. The presentation of all experimental events and the recording of response choices were accomplished through a Med Associates interface and a microcomputer running MED PC-IV State Notation programming software.
Procedure
All pigeons were initially trained to eat mixed grain from the illuminated food hopper and were autoshaped to peck at red and green, at a white vertical line, and at a white horizontal line presented randomly on either the left or the right response key. This initial training continued until all pigeons reliably pecked all of the stimuli presented on both of the response keys.
After pretraining, eight pigeons were randomly assigned to Group Empty and trained to discriminate between a short (2-s) and a long (8-s) interval signaled by two visual markers presented on the center key (1-s duration each). One indicated the beginning of the interval; the other, the end. The remaining eight pigeons were assigned to Group Filled and trained to discriminate between a short (2-s) and a long (8-s) continuous visual signal presented on the center key. For four pigeons in Group Empty, the marker was a white triangle, whereas for the other four pigeons it was a white circle. Similarly, for four pigeons in Group Filled, the visual signal was a white triangle, whereas for the remaining four pigeons it was a white circle. Comparison stimuli were presented simultaneously on the side keys for both groups. For four pigeons in Group Empty, red and green comparisons were presented after the white triangle marked the interval, and vertical and horizontal comparison stimuli were presented after the white circle marked the interval. This was reversed for the remaining four pigeons. For four pigeons in Group Filled, the red and green comparisons were presented after the white circle signaled the interval, and the vertical and horizontal comparison stimuli were presented after the white triangle signaled the interval. This was reversed for the remaining four pigeons. For both groups, the comparison stimuli that were designated correct following the short duration were red and vertical, and following the long duration they were green and horizontal.
For all of the pigeons, a single response to one of the comparisons turned both comparison stimuli off, and, if correct, provided access to mixed grain for 3 s. Incorrect responses to the comparison stimuli resulted in a blackout of 3 s, followed immediately by re‑presentation of the same interval duration and comparison stimulus configuration. A correct response on a correction trial resulted in access to mixed grain, although only the choice response on the initial (noncorrection) trial was used to calculate response accuracy.
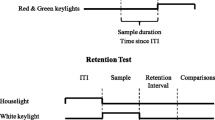
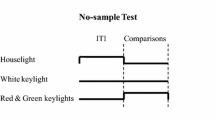
At the beginning of each trial, a randomly selected intertrial interval (4, 8, 16, or 32 s) was presented. No illumination was presented in the test chamber during the intertrial interval (ITI). At the end of the ITI, the signal was presented on the center key to pigeons in the filled-interval condition and the marker was presented on the center key to pigeons in the empty-interval condition. After the signal or the second marker terminated, the comparison stimuli were presented on the left- and right-side keys, counterbalanced across trials.
Each session of training for both groups consisted of 160 trials. In each block of eight trials, all combinations of the sample duration (short or long) and the two comparison stimulus configurations (red/green or vertical/horizontal) occurred twice. The order of the presentation was randomized individually for each bird. Each pigeon received a minimum of 15 sessions of training. After 15 sessions of training, a pigeon was advanced to psychophysical testing if it had achieved 80 % accuracy or above on both the short and long anchor durations for three consecutive sessions.
Each psychophysical test session consisted of 160 trials. Group Filled continued to only receive filled intervals and Group Empty only received empty intervals. The anchor durations were presented on 80 trials and the intermediate durations were presented on the remaining 80 trials. The intermediate durations were 2.6, 3.2, 4.0, 5.0, and 6.4 s. In each session, all intermediate durations were randomly tested sixteen times. The pigeons were still reinforced for responding correctly following the two training durations, but responses following intermediate durations were never reinforced. Twenty psychophysical test sessions were administered.
After psychophysical testing, the birds were retrained with 4- and 16-s anchor durations.
Apart from the increased anchor duration values, the discrimination training was the same as that described above. A minimum of ten sessions of training with the 4- and 16-s anchor durations were given. A pigeon was advanced to psychophysical testing if it had achieved 80 % accuracy or above on both the short and long anchor durations for three consecutive sessions. Seven of the eight pigeons in both Group Empty and Group Filled met the criteria to advance to testing after ten sessions. One pigeon in Group Empty and one in Group Filled required ten and six sessions of additional training, respectively. Over the final three sessions of training prior to psychophysical testing, the accuracy for Group Filled (M = 94.87, SD = 3.58) was significantly higher than that for Group Empty (M = 90.13, SD = 1.49), t(14) = 3.45, p < .05. Twenty sessions of psychophysical testing with intermediate durations of 5.2, 6.4, 8.0, 10.0, and 12.8 s were administered. All other aspects of the test procedure were similar to those described above.
