
Abstract
This study aimed to isolate and characterize bacteria from the codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae), and determine their virulence to its larvae. A total of 16 bacteria were isolated from larvae belonging to different instars. Based on morphological, biochemical, physiological, and molecular studies, the bacterial isolates were identified as Pseudomonas sp. (Cp1, 3, 5, and 13), Corynebacterium sp. (Cp2), Bacillus sp. (Cp4, 7, 9, 10, 12, and 15), Pectobacterium carotovorum (Cp6), Paenibacillus sp. (Cp8), Bacillus megaterium (Cp11), Bacillus pumilus (Cp14), and Terribacillus saccharophilus (Cp16). It was also determined the virulence of these isolates, where the highest potential activity was obtained by Bacillus sp. Cp9, with (76%) mortality. These results could be beneficial for future biocontrol programs of C. pomonella.
Similar content being viewed by others


Background
The codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae), is one of the most important pests in many orchards worldwide, mainly apples, pears, quince, peach, plum, apricot, and walnut. It causes economic losses in fruit production (Pajac et al. 2012 and Alford 2014). The larvae of this pest which overwinter in the cracked bark of tree trunks and in cocoons at packaging and storage places develop to pupae in late April to early May. The mated female moths emerging from pupae lay eggs at appropriate temperatures. The hatched larvae burrow into the fruit within 4–8 h and render them unsalable (Beers et al. 2003). In order to control this pest and to obtain undamaged fruits, traditional insecticides such as organophosphorus compounds and synthetic pyrethroids have been used. However, these insecticides cause unfavorable environmental impacts. In addition, some strains of this pest have acquired resistance to several insecticides (Lacey and Unruh 2005).
Entomopathogenic microorganisms such as bacteria, viruses, fungi, nematodes, and protists are able to infect different insect species, and they can be used as biological control agents against insect pests (Khetan 2001). Among entomopathogens, the entomopathogenic bacteria (EPB) play a key role in the commercial control of insect pests and Bacillus thuringiensis (Bt) is the species on which most of the scientific community and industry efforts have been focused (Owuama 2001 and Ruiu et al. 2013). Apart from Bt, many different EPB belonging to different species of Bacillus and other genera, such as Bacillus sphaericus, Paenibacillus papillae, and Serratia entomophila, are available as insecticides (Federici 2007).
Many insect life cycles are associated with symbiotic microorganisms, and there is increasing evidence that symbiotic microorganisms influence many insect features such as sex determination, nutrient exchange, nutrition, and digestion processes (Rajagopal 2009; Douglas 2014; and Brune 2014). However, some insect groups are not obligatory dependent on their microbiota (Douglas 2014). Symbiotic microorganisms, especially bacteria, can be used in the biological control of insect pests through the use of different methods, e.g., they can be used to express insecticidal toxins or proteins by using genetic engineering techniques (Beard et al. 1998). In addition, changing the dynamics among bacterial microbes in the insect gut could be used for controlling insect pests. For different purposes, the microbiota of many insect species has been determined (Sevim et al. 2012; Demirci et al. 2013; and Roopa et al. 2014).
This study aimed to isolate and characterize bacterial species from C. pomonella. Additionally, these bacterial isolates were tested against the larvae of the codling moth.
Methods
Collection of larvae
Different larval instars of C. pomonella were collected from infested walnut fruits at the vicinity of Kırşehir, Turkey, in the summer of 2015. The obtained larvae were separated according to the developmental stage (instars) and used in the process of bacterial isolation.
