
Abstract
Background
Evolution of the penis in sexual method of reproduction had showed complex developmental processes, leading to extreme species variation. The presence and absence of the os penis vary in taxa within mammals indicate adaptive value of the mammalian penis bone, and understanding the diversity is challenging. Surprisingly, little is known about its details.
Methods
Comparative analysis of the penis with special reference to the os penis (penile baculum) was analyzed in monkey (Macaca radiata), rat (Rattus norvegicus), and bat (Pipistrellus mimus).
Results
In monkey, the baculum appears like a miniature of a long bone, a clear central medullary cavity, and surrounded by osteons. Similarly, the baculum in rat showed a medullary canal and surrounded by smaller channels of osteons. In bat, it is seated over the corpus spongiosum that shows a small diameter channel which forms the main lamellar bone and appears to be a single osteon or Haversian system. Though microanatomical examination revealed that the presence of vascular and non-vascular structures contribute for penile erection, the presence of the os penis would be helpful for the penis erection and stiffness and vaginal friction.
Conclusions
Anatomical relationship to other penile elements and its specific functional significance and the explanation for this general adaptive diversity across species is challenging.
Similar content being viewed by others

Background
The penis is a copulatory organ designed to deliver sperm into the female reproductive tract, apart from eliminating urine. The penis in mammals is supported by a pair of hydrostatic skeletal elements: the corpus cavernosum and the corpus spongiosum (Dean and Lue 2005). The corpus cavernosum is a high-pressure system designed to increase the flexural stiffness of the penis during erection and to maintain its shape during copulation. The corpus spongiosum is a low-pressure system that inflates the glans during erection and supports its tissue during copulation (Sattar et al. 1995). Apart from this, additional non-vascular supportive erectile structures like (the baculum or os penis, preputial gland) help in penile erection, the mammalian baculum, or the os penis, is extremely morphologically diverse both in shape and size across species (Romer 1970; Ramm 2007; Stockley et al. 2013; Simmons and Firman 2014; Klaczko et al. 2015). Early records of observations of the os penis or baculum in mammals go back to Aristotle. Prosthetic surgery for treatment of impotence was born from the initial experience with the use of the “artificial os penis” (rib cartilage) in post-traumatic reconstruction in 1936 (Bretan 1989), with many research targeted to study the erectile dysfunction and impotency using animal models. Surprisingly, our understanding of the penile morphology and baculum are lacking in many species (Goran 1997; Klaczko et al. 2015; Schultz et al. 2016). In this paper, macro and microanatomical features of three eutherian penises were discussed. Animal species analyzed were monkey (Macaca radiata), rat (Rattus norvegicus), and bat (Pipistrellus mimus).
Methods
Animals used for this presented study were not procured exclusively for this investigation. All the study were done using corpse stored in the department tissue bank (from projects approved by institutional ethics committee). All the three set of animals used were devoid of deformity especially in the perineal region and scrotal content. The penile tissues were procured from monkey (Macaca radiata), rat (Rattus norvegicus), and bat (Pipistrellus mimus). Details are as follows: adult male monkeys of body weight ranging from 6 to 8 kg were selected for the study (n = 5). Adult male rats of body weight ranging from 200 to 250 g were selected for the study (n = 5). Our institute is prone for bat colonies, during a span of 5 years, and came across with three dead adult male bats (after tangled in mess grilled window or partition in the animal house area); the penis of these animals was analyzed, before they were discarded.
Radiological, morphological, and histological studies
The penile tissues collected from these mammals were subjected to radiological analysis. The X-ray images of the lateral view of the penis were taken. The organs were photographed and various morphological measurements like volume, weight, length, and width were noted using venire calipers. Volume was measured by water displacement method, and weight was measured using analytical balance. Penile tissues were fixed using formalin and processed for paraffin embedding, and sections were taken at 7 to 10 μm thickness. Sections were stained with hematoxylin (Harris’s) and counterstained using eosin.