In all statistical analyses reported, the rejection region was p < .05.
Results and discussion
Figure 1 illustrates the mean percentages of correct responding for Group Filled and Group Empty during the first 15 sessions of acquisition training across blocks of three sessions. As is illustrated in the figure, Group Filled acquired the duration discrimination faster than did Group Empty. Averaged over the last block of three sessions, the mean percentage correct for Group Filled, M = 93.83, SD = 4.61, was higher than the mean percentage correct for Group Empty, M = 82.06, SD = 8.16. An analysis of variance (ANOVA) was conducted on the first 15 sessions of acquisition training with Group as a between-subjects factor and Block as a within-subjects factor. The analysis revealed significant main effects of group, F(1, 14) = 6.82, MSE = 250.20, and block, F(4, 56) = 111.72, MSE = 26.89. As well, we found a significant Group × Block interaction, F(4, 56) = 8.00, MSE = 26.89: The accuracy for Group Filled was significantly higher than that for Group Empty during Blocks 3, 4, and 5, Fs(1, 14) = 7.84, 23.69, and 12.61, MSEs = 115.53, 44.73, and 43.97, respectively. All pigeons in Group Filled and four pigeons in Group Empty began testing after 15 sessions of training. The remaining four pigeons in Group Empty required between one and 14 additional sessions of discrimination training (i.e., a total of 15 to 29 sessions of training) to reach the testing criterion. These results replicate previous reports of faster acquisition for filled than for empty intervals (Kraemer, Randall, & Brown, 1997; Miki & Santi, 2005).

Mean percentages of correct responding for filled and empty intervals during acquisition of the discrimination between the 2- and 8-s anchor durations. Error bars represent the standard errors of the means
Figure 2 presents the psychophysical functions that were obtained following 2-versus-8-s training (top panel), and 4-versus-16-s training (bottom panel). The psychophysical functions for Group Empty were horizontally displaced to the left of the functions for Group Filled. That is, empty intervals were perceived as being longer than filled intervals of the same duration. The lines through the data points are the best-fitting functions that were obtained by using the four-parameter sigmoid equation in SigmaPlot 9, P(t) = a + b/(1 + exp[–(t – T50)/c]). P(t) is the percentage of “long” responding, and t is the signal duration. T50 is the PSE, which indicates the signal duration at which a subject was equally likely to select the “short” or the “long” response alternative. The slope of the function (c) provides an index of sensitivity to time, and the lower asymptote (a) and upper asymptote (b) indicate the percentages of “long” responding for the shortest and longest stimulus durations, respectively. The mean estimates of the PSE, slope, and asymptotes obtained by fitting four-parameter sigmoid functions to the individual pigeon data are shown in Table 1. For the 2- and 8-s anchor durations, the mean PSE for Group Empty was significantly less than the mean PSE for Group Filled, t(14) = 3.51. In addition, the mean PSE for Group Empty fell significantly below the geometric mean (GM) of 4 s, t(7) = 3.22, but the mean PSE for Group Filled did not significantly differ from the GM, t(7) = 1.72. For the 4- and 16-s anchor durations, the mean PSE for Group Empty was significantly less than the mean PSE for Group Filled, t(14) = 4.76, and the mean PSE for Group Empty was significantly below the GM of 8 s, t(7) = 3.96, whereas the mean PSE for Group Filled was significantly above the GM, t(7) = 2.64. As can be seen in Table 1, the between-group difference in the mean PSEs for the 4- and 16-s anchor durations (1.52) was slightly more than twice the difference obtained for the 2- and 8-s anchor durations (0.71). To confirm that the mean PSE for the 4- and 16-s durations was double the value of the mean PSE for the 2- and 8-s durations, for each pigeon the PSE estimated for the 2- and 8-s durations was doubled, and the difference between this value and the actual PSE obtained for the 4- and 16-s durations was examined. The resulting difference scores for both Group Empty (M = –0.10, SD = 0.91) and Group Filled (M = 0.02, SD = 0.98) did not differ significantly from zero, ts < 1.

Upper panel: Mean percentages of “long” responses as a function of signal duration for filled and empty intervals following training with anchor durations of 2 and 8 s. Lower panel: Percentages of “long” responses as a function of signal durations of 4 and 16 s. Error bars represent the standard errors of the means
Although we observed a clear difference in timing, the sensitivities to time reflected in the slopes of the psychophysical functions were equivalent for Group Filled and Group Empty during both the 2-versus-8-s test and the 4-versus-16-s test, ts < 1. The asymptotes of the psychophysical functions are also summarized in Table 1. The lower asymptote for Group Empty was higher than that for Group Filled at both the 2-versus-8-s test and the 4-versus-16-s test. However, neither of these differences was statistically significant by a two-tailed test. The upper asymptote for Group Empty was significantly lower than that for Group Filled at both the 2-versus-8-s test and the 4-versus-16-s test, ts(14) = 4.17 and 4.29, respectively.