Isolation of bacteria
The collected larvae were divided into three groups based on their instars. The first group consisted of the first and second larval instars, the second group consisted of the third instar, and the third group consisted of the fourth and fifth larval instars. The bacterial isolation was separately performed from these groups. A total of ten larvae were used for each group for the bacterial isolation. The larvae were surface-sterilized with 70% ethanol for 2–3 min and washed three times with sterile distilled water (Lipa and Wiland 1972). Thereafter, the larvae belonging to the different groups were separately placed into glass test tubes (10 ml) including 3 ml nutrient broth (Difco, NJ, USA) with sterile forceps and completely homogenized, using a sterile glass tissue grinder. The homogenates were filtered through two layers of sterile cheesecloth to remove insect debris. A series of dilutions from 10−1 to 10−8 were prepared from the insect homogenates, and 10−1, 10−3, 10−5, and 10−8 dilutions from each homogenate were plated on nutrient agar and then incubated at 30 °C for 3 days. In addition, these dilutions were heated at 80 °C for 10 min to eliminate non-spore-forming bacteria. Then, they were plated on nutrient agar and incubated at 30 °C for 3 days; then, the bacterial colonies were counted, and the total number of bacteria per larvae was calculated as 2 × 106 cfu (colony forming units). Moreover, the different bacterial colonies were streaked on nutrient agar and incubated at 30 °C for 18 and 48 h for slow-growing isolates to obtain pure cultures. The obtained pure cultures were stored in 20% glycerol at − 20 °C. The bacterial isolates were identified based on various tests. All isolates from this study are publicly accessible and were deposited at Microbiology Laboratory, Genetic and Bioengineering, Ahi Evran University, Kırşehir, Turkey.
Morphological characterization of the bacterial isolates
The bacterial isolates were morphologically characterized on the basis of their colony, cell, and spore features. Colony morphologies of the bacterial isolates were evaluated on nutrient agar plates by using a stereomicroscope (Demirci et al. 2013). Cell properties of the isolates were evaluated by the gram and endospore staining. The capsule layer of the bacterial isolates was determined by negative staining. The motility of the isolates was determined according to the method of Soutourina et al. (2001).
Physiological characterization of the bacterial isolates
The bacterial isolates were also physiologically characterized on the basis of their growth at different temperatures, NaCl concentrations, and pH. All isolates were inoculated into nutrient broth (3 ml) and incubated at different temperatures ranging from 4 to 55 °C. Also, all isolates were incubated into nutrient broth (3 ml) with different concentrations of NaCl, ranging from 3 to 15%. Finally, all isolates were incubated into nutrient broth (3 ml) with different pH values ranging from 3 to 12. Evaluations were visually made.
VITEK 2 microbial identification system
The bacterial isolates were also identified using the VITEK 2 microbial identification system. Firstly, the bacterial isolates were streaked on nutrient agar plates to obtain single colonies. The bacterial suspensions were prepared from a single colony, using 2 ml of 0.45% sterile saline solution to the equivalent of a 0.5 McFarland turbidity standard. Concentrations were checked with the VITEK colorimeter for each isolate. Additionally, the oxidase and catalase production of the isolates were manually determined. A total of two cards were used to identify the isolates. The GN ID card was used for gram-negative bacterial identification and the GP ID card was used for gram-positive bacterial identification. The bacterial suspensions prepared as above were inoculated onto these cards and incubated at 30 °C for 18 h. The time between preparation of the suspension and card filling was less than half an hour. The results were automatically evaluated with the VITEK 2 device (Ligozzi et al. 2002).
16S rRNA gene sequencing
The bacterial isolates were further characterized, using the partial sequencing of 16S rRNA gene. Genomic DNAs were extracted by the Genomic DNA isolation kit (Thermo Fisher Scientific, Waltham, MA, USA). The extracted DNAs were stored at − 20 °C until PCR was done.
