
Abstract
The whitefly species, Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae) is an important pest of many vegetable crops in the world. To evaluate the potential of the predatory species, Orius albidipennis Reuter (Hemiptera: Anthocoridae) in pest control, functional responses of the predator to different densities of eggs and third instar nymphs of the whitefly were determined under the laboratory conditions. Densities of (5, 8, 10, 15, 20, 25, 30, and 35) eggs and/or third instar nymphs per cucumber leaf disc were exposed to 1-day-old adults of the predator for 24-h period. The logistic regression analysis and Roger’s model were used for determination of type and parameters of the predator’s functional responses, respectively. The results showed that O. albidipennis exhibited type II and III functional responses, when fed on the whitefly eggs and third instar nymphs, respectively. The handling times (Th) of O. albidipennis were calculated to be 0. 35 h and 1. 01 h, when it was fed on eggs and third instar nymphs of B. tabaci. The attack rate value (a) of the bug were calculated as 0.0723 h−1 on eggs, and the constant (b) was 0.0069, when it preyed on the third instar nymphs of the whitefly. The maximum attack rate (T/Th) was calculated as 68.39 eggs and 23.20 third instar nymphs. It could be concluded that O. albidipennis was efficient in controlling B. tabaci. However, further studies in the greenhouse and field conditions are needed.
Similar content being viewed by others

Background
The tobacco whitefly Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) is a key pest of numerous host plant species found in tropical and warm temperate regions of the world (Stansly and Natwick 2010; Stansly et al. 2010). The host plants may directly experience injury from sap removal, honeydew excretion that creates favorite conditions for the rapid growth of sooty mold, which interferes with photosynthesis and also causes reduction in the product quality and acceptability, as well as physiological disorder (Stansly and Natwick 2010; Stansly et al. 2010). The whitefly also causes indirect damage by virus transmission. For control of the pest, chemical insecticides have usually been applied by farmers, but the problems associated with chemical pesticides, including rapid development of insecticide resistance, side effect on non-target organisms, secondary pest outbreaks, environmental contaminations, and adverse effects on human health have led to the development of integrated pest management (IPM) (Pedigo 2002). Biological control has been a valuable component in IPM programs around the world for many years (Orr 2009).
Predatory bugs, belonging to the genus Orius (Hemiptera: Anthocoridae), are generalist predators of various soft bodied arthropods, including whiteflies (Reitz et al. 2006). Orius albidipennis Reuter is a common predator in several regions of Iran, and its potential as a biocontrol agent has been reported, especially under greenhouse conditions (Salehi et al. 2016). It is crucial to evaluate efficiency of natural enemies before using them in a biological control program (Fathipour et al. 2006). One of the criteria for evaluating a predator efficacy is its feeding response to changes in prey species, density, or its functional response (Holling 1965). Functional responses of Orius bugs to some pests were determined by previous researchers. For instance, functional responses of O. insidiousus Say to Panonychus ulmi Koch (McCaffrey and Horsburgh 1986), Frankliniella occidentalis Pergande (Coll and Ridgway 1995), and Aphis glycines Matsumura (Rutledge and O’Neil 2005) were studied. But, no effort has been done to determine functional response of O. albidipennis to B. tabaci.
Therefore, the present study aimed to estimate the functional response of the adult bugs to eggs and third instar nymphs of B. tabaci under the laboratory conditions.
Materials and methods
Host plants
Cucumber (Cucumis sativus cv. Servis plus) seeds were grown in plastic pots (8-cm diameter and 13-cm height) filled with a soil-cocopeat mix (1:1, v:v) and irrigated every 3 days. The pots were maintained in insect cages (120 × 100 × 60 cm) inside a controlled environment room (25 ± 5 °C, 16 L:8D). After 5–6 weeks from cultivation, the host plants were used for establishment of the whitefly colony and for trials.
Insect rearing
Adults and nymphs of B. tabaci were collected from cucumber fields in Mollasani region, Ahwaz, Khuzestan province, southwest of Iran. The samples were introduced into rearing cages (120× 100 × 60 cm) containing the cucumber pots. The pots were kept inside a controlled environment room (25 ± 1 °C, 65 ± 5% RH and a photoperiod of 16 L:8D). After the establishment of the colony, eggs and third instar nymphs were used in the experiments. To obtain the cohorts, five couples of whitefly adults (each included one male and one female) were collected from the rearing stock and confined on host plant leaves by a clips cage (2 .5-cm diameter). The adults were removed after 24 h. The eggs and third instar nymphs were obtained for the experimentation after 1 and 10 days, respectively.