The evidence presented above that the mean PSEs for empty and filled intervals were multiples of the magnitudes of the anchor duration pairs leads to the expectation that the psychophysical functions should superimpose across anchor duration sets. Figure 3 shows these superimposition plots, and as expected, there was very good superposition for the different anchor duration sets, indicative of a proportional shift in the functions. However, the psychophysical functions do not superimpose across the empty and filled intervals. The functions for Group Filled are steeper in relative time than are the functions for Group Empty.

Superimposition plot of the averaged response functions for empty and filled intervals. The labels 2:8 and 4:16 refer to the anchor durations used during training. The values on the x-axis represent the signal duration divided by the point of subjective equality (PSE)
One issue of concern regarding the data and interpretation of the results is the possible effect of endpoint differences in the psychophysical functions on the reported difference in the PSEs. Accuracy at both the short and long anchor durations was lower for empty than for filled intervals. Previously published data from our lab indicated that endpoint differences are not reliably diagnostic of PSE differences between empty and filled intervals. Miki and Santi (2005) observed the same PSE difference in timing between empty and filled intervals as those reported here with anchor durations of 1 and 4 s, 2 and 8 s, and 4 and 16 s. However, in their study, for each set of anchor durations, no statistically significant difference emerged between the filled and empty intervals at the shortest or the longest signal duration. Santi et al. (2007, Exp. 2) manipulated the ambient illumination conditions during psychophysical testing. When the test condition was darkness, they found a significant difference in the PSEs for empty and filled intervals, but no statistically significant accuracy difference between empty and filled intervals at the 2- and 8-s anchor durations. When the test condition was light, they found a statistically significant endpoint accuracy difference for filled and empty intervals, but no significant difference in the PSEs. These results indicate that endpoint accuracy differences in the psychophysical functions for empty and filled intervals are neither necessary nor sufficient conditions for observing PSE differences.
Another issue of concern, related to the endpoint differences, is the potential impact of pigeons occasionally missing the presentation of a marker stimulus on the timing function for empty intervals. For example, if the pigeon misses the first marker, it may use the interval between a marker from the previous trial and the detected second marker on the current trial to make a response, which would lead to a bias to judge durations as long. On the other hand, on trials in which the first marker was missed, pigeons might not initiate timing until the second marker had occurred. Consequently, one might expect a choose-short bias on trials in which the test stimuli immediately followed the second marker (0-s delay). Santi, Ross, Coppa, and Coyle (1999) examined the effect of omitting either the first or the second marker on responding following an empty interval. Omitting the first marker on a 0-s-delay test trial did not result in a bias to judge the duration as long. In fact, it produced a statistically significant bias to judge the interval as short (M = 60 %). Omitting only the second marker on a 0-s-delay test trial resulted in no significant bias effects; accuracy remained above 80 % correct if the first visual marker was presented and only the second visual marker was omitted. Finally, if a pigeon were to miss the presentation of both markers, it might respond to the test stimuli by guessing, which would lead to a flattening of the psychophysical function. Consistent with this expectation, Grant (2001) found that omitting both markers led to 50 % accuracy and no response bias to either the short- or the long-associated response. Although missing both markers would lead to lower accuracy for both the short and long anchor durations, we have previously discussed how endpoint differences are not predictive of differences in the PSEs for empty and filled intervals.
Overall, the present results using a between-subjects design replicate previous within-subjects reports (Miki & Santi, 2005; Santi et al., 2010; Santi et al., 2007) that pigeons perceive time differently if they are required to time an empty interval (an interval demarcated by a brief marker at the start and end of the to-be-timed interval), as opposed to a filled interval (signal continuously present). That is, pigeons judge empty intervals to be longer than filled intervals. As in the previous within-subjects studies, the present between-subjects study shows that this effect is a multiplicative effect across different sets of anchor durations rather than an additive effect. In the present study, the PSE difference between empty and filled intervals doubled as the anchor durations used in training were doubled. This difference is more consistent with a difference in clock rates for empty and filled intervals than with a difference in the latencies to initiate timing. According to a switch-latency difference, the PSE difference should have remained the same, regardless of the anchor durations used in training.
The empty–filled timing difference obtained in this experiment is inconsistent with the expectation based on the mixed memory model. As we noted in the introduction, the model makes no prediction about whether a timing difference will occur for empty and filled intervals; however, it does provide an explanation of this difference in terms of a faster clock rate for empty intervals, and it suggests that in order to observe this timing difference, the accumulator values for the empty and filled intervals must be stored within a single reference memory distribution representing the short values for the two intervals and within a single reference memory distribution representing long values for the two intervals. Contrary to this expectation, the present study indicates that the empty–filled timing difference can be observed in a between-subjects design.