Results
Morphological and histological studies
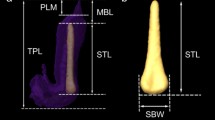
The X-ray images showed the presence of fibrous skeletal structure, the baculum, or os penis in monkey and rat. With more of cartilaginous substance, the X-ray images failed to produce X-ray shadow of the os penis in the bat. The preputial glands in monkey and bat were not prominent or rudimentary, but in the case of rat, it was very prominent (images not shown). In bonnet monkey, the prepuce is always retracted in a position exposing the glans penis and it is triangular or button-shaped, details were given elsewhere (Prakash et al. 2009). Histological observation indicates a shaft with a medullary cavity in these animals. A small diameter of this channel in the bat is the main lamellar bone with a single osteon or Haversian system with erectile tissue on either side. In rat and monkey, multiple lamellar patterns surround the medullary cavity. Histological (H&E stain), morphological, and radiological images of the baculum are shown in Fig. 1, and morphological measurements of the baculum and penis are presented in Table 1.

Photomicrograph (H&E staining) of the penile transverse section taken from the monkey (a), rat (b), and bat (c). First column: In the monkey, the baculum appears like a miniature of a long bone, a clear central medullary cavity was seen (arrow head) and surrounded minor channels. Similar in rat baculum (arrow), a medullary canal (arrow head) is surrounded by smaller canals. The baculum in the bat (arrow) is seated over the corpus spongiosum (CS) with corpus cavernosum (CC) along its lateral aspect (bilaterally), arrowhead pointing urethral orifice in the corpus spongiosum. Small diameter channel in the bat is the main lamellar bone which appears as a single osteon or Haversian system). Second column: Dissected and isolated baculum from monkey and rat with corresponding radiological images placed inside (baculum—indicated by an arrow)
Discussion
Darwin proposed sexual selection to explain why in some species the males have such magnificent ornaments and in some species, such impressive weapons and females choose particularly ornate males (female choice) or accept the winner of males (male-male competition). Darwin describes female choice as accepting “not the male which is most attractive to her, but the one which is least distasteful” (Weihe 1987; Harcourt and Gardiner 1994; Harcourt 1995). Thus, secondary sexual characteristics (larger size, large antlers, and brightly colored plumage) appear to provide a breeding advantage. Similarly, the primary sexual characteristics of the external genitalia must also provide a reproductive advantage for each species. It is likely to provide support during prolonged intromissions, female genital stimulation, and/or sperm transport facilitation and delivery (Monica and Scalici 2017).
Penile morphology in these animals indicates crucial role in the success of sexual reproduction across the animal kingdom. Studies suggest that the evolution of these complex penile morphologies has been influenced by female choice (Darwin 1971; Diamond 1997; Short 1981). The penile morphology may contribute to improved sperm competition and adaptation (Prakash et al. 2014). There is extensive morphological and functional diversity in the penis associated with these structures; this includes the baculum (os penis), preputial glands, penile spines, penile horny papillae, periurethral lappets, paired corpora cavernosum, ornate glans penis, and absent glans penis (Cold and Tarara 1997).
The presence of the baculum in mammals (excluding human) seems to have enormous influence on sexual behavior. The presence of the baculum in these species could facilitate rapid erection and sustainability (Dixson 2012; Herdina et al. 2015). Thus, they may be helpful for the penis erection and stiffness, and vaginal friction (Kelly 2000). All the three eutherian mammals, the monkey, rat, and bat, have baculum in the penis (except few taxa in bats, vide infra). The sexual behavior observed in the monkey (Gunst et al. 2013) and in the rat (Suresh and Prakash 2012) has shown multiple mounting and ejaculation in these animals. Apart from presumably functional explanations for the baculum like facilitating sperm transfer, it may stimulate the cervix, inducing ovulation in some species (Sachs et al. 1984; Parag et al. 2006). However, the phylogenetic distribution is generally lacking (Dixson 1987, 1995) and baculum length is found to scale only weakly with male body mass (Ramm 2007).