Adults of O. albidipennis were collected from corn and sunflower fields in Mollasani region, Khuzestan province, southwest of Iran. The female bugs were isolated in a Plexiglas cylinder (18 cm high, 7 .5cm diameter). The top of lid cylinder was covered by a fine net for ventilation as well as the holes placed on its surface. The nymphs and adults of the predatory bug were fed on frozen Ephestia kuehniella Zeller (Lepidoptera: Pyralidae) eggs and pollens of corn and sunflower. The bean pods, Phaseulus vulgaris L., were used as oviposition substrates. The predator was reared at (25 ± 1 °C, 65 ± 5% RH, and 16:8 h (L:D)) in an environmental controlled room. For reduction of cannibalism, the rearing cylinders were lined by crumpled wipe papers.
Experimental design
The experimental arena was a Petri dish (10-cm diameter). A hole (2-cm diameter) was constructed in the fine gauze cover lid for ventilation. Inside each Petri dish, a cucumber leaf disc (5-cm diameter) was placed upside down on a wetted filter paper to keep the leaf discs fresh (Montserrat et al. 2000). Densities of (5, 8, 10, 15, 20, 25, 30, and 35) the preys (eggs or third instar nymphs of B. tabaci) per arena were used. The densities were determined according to preliminary tests. A female predator (1 to 5 days old), kept starved for 24 h before the experiment, was introduced to each arena. Following 24 h of exposure, the predatory bugs were removed and the numbers of consumed eggs and nymphs were recorded, separately. The trials were performed at (25 ± 1 °C, 65 ± 5% RH, and a photoperiod of 16:8 h (L: D)) in an environmentally controlled room. Ten replicates were conducted at each prey density.
Data analyses
Analysis of variance (ANOVA) by SAS 9.1 was used to compare consumed preys among different prey densities. The type of the functional response of predator was determined by logistic regression analysis of the proportion of consumed preys (Na/N0), as a function of prey offered (N0) (Juliano 2001). Data were fitted to a polynomial function that describes the relationship between Ne/N0 and N0 (Eq. 1):
where Na is the number of consumed prey, N0 is the initial number of preys, and P0, P1, P2, and P3 are the intercept of the linear, quadratic, and cubic coefficients, respectively. These parameters were calculated, using the method of maximum likelihood (PROC CATMOD, SAS Institute, 2001). When P1 > 0 and P2 < 0, the proportion of consumed preys is positively density-dependent—the data describes a type III of functional response, but if P1 < 0 and P2 > 0, the proportion of consuming preys decreases gradually as the initial number of prey offered increases—the data indicates a type II of functional response (Juliano 2001). In the second step of the analysis, nonlinear least squares regression (PROC NLIN; SAS Institute Inc. 2001) was used to fit Roger’s random attack model, which describes a type II or III functional response (Eq 2 and Eq 3, respectively), for estimation of the functional response parameters, respectively. Because prey density was reduced during the trial, this model, which does not assume a constant prey density, is a suitable choice for the analysis (Juliano 2001).
where Na is the number of consumed prey, N0 is the initial number of prey, a is the instantaneous attack rate (searching efficiency), T is the total amount time available for searching (in this experiment T = 24 h), Pt the number of predators, and Th is the handling time.
where b, c, and d are constants from the function that relates the attack coefficient (a) and N0 in the type III functional response. The attack coefficient (a) is calculated by Eq 4 (Hassell 1978).
The curves of the number of consumed prey by O. albidipennis females to different densities of the eggs and third instar nymphs of B. tabaci were depicted by Excel software.
Results and discussion
The results showed that at densities of 5–25 eggs or nymphs per arena, the consumed prey significantly increased with increasing prey density. But at densities more than 25 eggs or nymphs per arena, the number of consumed prey was not significantly different (Table 1).
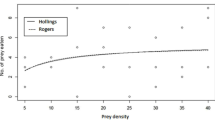
The obtained data from the logistic regression analysis of the functional response of the females of O. albidipennis implicated that the predatory bug showed the type II and III functional responses, when fed on the whitefly eggs and third instar nymphs, respectively (Table 1). Functional response curves of female adults of O. albidipennis to the various densities of eggs and third instar nymphs of B. tabaci are presented in (Figs. 1 and 2), respectively. The number of consumed eggs or nymphs by the bug increased with increasing prey density (Figs. 1 and 2).