Approximately 1.450 bp of the 16S rRNA gene region was targeted and amplified. The primer pairs of 27F (5′-AGAGTTTGATCMTGGCTCAG-3′ as forward) and 1492R (5′-GGYTACCTTGTTACGACTT-3′ as reverse) were purchased from MACROGEN and used for amplification. The total volume of PCR reactions was 50 μl to which 50–100 ng genomic DNA was added. The PCR mix of 50 μl per sample contained 25 pmol of each primer, 200 mM each of the dNTPs, 1×PCR buffer, 3 mM MgCl2 and 1.5 U Taq DNA polymerase. After adding all the components, the final volume was adjusted to 50 μl with sterile distilled water. The PCR program consisted of 95 °C (60 s) for the initial denaturation, followed by 35 cycles of 94 °C (45 s) for denaturation, 55 °C (30 s) for annealing, 72 °C (1.5 min) for extension, and a final extension of 72 °C (5 min). After performing PCR, 5 ml of the products was analyzed by electrophoresis on 1.0% agarose gel containing ethidium bromide to check the sizes and amounts of the amplicons. After checking PCR products, the accurate products were sent to MACROGEN (the Netherlands) for sequencing. The PCR products were sequenced with the primer pairs 518F (5′-CCAGCAGCCGCGGTAATACG-3′) and 800R (5′-TACCAGGGTATCTAATCC-3′). The obtained sequences were subjected to the nucleotide BLAST searches in the NCBI GenBank database to get the percentage similarity of the bacterial isolates to the most related bacterial species (Altschul et al. 1990).
Phylogenetic analysis
Phylogenetic analysis of the bacterial isolates and their closely related species was performed for molecular characterization of the bacterial isolates. The sequences were edited using Bioedit, and multiple sequence alignments were created by using 16S rRNA sequences belonging to our strains and different bacterial species from the NCBI GenBank database for the purpose of developing a phylogenetic tree. The multiple sequence alignment was performed with ClustalW in Bioedit (Hall 1999). Finally, the sequences were subjected to neighbor-joining analysis with p-distance correction, gap omission, and 1.000 bootstrap pseudoreplicates using MEGA 6.0 (Tamura et al. 2013).
Nucleotide sequence accession numbers
The GenBank accession numbers of the 16S rRNA gene sequences belonging to the bacterial isolates from this study are listed in Table 4.
Bioassay
Each bacterium isolated from C. pomonella in the stock culture was streaked on nutrient agar to obtain a single colony and to check the purity of the cultures. After that, 3 ml of nutrient broth was inoculated from each single colony of 16 bacteria and incubated at 30 °C overnight. At the end of the incubation period, the bacterial density was measured at 600 nm absorbance and adjusted to 1.8 × 109 cfu/ml by centrifugation (4.000 rpm for 15 min) and using sterile phosphate buffer solution (PBS) (Moar et al. 1995). The bacterial solutions were freshly prepared and used for bioassay.
For the bioassay experiments, healthy C. pomonella larvae were obtained from the laboratory culture at Ahi Evran University, Genetic Bioengineering and Microbiology Laboratory. Healthy larvae were randomly selected and used for the bioassay. Twenty-five grams of freshly prepared artificial diet (for diet ingredients and rearing conditions (Fukova et al. 2005)) was inoculated with 1 ml of the bacterial suspension prepared as described above for each isolate. For the control group, 25 g of the artificial diet had 1 ml of the sterile PBS added. The contaminated artificial diets were separately placed into plastic boxes (20 × 10 × 8 mm) with ventilated lids to permit airflow. After that, ten third instar C. pomonella larvae were placed into the box for each replicate and allowed to feed on the contaminated diets. A total of ten larvae were used for each replicate, and all experiments were repeated three times. Finally, the plastic boxes were incubated at 25 °C under 16:8 (day:night) light regime. After 10 days, the boxes were checked for larval mortality, and the number of dead larvae was recorded. Mortality data were corrected based on Abbott’s formula (Abbott 1925). To determine the differences among the isolates and the control group, the data were subjected to ANOVA and subsequently to the LSD multiple comparison test. Before performing ANOVA, all data were tested for homogeneity of variance using Levene’s statistic. All tests were performed with SPSS 16.0 statistical software.