Interestingly, the between-subjects results in this experiment were opposite those reported by Kraemer et al. (1997), who found that pigeons judged filled intervals as being longer in duration than empty intervals. However, the nature of the empty and filled intervals was different in the Kraemer et al. (1997) work. In contrast to the present experiment, Kramer et al. (1997) signaled filled intervals by turning on a houselight for an interval of time in an otherwise dark operant chamber, and signaled empty intervals by turning off a houselight for an interval of time in an otherwise illuminated chamber. To determine whether the discrepancy with previous results was dependent on the way in which empty and filled intervals were implemented (i.e., changes in overall ambient illumination vs. localized key light markers and signals), in Experiment 2 we obtained psychophysical functions for empty and filled intervals using a procedure that was more similar to that used by Kraemer et al. (1997).
Experiment 2
Method
Subjects
Following the completion of Experiment 1, the same subjects were used in Experiment 2. Each bird remained in the same group to which it had been assigned in Experiment 1.
Apparatus
The apparatus was identical to the one used in Experiment 1.
Procedure
The procedure was identical to that of Experiment 1, except that for Group Filled the interval to be timed was signaled by the presentation of houselight illumination in an otherwise dark chamber, and for Group Empty, the interval was signaled by turning off houselight illumination in an otherwise illuminated chamber. The durations of the intervals were 4 and 16 s. All birds received a minimum of ten sessions of training, with the exception of one subject from Group Filled, who finished testing for Experiment 1 later than the other subjects, and advanced to testing after achieving the accuracy criterion following nine sessions of training. During the last three sessions of training prior to psychophysical testing, accuracy was very high and did not significantly differ between Group Empty (M = 94.45, SD = 3.42) and Group Filled (M = 90.99, SD = 5.14), t(14) = 1.59.
Psychophysical testing was conducted for 20 sessions, as was described in Experiment 1, except that the signal durations were signaled by the presence (Group Filled) or absence (Group Empty) of houselight illumination.
Results and discussion
Figure 4 displays the psychophysical functions for Group Filled and Group Empty, with the lines representing the best-fitting functions obtained from the four-parameter sigmoid equation. The functions were superimposed, indicating no timing difference between filled and empty intervals.

Mean percentages of “long” responses as a function of signal duration for filled and empty intervals following training with anchor durations of 4 and 16 s, with intervals indicated by changes in ambient illumination in Experiment 2. Error bars represent the standard errors of the means
The psychophysical functions for each pigeon were individually fit to obtain estimates of the PSE, slope, and asymptotes. The mean PSEs for Group Empty (M = 7.72, SD = 0.79) and Group Filled (M = 7.74, SD = 0.74) did not differ significantly from each other, or from the GM of 8 s. Relative to the results obtained in Experiment 1 for the 4- and 16-s anchor durations, the mean PSE appeared to shift for both groups. For Group Empty, the mean PSE was significantly greater than in Experiment 1, t(7) = 2.20, by a one-tailed (p = .032) but not by a two-tailed (p = .064) test, whereas for Group Filled, it was significantly less, t(7) = 2.91, by either test. The significant change in the PSEs suggests that pigeons judged the durations of empty and filled intervals differently relative to how these intervals had been judged in Experiment 1. Group Empty judged empty intervals signaled by houselight offset to be significantly shorter (i.e., PSE shifted to the right) than empty intervals signaled by markers. Group Filled judged filled intervals signaled by houselight illumination to be significantly longer (i.e., PSE shifted to the left) than filled intervals signaled by key light illumination. As in Experiment 1, the slopes of the psychophysical functions were equivalent for Group Empty (M = 1.71, SD = 0.29) and Group Filled (M = 1.76, SD = 0.21), t < 1. This indicates that the sensitivity to time was the same, regardless of whether the pigeons were timing filled or empty intervals. The lower asymptote for Group Empty (M = 0.06, SD = 0.16) was less than that for Group Filled (M = 2.07, SD = 5.19), whereas the upper asymptote for Group Empty (M = 93.06, SD = 3.61) was greater than that for Group Filled (M = 89.26, SD = 9.56); however, neither difference was statistically significant, ts ≤ 1.05.
These results are not consistent with the findings of Kraemer et al. (1997, Exp. 1), who found that filled intervals were judged as being longer than empty intervals, and that pigeons were more accurate when timing a filled interval rather than an empty interval.