Mean number of Bemisia tabaci eggs preyed upon by females of Orius albidipennis in 24 h as a function of egg density

Mean number of Bemisia tabaci third instar nymphs preyed upon by females of Orius albidipennis in 24 h as a function of nymph density
In many cases, Orius spp. exhibited type II functional response (Holing, 1965); however, type III functional response shows the ability of predators to increase their rate of attack with increasing in prey density (Hassell 1978), which means that the predator learned how to overcome some hurdles with catching the prey (Schenk and Bacher 2002). Type II functional response was previously reported for O. albidipennis, when preyed on Megalurothrips sjostedti Tymbom larvae (Gitonga et al. 2002), T. tabaci (Thysanoptera: Thripidae) (Madadi et al. 2007), and Tetranychus turkestani Ugarov & Nikolski (Acari: Tetranychidae) (Hassanzadeh et al. 2015). However, the predatory bug showed type III functional response to Tuta absoluta Meyreck (Lepidoptera: Gelechiidae) (Salehi et al. 2016). In contrast to the present results, type II functional responses of 4 heteropteran predators, Dicyphus tamaninii Wagner and Macrolophus caliginosus Wagner (Het., Miridae), Orius majusculus Reuter and Orius laevigatus Fieber (Het., Anthocoridae), to Trialeurodes vaporariorum Westwood (Homoptera: Aleyrodidae) nymphs were determined (Montserrat et al. 2000). Similar to the obtained data, M. caliginosus bug exhibited type III functional response to nymphs of T. vaporariorum (Enkegaard et al. 2001). Obtained results indicated that types and estimated parameters of the functional response of the predatory bug were different, when it was fed on eggs or nymphs of B. tabaci. This variation may be related to the differences in size. Size of prey is an important factor that determines the type and parameters of a predator functional response (Hassell et al. 1977 and Streams 1994). These findings are in line with the results of Gitonga et al. (2002) who demonstrated that the O. albidipennis showed different types and parameters of functional response, according to its prey life stages. Functional response models often assume the homogenous prey size and size-independent feeding rates. However, variation in prey size due to ontogeny and competition is ubiquitous, and predation rates are often size dependent (McCoy et al. 2011).
Estimated functional response parameters, when the bug preys on eggs or third instar nymphs of the pest, are shown in Table 2. The handling times (Th) of O. albidipennis were calculated to be 0.350 h and 1.034 h, when fed on eggs and third instar nymphs of B. tabaci, respectively. The attack rate value (a) of the bug was calculated as 0.0723 h−1 on eggs, and the constant (b) was 0.0069, when it preyed on the third instar nymphs of the whitefly. The maximum attack rate (T/Th) was calculated as 68.39 eggs and 23.20 third instar nymphs (Table 2). The calculated values of Th and a in the present study were different from the values reported for O. majuscules and O. laevigatus to T. vaporarium (Montserrat et al. 2000). The difference may be due to different prey and predator species.
Conclusion
Determination of the functional responses of O. albidipennis under laboratory conditions, using simple arenas and a single prey type, may be of a limited value to predict entire predator’s behavior under field or greenhouse conditions. Therefore, further studies in the greenhouse and field conditions are needed.