Results and discussion
A total of 16 isolates of bacteria were obtained from the treated C. pomonella larvae. Among these isolates, five were from the first and second larval instars, eight from the third larval instar, and three from the fourth and fifth larval instars. The isolates were characterized on the basis of their morphological, physiological, and molecular characteristics. Colonies of all isolates were smooth, except for Cp9 and Cp11, which were rough. Only one isolate (Cp12) had a mucoid colony. Three isolates (Cp3, Cp5, and Cp13) had yellow colonies, and one isolate (Cp4) had a pink colony. The other isolates produced creamy-colored colonies. Five isolates (Cp1, Cp3, Cp5, Cp6, and Cp13) were gram-negative, and the others were gram-positive. All isolates were bacilli-shaped, except for Cp2 and Cp3 which were coccus shaped. It was found that six isolates (Cp7, Cp8, Cp9, Cp10, Cp14, and Cp16) formed spores. Only two isolates (Cp1 and Cp13) had capsules. All morphological characteristics of the bacterial isolates are given in Table 1.
All isolates were able to grow in 3% NaCl, and only one isolate (Cp3) could not grow in 4% NaCl. Growth characteristics of the isolates in other NaCl concentrations were variable, depending on the isolate. All isolates were able to grow in the pH range of 3, 4, and 5, except for Cp12. All isolates were able to grow at pH 6 and 7. Growth characteristics of the isolates at other pHs were variable, depending on the isolate. None of the isolates grew at pH 4 and 55 °C, and growth properties of the isolates at 30, 37, 45, and 50 °C were variable, depending on the isolate. Physiological properties of the isolates are given in Table 2. The VITEK 2 microbial characterization system for biochemical characterization of the isolates and gram-negative and gram-positive cards was used to identify them. Five isolates (Cp6, Cp12, Cp14, Cp15, and Cp16) were not able to be characterized by VITEK 2. Other identifications are given in Table 3.
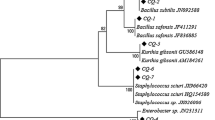
The bacterial isolates were also characterized on the basis of 16S rRNA gene sequencing to verify the recorded conventional characterizations of the isolates. Based on molecular characterization, the bacterial isolates were identified as Pseudomonas sp. (Cp1, 3, 5, and 13), Corynebacterium sp. (Cp2), Bacillus sp. (Cp4, 7, 9, 10, 12, and 15), Pectobacterium carotovorum (Cp6), Paenibacillus sp. (Cp8), Bacillus megaterium (Cp11), Bacillus pumilus (Cp14), and Terribacillus saccharophilus (Cp16) (Table 4). This identification was also supported by phylogenetic analysis (Fig. 1).

Phylogenetic tree derived from neighbor-joining analysis of 16S rRNA sequences (1400 bp) from the flora members of C. pomonella and their closely related species. Bootstrap values based on 1000 replicates were indicated above nodes. Bootstrap values C ≥ 70 are labeled. C. pomonella isolates were indicated with black circle. The scale on the bottom of the dendrogram indicates the degree of dissimilarity
All isolates caused different mortality values in comparison to each other (F = 15.43, df = 16, p < 0.05). The highest mortality values were obtained from Bacillus sp. Cp4, Cp9, and Cp10 with 70, 76, and 63%, respectively (F = 15.43, df = 16, p < 0.05). Other mortalities ranged from 3 to 56% (Fig. 2). Ertürk and Demirbağ (2006) studied the ability of culturing a bacterial flora of C. pomonella. They obtained eight bacterial isolates from the larvae of this pest, collected from apple fruits. Also, the bacterial flora were Proteus rettgeri (Cp1), Eschericia coli (Cp2), Pseudomonas stutzeri (Cp3), Pseudomonas aeroginosa (Cp4), Bacillus laterosporus (Cp5), Micrococcus sp. (Cp6), Proteus vulgaris (Cp7), and Deinococcus sp. (Cp8). However, in this study, 16 bacterial isolates from the same insect collected from walnut fruits were obtained. The microbiota of insects was affected by many factors such as diet, development stage, habitat, and phylogeny of the host (Yun et al. 2014). The difference between these studies with respect to the bacterial diversity might be due to the use of different diets of C. pomonella larvae.