General discussion
The purpose of conducting these experiments was to determine whether the empty–filled timing difference is dependent on the mixing of the nontemporal properties of signal durations within reference memory. That is, whether or not the detection of a clock-rate difference between empty and filled intervals is dependent on reference memory distributions that contain anchor duration values of both empty and filled intervals. Although the mixed-memory model was developed to account for timing differences due to modality, it suggests that in order to detect a difference in the clock rates with which empty and filled intervals are timed, reference memory distributions must contain values from both types of intervals. Experiment 1 produced results that were inconsistent with expectations derived from an extension of the mixed-memory model to empty–filled timing differences.
The between-group empty-filled timing difference obtained in Experiment 1 increased proportionally to the increase in anchor duration value (difference score of 0.71 vs. 1.52). The slopes of the psychophysical functions in both groups also displayed a multiplicative effect, as the slopes for both groups increased by a factor of two or greater as the anchor duration values were doubled. This multiplicative effect confirms that the empty–filled timing difference in Experiment 1 was more likely the result of pulse accumulation for empty intervals being greater than for filled intervals. The proportional increase also rules out the possibility that the pigeons timed the markers that signaled the empty intervals. If pigeons timed one or both markers along with the empty interval duration, the empty-filled timing difference should have remained constant across varying anchor durations (Miki & Santi, 2005).
Despite the use of a between-subjects design, the results of Experiment 2 of the present research were inconsistent with the results obtained by Kraemer et al. (1997). Kraemer et al. (1997) also used a between-subjects design, but they found that pigeons judged filled intervals to be longer in duration than empty intervals. The timing difference obtained by Kraemer et al. (1997) is consistent with the timing effect displayed by rats (e.g., Santi et al., 2005) and humans (Penney et al., 2000; Wearden, 1991; Wearden, Todd, & Jones, 2006). However, in pigeons, the opposite effect (i.e., the empty–filled timing difference) has been consistently obtained within-subjects (Miki & Santi, 2005; Santi et al., 2010; Santi et al., 2007). The discrepancy in the results between our Experiment 2 and Kraemer et al. (1997) are difficult to explain. In both studies, the intervals to be timed were signaled by the presence (Group Filled) or the absence (Group Empty) of ambient chamber illumination. Although there were differences in the scalings of the intermediate durations (equal log intervals in Exp. 2 vs. equal linear intervals in Kraemer et al., 1997), it is not clear how this methodological difference could have been responsible for the difference in results. Nevertheless, Experiment 2 suggests that the way in which the intervals are implemented affects the way that pigeons time the intervals. When empty intervals are signaled by visual markers, pigeons consistently show that the intervals are perceived as longer than filled intervals signaled by the continuous presentation of a discrete visual stimulus (Exp. 1 of the present study; Miki & Santi, 2005; Santi et al., 2010; Santi et al., 2007). However, empty intervals are not perceived as longer than filled intervals if they are signaled by auditory markers (Santi et al., 2007) or by changes in ambient illumination (Exp. 2 of the present study).
Although many features of the data are consistent with our explanation of differences in time perception for empty and filled intervals, this interpretation does have some shortcomings. If pigeons perceived empty intervals to be longer than filled intervals, then one would expect that the magnitude of the difference in “long” responding between empty and filled intervals would continue to increase as signal duration increased. Contrary to this expectation, “long” responding was not higher for empty intervals than for filled intervals at the long anchor duration. This feature of the data contributed to the failure of the empty and filled psychophysical functions to superimpose at the higher signal duration/PSE ratios (i.e., 1.5 and 2.0). Another shortcoming is related to our attention-sharing explanation of the empty–filled timing difference in pigeons. According to this hypothesis, empty intervals of darkness allow pigeons to devote more attention to timing the signal duration, because there would be little distracting visual information to attend to in the chamber. Although empirical tests of this hypothesis have produced supportive findings (see Santi et al., 2007), the results of Experiment 2 are not consistent with it. In Experiment 2, the empty interval was darkness, and the filled interval was chamber illumination. According to the hypothesis, the chamber illumination during a filled interval should have resulted in less attention to temporal processing, because of the tendency for visual stimuli in the chamber to capture attention. However, no difference in empty–filled timing was obtained in Experiment 2.