References
Coll M, Ridgway RL (1995) Functional and numerical responses of Orius insidiosus (Heteroptera: Anthocoridae) to its prey in different vegetable crops. Ann Entom Soc Am 88(6):732–738
Enkegaard A, Brodesgaard H, Hansen DL (2001) Macrolophus caliginosus: functional response to whiteflies and preference and switching capacity between whiteflies and spider mites. Entomol -Exp Appl 101(1):81–88
Fathipour Y, Hosseini A, Talebi A, Moharramipour S (2006) Functional response and mutual interference of Diaeretiella rapae (Hymenoptera: Aphidiidae) on Brevicoryne brassicae (Homoptera: Aphididae). Entomol Fen 17:90–97
Gitonga LM, Overholt WA, Lohr JK, Magambo JK, Mucke JM (2002) Functional response of Orius albidipennis (Hemiptera: Anthocoridae) to Megalurothrips sjostedti (Thysanoptera: Thripidae). Biol Control 24:1–6
Hassanzadeh H, Esfandiari M, Shishehbor P, Rajabpour A (2015) Functional response of different developmental stages of Orius albidipennis (Reuter) (Hem.: Anthocoridae) feeding on the strawberry spider mite, Tetranychus turkestani (Ugarov & Nikolski) (Acari: Tetranychidae). -Plant Protec. 38(3): 63–74. (in Persian)
Hassell MP (1978) The dynamics of arthropod predator-prey system. Princeton University Press, US, p 248
Hassell MP, Lawton JH, Beddington JR (1977) Sigmoid functional responses by invertebrate predators and parasitoids. J Anim Ecol 46:249–262
Holling CS (1965) The functional response of predators to prey density and its role in mimicry and population regulation. Entomol Soc Canada 45:1–60
Juliano SA (2001) Non-linear curve fitting: predation and functional response curves. In: Scheiner SM, Gurevitch J (eds) Design and analysis of ecological experiments, 2nd edn. Chapman and Hall, New York, pp 178–196
Madadi H, Enkegaard A, Bodsgaard HF, Kharazi-Pakdel A, Mohaghegh J, Ashouri A (2007) Host plant effects on the functional response of Neoseiulus cucumeris to onion thrips larvae. J Appl Entomol 131(9–10):728–733
McCaffrey JP, Horsburgh RL (1986) Functional response of Orius insidiosus (Hemiptera: Anthocoridae) to the European red mite, Panonychus ulmi (Acari: Tetranychidae), at different constant temperatures. Environ Entomol 15(3):532–535
McCoy MV, Bolker BM, Warkentin KM, Vonesh JR (2011) Predicting predation through prey ontogeny using size-dependent functional response models. -Amer. Nat. 177(6):752–766
Montserrat M, Albajes R, Castane C (2000) Functional response of four heteropteran predators preying on greenhouse whitefly (Homoptera: Aleyrodidae) and western flower Thrips (Thysanoptera: Thripidae). Environ Entomol 29(5):1075–1082
Orr D (2009) Biological control and integrated pest management. In: Peshin R, Dhawan AK (eds) Integrated pest management. Innovation-Development Process. Springer, Netherland, pp 504–527
Pedigo LP (2002) Entomology and pest management. Iowa University press, Iowa, US, p 420
Reitz SR, Funderburk JE, Waring SM (2006) Differential predation by the generalist predator Orius insidiosus on congeneric species of thrips that vary in size and behaviour. Entomol Exp Appl 119:179–188
Rutledge CE, O’Neil RJ (2005) Orius insidiosus (Say) as a predator of the soybean aphid, Aphis glycines Matsumura. Biol Control 33(1):56–64
Salehi Z, Yarahmadi F, Rasekh A, Zandi Sohani N (2016): Functional responses of Orius albidipennis Reuter (Hemiptera, Anthocoridae) to Tuta absoluta Meyrick (Lepidoptera, Gelechiidae) on two tomato cultivars with different leaf morphological characteristics. Entomol Gen 36(2): 127–136
Schenk D, Bacher S (2002) Functional response of a generalist predator to one of its prey species in the field. J Anim Ecol 71:524–531
Stansly PA, Natwick ET (2010) Integrated system foir managing Bemisia tabaci in protected and open field agriculture. In: Stanly PA, Naranjo SE (eds) Bemisia: Bionomics and management of a global pest. Springer Science+ Business Media B.V, pp 467–497
Stansly PA, Steven N, Legg JP (2010) Biology and ecology of Bemisia tabaci. In: Stansly PA, Steven N (eds) Bemisia: Bionomics and management of a global pest. Springer Science+ Business Media B.V, p 533
Streams FA (1994) Effect of prey size on attack component of the functional response by Notonecta undulate. Oecologia 98(1):57–63
Acknowledgements
The research was supported by Agricultural Sciences and Natural Resources University of Khuzestan [grant number: 9228409].
Funding
The research was supported by grant from Agricultural Sciences and Natural Resources University of Khuzestan [9228409].
Availability of data and materials
Not applicable
Author information
Authors and Affiliations
Contributions
FY and NZS designed the experiments. AS performed the experiments. FY and NZS analyzed the data. FY wrote the manuscript. The manuscript studied and certificated by all authors. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Shahpouri, A., Yarahmadi, F. & Zandi Sohani, N. Functional response of the predatory species Orius albidipennis Reuter (Hemiptera: Anthocoridae) to two life stages of Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae). Egypt J Biol Pest Control 29, 14 (2019). https://doi.org/10.1186/s41938-019-0119-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-019-0119-7