Virulence of the bacterial isolates using the bacterial concentration of 1.89 × 109 cfu/ml against C. pomonella larvae within 10 days after application. Mortality data were corrected according to Abbott’s formula (Abbott 1925). Bars indicate standard deviation. Different lowercase letters represent statistically significant differences among larval mortalities. Cp1 Pseudomonas sp., Cp2 Corynebacterium sp., Cp3 Pseudomonas sp., Cp4 Bacillus sp., Cp5 Pseudomonas sp., Cp6 Pectobacterium carotovorum, Cp7 Bacillus sp., Cp8 Paenibacillus, Cp9 Bacillus sp., Cp10 Bacillus sp., Cp11 B. megaterium, Cp12 Bacillus sp., Cp13 Pseudomonas sp., Cp14 B. pumilus, Cp15 Bacillus sp., and Cp16 Terribacillus saccharophilus
Among the EPB, spore-forming bacilli are the major group of species of bacteria that infect and kill insects (Aronson et al. 1986). Many different Bacillus species have been isolated from different insects which are harmful in both agriculture and forestry. In the present study, eight different Bacillus species were obtained and characterized, and some of them, namely Cp4, Cp9, and Cp10, showed a high virulence against larvae of C. pomonella.
The genus Pseudomonas contains 191 currently described species (Euzeby 1997). Some of them are entomopathogenic such as Pseudomonas aeruginosa, Pseudomonas fluorescens, Pseudomonas putida, Pseudomonas entomophila, and Pseudomonas taiwanensis (Khetan 2001; Mahar et al. 2005; Chen et al. 2014; and Dieppois et al. 2014). In the present study, four isolated Pseudomonas species were not characterized at the species level. Among them, Pseudomonas sp. Cp1 showed insecticidal activity against the larvae of the codling moth. This may suggest that probably a new Pseudomonas species might be isolated from the codling moth. However, more detailed identification studies should be conducted to verify this probability.
The genus Paenibacillus includes bacteria which are facultative anaerobic and endospore-forming. This genus was previously included in the Bacillus genus but was reclassified as a separate genus (Ash et al. 1993). The members of this genus, which have been isolated from various environments such as soil, rhizosphere, water, clinical samples, and insects, are becoming important in agricultural and medical applications (McSpadden Gardener 2004; Lal and Tabacchioni 2009; and Danismazoglu et al. 2012). This genus includes some insect pathogenic bacteria such as Paenibacillus larvae, Paenibacillus popilliae, and Paenibacillus lentimorbus (Ruiu 2015). In the present study, one Paenibacillus sp. (Cp8) from live larva was isolated. However, it did not show any insecticidal activity against the larvae of the codling moth.
The genus Corynebacterium contains gram-positive, aerobic, and rod-shaped bacteria. The members of this genus are widespread in nature and have been isolated from different human and animal habitats (Collins et al. 2004). Some non-pathogenic members of this genus are also intensely used in industrial applications such as the production of amino acids, bioconversion of steroids, degradation of hydrocarbons, and cheese aging (Yamada et al. 1972 and Lee et al. 1985). Some members of this genus have been isolated from insects (Bucher 1981 and Hoeven et al. 2008). In this study, a Corynebacterium sp. (Cp2) was isolated from the codling moth, but it had no insecticidal activity against the larvae.
The genus Pectobacterium (formerly known as Erwinia) is a member of the family Enterobacteriaceae, and some species have been isolated from different environments such as soil, water, plants, and invertebrates (Ian et al. 2003 and Glasner et al. 2008). Some species within this genus, such as P. carotovorum, is an important plant pathogen of many vegetable plants such as tomato, potato, and carrot (Ma et al. 2007). Some studies showed an isolation of the members of this genus from insects (Gnanamanickam 2006). In the present study, the species (Cp6) was also isolated, but it had no any insecticidal activity.