Penney et al. (2000) acknowledged that some studies with humans have obtained between-group modality effects, but they suggested that these studies “required participants to compare the [auditory or visual] signal to an internal representation held prior to the experiment (e.g., a clock second)” (p. 1770). More recently, Wearden et al. (2006) reported a modality timing difference in humans when auditory and visual durations were used, regardless of whether a within-group or between-group design was used. Wearden et al. (2006) also argued that the use of a common standard with which to compare the durations of an auditory or a visual stimulus could give rise to this modality effect. Although this makes sense in the context of human timing experiments using a verbal estimation procedure, it is less clear how it would apply in the case of bisection experiments. However, the between-group differences in timing behavior consistent with scalar timing theory could occur in a bisection study as a result of memory distortions for the two types of signal durations being studied (e.g., empty and filled intervals). According to scalar timing theory, clock values are transferred to reference memory according to the equation M = (1/Y) * t, where M is the reference memory value stored, Y is the memory constant, and t is the objective time (Meck, 1983). If the pulse counts for empty and filled intervals were transferred to reference memory with different memory constants (e.g., Y = 1.1 for empty intervals and Y = 0.9 for filled intervals), then the count stored for empty intervals would be lower than the value recorded by the clock, whereas the count stored for filled intervals would be greater than the value recorded by the clock. As a result, even if no difference was present in the rates of pulse accumulation for empty and filled intervals (i.e., equivalent values in the clock stage), when the clock reading was compared to the distorted value stored in reference memory, empty intervals would tend to be judged as longer, and filled intervals would tend to be judged as shorter, regardless of whether a between-group or within-group design was used. The problem for this memory distortion account is providing a good rationale for why a memory constant difference might have been present for empty and filled intervals, yet not be expressed when the empty and filled intervals were signaled by changes in the ambient illumination conditions in Experiment 2. At present, the more parsimonious explanation for our previous findings (Miki & Santi, 2005; Santi et al., 2010; Santi et al., 2007) with respect to empty–filled timing differences is competition for attention by visual features that gives rise to different pulse counts during the clock stage. However, in light of the between-group differences observed in the present study, future research should consider possible memory distortions.
In summary, the present data suggest that pigeons judge empty intervals to be longer than equal-length filled intervals, even when memory mixing of both interval types is not possible. Increasing the anchor duration values resulted in a multiplicative effect, providing strong evidence for a difference in pulse accumulation between filled and empty intervals. The results of Experiment 2 indicate that the ways in which empty and filled intervals are implemented in the procedure impact how pigeons judge the intervals’ duration.
References
Abel, S. M. (1972a). Discrimination of temporal gaps. Journal of the Acoustical Society of America, 52, 519–524. doi:10.1121/1.1913139
Abel, S. M. (1972b). Duration discrimination of noise and tone bursts. Journal of the Acoustical Society of America, 51, 1219–1223. doi:10.1121/1.1912963
Allan, L. G. (1979). The perception of time. Perception & Psychophysics, 26, 340–354. doi:10.3758/BF03204158
Allan, L. G. (1992). The internal clock revisited. In F. Macar, V. Pouthas, & W. Friedman (Eds.), Time, action and cognition: Towards bridging the gap (pp. 191–202). Dordrecht, The Netherlands: Kluwer.
Allan, L. G., & Gibbon, J. (1991). Human bisection at the geometric mean. Learning and Motivation, 22, 39–58. doi:10.1016/0023-9690(91)90016-2
Block, R. A., & Zakay, D. (1997). Prospective and retrospective duration judgments: A meta-analytic review. Psychonomic Bulletin & Review, 4, 184–197. doi:10.3758/BF03209393
Buhusi, C. V., & Meck, W. H. (2006a). Interval timing with gaps and distracters: Evaluation of the ambiguity, switch, and time-sharing hypotheses. Journal of Experimental Psychology: Animal Behavior Processes, 32, 329–338. doi:10.1037/0097-7403.32.3.329
Buhusi, C. V., & Meck, W. H. (2006b). Time sharing in rats: A peak interval procedure with gaps and distracters. Behavioural Processes, 71, 107–115. doi:10.1016/j.beproc.2005.11.017
Buhusi, C. V., Pascalis, J. P. G., & Cerutti, D. G. (2006). Time sharing in pigeons: Independent effects of gap duration, position and discriminability from the timed signal. Behavioural Processes, 71, 116–125. doi:10.1016/j.beproc.2005.10.006
Chaston, A., & Kingstone, A. (2004). Time estimation: The effect of cortically mediated attention. Brain and Cognition, 55, 286–289. doi:10.1016/j.bandc.2004.02.013
Church, R. M., & Deluty, M. Z. (1977). Bisection of temporal intervals. Journal of Experimental Psychology: Animal Behavior Processes, 3, 216–228. doi:10.1037/0097-7403.3.3.216
Craig, J. C. (1973). Constant error in the perception of brief temporal intervals. Perception & Psychophysics, 13, 99–104. doi:10.3758/BF03207241
Fraisse, P. (1978). Time and rhythm perception. In E. Carterette & M. Friedman (Eds.), Handbook of perception (Vol. 8, pp. 203–254). New York, NY: Academic Press.
Gallistel, C. R. (1990). The organization of learning. Cambridge, MA: The MIT Press.
Gibbon, J., Church, R. M., & Meck, W. H. (1984). Scalar timing in memory. Annals of the New York Academy of Sciences, 423, 52–77. doi:10.1111/j.1749-6632.1984.tb23417.x
Goldfarb, J. L., & Goldstone, S. (1963). Time judgment: A comparison of filled and unfilled durations. Perceptual and Motor Skills, 16, 376. doi:10.2466/pms.1963.16.2.376
Goldstone, S., & Goldfarb, J. L. (1963). Judgment of filled and unfilled durations: Intersensory factors. Perceptual and Motor Skills, 17, 763–774. doi:10.2466/pms.1963.17.3.763
Goldstone, S., & Lhamon, W. T. (1974). Studies of the auditory–visual differences in human time judgments: 1. Sounds are judged longer than lights. Perceptual and Motor Skills, 39, 63–82. doi:10.2466/pms.1974.39.1.63
Grant, D. S. (2001). Memory for empty time intervals. Animal Learning & Behavior, 29, 293–301. doi:10.3758/BF03192896
Grant, D. S., & Talarico, D. C. (2004). Processing of empty and filled time intervals in pigeons. Learning & Behavior, 32, 477–490. doi:10.3758/BF03196043
Grondin, S. (1993). Duration discrimination of empty and filled intervals marked by auditory and visual signals. Perception & Psychophysics, 54, 383–394. doi:10.3758/BF03205274
Grondin, S. (2001). From physical time to the first and second moments of psychological time. Psychological Bulletin, 127, 22–44. doi:10.1037/0033-2909.127.1.22
Grondin, S. (2003). Sensory modalities and temporal processing. In H. Helfrich (Ed.), Time and mind II: Information processing perspectives (pp. 61–77). Cambridge, MA: Hogrefe & Huber.
Grondin, S. (2008). Methods for studying psychological time. In S. Grondin (Ed.), Psychology of time (pp. 51–74). Bingley, UK: Emerald.
Kraemer, P. J., Brown, R. W., & Randall, C. K. (1995). Signal intensity and duration estimation in rats. Behavioural Processes, 34, 265–268. doi:10.1016/0376-6357(95)00003-D
Kraemer, P. J., Randall, C. K., & Brown, R. W. (1997). The influence of stimulus attributes on duration of matching-to-sample in pigeons. Animal Learning & Behavior, 25, 148–157. doi:10.3758/BF03199052
Lejeune, H., Macar, F., & Zakay, D. (1999). Attention and timing: dual task performance in pigeons. Behavioural Processes, 45, 141–157. doi:10.1016/S0376-6357(99)00015-7
Meck, W. H. (1983). Selective adjustment of the speed of internal clock and memory processes. Journal of Experimental Psychology: Animal Behavior Processes, 9, 171–201. doi:10.1037/0097-7403.9.2.171
Meck, W. H. (1984). Attentional bias between modalities: Effect on the internal clock, memory, and decision stages used in animal time discrimination. In J. Gibbon & L. Allan (Eds.), Annals of the New York Academy of Sciences: Timing and time perception (Vol. 423, pp. 528–545). New York, NY: New York Academy of Sciences.
Meck, W. H. (1991). Modality-specific circadian rhythmicities influence mechanisms of attention and memory for interval timing. Learning and Motivation, 22, 153–179. doi:10.1016/0023-9690(91)90021-Y
Miki, A., & Santi, A. (2005). The perception of empty and filled time intervals by pigeons. Quarterly Journal of Experimental Psychology, 58B, 31–45. doi:10.1080/0272499044000032
Molet, M., Alessandri, J., & Zentall, T. R. (2011). Subjective time: Cognitive and physical secondary tasks affect timing differently. Quarterly Journal of Experimental Psychology, 64, 1344–1353. doi:10.1080/17470218.2011.552728
Penney, T. B., Gibbon, J., & Meck, W. H. (2000). Differential effects of auditory and visual signals on clock speed and temporal memory. Journal of Experimental Psychology: Human Perception and Performance, 26, 1770–1787. doi:10.1037/0096-1523.26.6.1770
Penney, T. B., Gibbon, J., & Meck, W. H. (2008). Categorical scaling of duration bisection in pigeons (Columba livia), mice (Mus musculus), and humans (Homo sapiens). Psychological Science, 19, 1103–1109. doi:10.1111/j.1467-9280.2008.02210.x
Poynter, W. D. (1989). Judging the duration of time intervals: A process of remembering segments of experience. In I. Levin & D. Zakay (Eds.), Time and human cognition: A life-span perspective (pp. 305–321). Amsterdam, The Netherlands: Elsevier.