Terribacillus is a genus of the family Bacillaceae that contains species that are aerobic, spore-forming, gram-positive, rod-shaped, and halophilic (An et al. 2007). The members of this genus have been isolated from various environments such as soil and saline lake sediments (An et al. 2007 and Liu et al. 2010). Some studies showed an association of some species of this genus with invertebrates (Menezes et al. 2010 and Vicente et al. 2013). In this study, also one Terribacillus species (T. saccharophilus Cp16) was isolated but it had no any insecticidal activity.
Conclusions
The aerobic gut bacteria of C. pomonella were isolated and characterized searching for bacterial control agents which may be used against it. Some of the flora members (Cp1, Cp4, Cp9, and Cp10) showed significant insecticidal activity under laboratory conditions, especially Bacillus sp. Cp9 that showed promising results against larvae of the pest. Further studies are still needed to determine the efficacy of this isolate under field conditions. Mass production and formulation studies are also warranted.
References
Abbott WS (1925) A method of computing the effectiveness of an insecticide. J Econ Entomol 18:265–267
Alford DV (2014) Pests of fruit crops: a color handbook, 2nd edn. CRC press, Boca Raton, Florida
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215:403–410
An SY, Asahara M, Goto K, Kasai H, Yokota A (2007) Terribacillus saccharophilus gen. nov., sp. nov. and Terribacillus halophilus sp. nov. spore-forming bacteria isolated from field soil in Japan. Int J Syst Evol Microbiol 57:51–55
Aronson AI, Beckman W, Dunn P (1986) Bacillus thuringiensis and related insect pathogens. Microbiol Rev 50(1):1–24
Ash C, Priest FG, Collins MD (1993) Molecular identification of rRNA group 3 bacilli (Ash, Farrow, Wallbanks and Collins) using a PCR probe test. Ant van Leeuwen 64:253–260
Beard CB, Durvasula RV, Richards FF (1998) Bacterial symbiosis in arthropods and the control of disease transmission. Emerg Infect Dis 4:581–591
Beers EH, Suckling DM, Prokopy RJ, Avilla J (2003) Ecology and management of apple arthropod pests. In: Ferree DC, Warrington IJ (eds) Apples: botany, production and uses. CABI publishing, Oxon, pp 489–520
Brune A (2014) Symbiotic digestion of lignocellulose in termite guts. Nat Rev Microbiol 12:168–180
Bucher C (1981) Identification of bacteria found in insects. In: Burges HD (ed) Microbial control of pests and plant diseases. Academic Press, New York, pp 7–33
Chen WJ, Hsieh FC, Hsu FC, Tasy YF, Liu JR, Shih MC (2014) Characterization of an insecticidal toxin and pathogenicity of Pseudomonas taiwanensis against insects. PLoS Pathog 10(8):e1004288
Collins MD, Hoyles L, Foster G, Falsen E (2004) Corynebacterium caspium sp. nov., from a Caspian seal (Phoca caspica). Int J Syst Evol Microbiol 54:925–928
Danismazoglu M, Demir I, Sevim A, Demirbag Z, Nalcaciolglu R (2012) An investigation on the bacterial flora of Agriotes lineatus (Coleoptera: Elateridae) and pathogenicity of the flora members. Crop Proctec 40:1–7
Demirci M, Sevim E, Demir İ, Sevim A (2013) Culturable bacterial microbiota of Plagioderaversicolora (L.) (Coleoptera: Chrysomelidae) and virulence of the isolated strains. Folia Microbiol 58:201–210
Dieppois G, Opota O, Laculat J, Lemaitre B (2014) Pseudomonas entomophila: a versatile bacterium with entomopathogenic properties. In: Ramos JL, Goldberg JB, Filloux A (eds) Pseudomonas, vol 7: new aspects of Pseudomonas biology. Springer, Dordrecht, pp 25–49