Rammsayer, T. H. (2010). Differences in duration discrimination of filled and empty auditory intervals as a function of base duration. Attention, Perception, & Psychophysics, 72, 1591–1600. doi:10.3758/APP.72.6.1591
Rammsayer, T. H., & Lima, S. D. (1991). Duration discrimination of filled and empty auditory intervals: Cognitive and perceptual factors. Perception & Psychophysics, 50, 565–574. doi:10.3758/BF03207541
Rammsayer, T. H., & Skrandies, W. (1998). Stimulus characteristics and temporal information processing: Psychophysical and electrophysiological data. Journal of Psychophysiology, 12, 1–12.
Roberts, W. A., Cheng, K., & Cohen, J. S. (1989). Timing light and tone signals in pigeons. Journal of Experimental Psychology: Animal Behavior Processes, 15, 23–35. doi:10.1037/0097-7403.15.1.23
Santi, A., Adams, A., & Bassett, J. (2010). The role of keypecking during filled intervals on the judgment of time for empty and filled intervals by pigeons. Learning & Behavior, 38, 42–49. doi:10.3758/LB.38.1.42
Santi, A., Keough, D., Gagne, S., & Van Rooyen, P. (2007). Differential effects of empty and filled intervals on duration estimation by pigeons: Tests of an attention-sharing explanation. Behavioural Processes, 74, 176–186. doi:10.1016/j.beproc.2006.08.008
Santi, A., Miki, A., Hornyak, S., & Eidse, J. (2005). The perception of empty and filled time intervals by rats. Behavioural Processes, 70, 247–263. doi:10.1016/j.beproc.2005.07.006
Santi, A., Ross, L., Coppa, R., & Coyle, J. (1999). Pigeons’ memory for empty time intevals marked by visual or auditory stimuli. Animal Learning & Behavior, 27, 190–205. doi:10.3758/BF03199675
Stubbs, D. A., Dreyfus, L. R., & Fetterman, J. G. (1984). The perception of temporal events. In J. Gibbon & L. Allan (Eds.), Timing and time perception (pp. 30–42). New York, NY: New York Academy of Sciences.
Sutton, J. E., & Roberts, W. A. (2002). The effect of nontemporal information processing on time estimation in pigeons. Learning and Motivation, 33, 124–140. doi:10.1006/lmot.2001.1103
Wearden, J. H. (1991). Human performance on an analogue of an interval bisection task. Quarterly Journal of Experimental Psychology, 43B, 59–81. doi:10.1080/14640749108401259
Wearden, J. H., & Ferrara, A. (1995). Stimulus spacing effects in temporal bisection by humans. Quarterly Journal of Experimental Psychology, 48B, 289–310. doi:10.1080/14640749508401454
Wearden, J. H., & Ferrara, A. (1996). Stimulus range effects in temporal bisection by humans. Quarterly Journal of Experimental Psychology, 49B, 24–44. doi:10.1080/713932615
Wearden, J. H., Rogers, P., & Thomas, R. (1997). Temporal bisection in humans with longer stimulus durations. Quarterly Journal of Experimental Psychology, 50B, 79–94. doi:10.1080/713932643
Wearden, J. H., Todd, N. P. M., & Jones, L. A. (2006). When do auditory/visual differences in duration judgments occur? Quarterly Journal of Experimental Psychology, 59, 1709–1724. doi:10.1080/17470210600784649
Wilkie, D. M. (1987). Stimulus intensity affects pigeons timing behavior: Implications for an internal clock model. Animal Learning & Behavior, 15, 35–39. doi:10.3758/BF03204901
Zakay, D., & Block, R. A. (1996). The role of attention in time estimation processes. In M. A. Pastor & J. Artieda (Eds.), Time, internal clocks, and movement (pp. 143–164). Amsterdam, The Netherlands: Elsevier.
Zakay, D., & Block, R. A. (1997). Temporal cognition. Current Directions in Psychological Science, 6, 12–16. doi:10.1111/1467-8721.ep11512604
Zentall, T. R., & Singer, R. (2008). Required pecking and refraining from pecking refraining from pecking alters judgments of time by pigeons. Learning & Behavior, 36, 55–61. doi:10.3758/LB.36.1.55
Author note
This research was supported by Grant No. OGPOOD6378 from the Natural Sciences and Engineering Research Council of Canada to A. S. The authors thank Kelley Putzu and Tammy Buitenhuis for their animal care assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Price, E., Santi, A. The effect of filled and empty intervals on clock and memory processes in pigeons. Learn Behav 42, 153–163 (2014). https://doi.org/10.3758/s13420-014-0134-1
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13420-014-0134-1