Douglas AE (2014) The molecular basis of bacterial–insect symbiosis. J Mol Biol 426:3830–3837
Ertürk Ö, Demirbağ Z (2006) Studies on bacterial flora and biological control agent of Cydiapomonella L. (Lepidoptera: Tortricidae). Afr J Biotechnol 5(22):2081–2085
Euzeby JP (1997) List of bacterial names with standing in nomenclature: a folder available on the internet. Int J Syst Bacteriol 47(2):590–592
Federici BA (2007) Bacteria as biological control agents for insects: economics, engineering, and environmental safety. In: Vurro M, Gressel J (eds) Novel biotechnologies for biocontrol agent enhancement and management. Springer, Dordrecht, pp 25–51
Fukova I, Nguyen P, Marec F (2005) Codling moth cytogenetics: karyotype, chromosomal location of rDNA, and molecular differentiation of sex chromosomes. Genome 48:1083–1092
Glasner JD, Marquez-Villavicencio M, Kim HS, Jahn CE, Ma B, Biehl BS, Rissman AI, Mole B, Yi X, Yang CH, Dangl JL, Grant SR, Perna NT, Charkowski AO (2008) Niche-specificity and the variable fraction of the Pectobacterium pan-genome. Mol Plant-Microbe Interact 21:1549–1560
Gnanamanickam SS (2006) Plant-associated bacteria. Springer, Dordrecht
Hall TA (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp 41:95–98
Hoeven RVD, Betrabet G, Forst S (2008) Characterization of the gut bacterial community in Manduca sexta and effect of antibiotics on bacterial diversity and nematode reproduction. FEMS Microbiol Lett 286:249–256
Ian KT, Kenneth SB, Holeva CM, RJP B (2003) Soft rot erwiniae: from genes to genomes. Mol Plant Pathol 4(1):17–30
Khetan SK (2001) Microbial Pest Control. Marcel Dekker, New York
Lacey LA, Unruh TR (2005) Biological control of codling moth (Cydia pomonella, Lepidoptera: Tortricidae) and its role in integrated pest management, with emphasis on entomopathogens. Vedalia 12(1):33–60
Lal S, Tabacchioni S (2009) Ecology and biotechnological potential of Paenibacillus polymyxa: a minireview. Indian J Microbiol 49:2–10
Lee CW, Lucas S, Desomazeaud MJ (1985) Phenylalanine and tyrosine catabolism in some cheese coryneform bacteria. FEMS Microbiol Lett 26(2):201–205
Ligozzi M, Bernini C, Bonora MG, de Fatima M, Zuliani J, Fontana R (2002) Evaluation of the VITEK 2 system for identification and antimicrobial susceptibility testing of medically relevant gram-positive cocci. J Clin Microbiol 40(5):1681–1686
Lipa JJ, Wiland E (1972) Bacteria isolated from cutworms and their infectivity to Agrotis sp. Acta Microbiol Pol 4:127–140
Liu W, Jiang L, Guo C, Yang SS (2010) Terribacillus aidingensis sp. nov., a moderately halophilic bacterium. Int J Syst Evol Microbiol 60:2940–2945
Ma B, Hibbing ME, Kim HS, Reedy RM, Yedidia I, Breuer J, Breuer J, Glasner JD, Perna T, Kelman A, Charkowski AO (2007) Host range and molecular phylogenies of the soft rot enterobacterial genera Pectobacterium and Dickeya. Phytopathology 97(9):1150–1163
Mahar AN, Darban DA, Gowen SR, Hague NGM, Jan ND, Munir M, Mahar AQ (2005) Use of entomopathogenic bacterium Pseudomonas putida (Enterobacteriaceae) and its secretion against greater wax moth, Galleria mellonella pupae. J Entomol 2(1):77–85
McSpadden Gardener BB (2004) Ecology of Bacillus and Paenibacillus spp. in agricultural systems. Phytopathology 94:1252–1258
Menezes CB, Bonugli-Santos RC, Miqueletto PB, Passarini MR, Silva CH, Justo MR, Leal RR, Fantinatti-Garboggini F, Oliveira VM, Berlinck RG, Sette LD (2010) Microbial diversity associated with algae, ascidians and sponges from the north coast of São Paulo state, Brazil. Microbiol Res 165:466–482
Moar WJ, Pusztzai-Carey M, Mack TP (1995) Toxicity of purified proteins and the HD-1 strain from Bacillus thuringiensis against lesser cornstalk borer (Lepidoptera: Pyralidae). J Econ Entomol 88:606–609
Owuama CI (2001) Entomopathogenic symbiotic bacteria, Xenorhabdus and Phororhabdus of nematodes. World J Microbiol Biotechnol 17:505–515
Pajac I, Baric B, Mikac K, Pejic I (2012) New insights into the biology and ecology of Cydia pomonella from apple orchards in Croatia. Bull Insectol 65(2):185–193
Rajagopal R (2009) Beneficial interactions between insects and gut bacteria. Indian J Microbiol 49:114–119
Roopa HK, Rebijith KB, Asokan R, Mahmood R, Kumar K (2014) Isolation and identification of culturable bacteria from honeydew of whitefly, Bemisia tabaci (G.) (Hemiptera: Aleyrodidae). Meta Gene 2:114–122
Ruiu L (2015) Insect pathogenic bacteria in integrated pest management. Insects 6:352–367
Ruiu L, Satta A, Floris I (2013) Emerging entomopathogenic bacteria for insect pest management. Bull Insectol 66(2):181–186
Sevim E, Çelebi Ö, Sevim A (2012) Determination of the bacterial flora as a microbial control agent of Toxoptera aurantii (Homoptera: Aphididae). Biologia 67(2):397–404
Soutourina OA, Semenova EA, Parfenova VV, Danchin A, Bertin P (2001) Control of bacterial motility by environmental factors in polarly flagellated and peritrichous bacteria isolated from Lake Baikal. Appl Environ Microbiol 67:3852–3859
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30:2725–2729
Vicente CSL, Nascimento FX, Espada M, Barbosa P, Hasegawa K, Mota M, Oliveira S (2013) Characterization of bacterial communities associated with the pine sawyer beetle Monochamus galloprovincialis, the insect vector of the pinewood nematode Bursaphelenchus xylophilus. FEMS Microbiol Lett 347:130–139
Yamada K, Kinoshita S, Tsunoda T, Aida K (1972) The microbial production of amino acids. Wiley, New York
Yun JH, Roh SW, Whon TW, Jung MJ, Kim MS, Park DS, Yoon C, Nam YD, Kim YJ, Choi JH, Kim JY, Shin NR, Kim SH, Lee WJ, Bae JW (2014) Insect gut bacterial diversity determined by environmental habitat, diet, developmental stage, and phylogeny of host. Appl Environ Microbiol 80(17):5254–5264
Acknowledgements
We would like to thank Dr. Frantisek Marec for providing C. pomonella eggs.
Funding
This study was supported by Ahi Evran University Scientific Research Projects Coordination Unit, project number: PYO-MÜH.4001.15.008.
Availability of data and materials
Not applicable.
Author information
Authors and Affiliations
Contributions
ES carried out a large part of the whole experiments. MÇ collected the insect specimens in the field and participated in the bacterial isolation experiments. FMS carried out the VITEK-2 bacterial identification experiments. AS participated in the 16S rRNA gene sequencing, phylogenetic analysis, statistical analysis, and writing of the whole manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
None of the authors have any competing interests in the manuscript.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Sevim, E., Çocar, M., Sezgin, F.M. et al. Aerobic gut bacterial flora of Cydia pomonella (L.) (Lepidoptera: Tortricidae) and their virulence to the host. Egypt J Biol Pest Control 28, 30 (2018). https://doi.org/10.1186/s41938-018-0036-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-018-0036-1