Though bats are one of the largest mammalian groups with an os penis, however, there are few taxa where the baculum seems to be absent. Literature indicates that the baculum either completely absent in the Phyllostomidae and Noctilionidae families or absent in some species within the families Mormoopidae, Vespertilionidae, Molossidae, and Pteropodidae, and no report on Chiroptera (Williams-Ashman 1990; Herdina et al. 2010). This clearly indicates no relationships between genital morphology and sexual selection across mammalian taxa. A number of species studied for the baculum appears to be fewer; moreover, there seems to be a difference in shape and length of the baculum among bat species. The white-striped mastiff bat has an unusual short trifid baculum. The Beccari’s mastiff bat and the little mastiff bat have a rod-like relatively longer baculum with a slightly spatulate tip (Krutsch and Crichton 1987).
The os penis in the rat and mouse are from the proximal and distal segments at glans penis (Vilmann 1982; Vilmann and Vilmann 1983; Rasmussen et al. 1986; Murakami 1987; Yamamoto 1987). In rats, the os penis is well developed. These results suggest that postcopulatory sexual selection influences genital length in at least some mammals. However, significant questions that remain are as to why selection on the baculum should differ between mammalian groups and the exact mechanistic basis through which males benefit from increase genital length or other possible functions are highly debatable (Hamilton 1936, 1946; Hooper 1958; Lidicker 1968; McClintock et al. 1982; Williams-Ashman 1990).
In general, the os penis is located from the dorsal to the urethra and medial to the corpora cavernosa in the penile shaft, and their distal end often variably extends into the glans. Wide degrees of variation observed in the shape of the baculum, usually rod-like, but more complex structural patterns can be seen across taxas. For example, the distal end of the os penis in rat shows comb-like contours with terminal bent, serrated projections that protrude out of the skin of the glans. A similar structure was also reported in some squirrel, voles, and rice rats (Patterson and Thaeler 1982; Patterson 1983).
The baculum in the rat (Rattus norvegicus) has been suggested as a load-bearing structure that can transmit impeding force during mounting between the baculum and the corpus cavernosum and thereby can raise intracavernosal pressure (Kelly 2000). It has been noted that this is a very good rodent experimental model in a study involving mating behavior and penile reflex analysis (Suresh and Prakash 2011; Suresh and Prakash 2012). Further studies to establish the role of the baculum on corpus cavernosum stiffness would be interesting.
Many suggestive functions of the baculum have proposed such as support and maintenance of penile erections, vaginal penetration, and enhancing friction of the vaginal wall, which might elicit neuroendocrine in the female tract. Development and growth of the baculum are controlled by androgens similar to other penile components (Murakami and Mizuno 1986). An experimental study involving androgen upregulation in neonate females showed baculum-like structures indicating androgen-dependent development. However, these changes in the baculum are not expected in adult rat during the administration of androgens (Glucksmann and Cherry 1972; Yoshida and Huggins 1980). More interestingly, the recent findings suggest that not only androgen but also locally aromatized estrogen is involved in the early growth and development of the baculum (Yonezawa et al. 2011).
In M. radiata, the baculum is well developed and excellently supporting the glans penis, especially during erection. Part of our study indicates that the presence of spines in glans penis could facilitate friction and increase vaginal secretion, which may be essential for the formation and maintenance of vaginal plug essential to combat the spells of dry environment seen even in favorable breeding months (Prakash et al. 2009) in the southern part of India.
The helmet-shaped glans penis is virtually identical in M. fuscata, M. mulatta, M. cyclopis, and M. fascicularis and extends dorsally and ventrally along the shaft of the penis (Fooden 1971). No such extensions in M. radiata in our observation (Prakash et al. 2009). Similar to all other primates (with the exception of human, brachyteles, and langothrix), the penis of this monkey contains a baculum. This is assumed to represent atavism or the appearance of a characteristic found in remote ancestors but not in nearer ancestors (Hoeg 1986). The baculum in M. fuscata, M. mulatta, and M. fascicularis is of similar form; these animals constitute a closely interrelated group based on penile characteristics. But the baculum in M. fascicularis is smaller than in M. fuscata and M. mulatta (Fooden 1971). Surprisingly, the baculum in M. radiata is longer than in M. fuscata and M. mulatta, which may play a major role in M. radiata copulation. Preliminary observation on M. radiata copulatory behavior in captivity was similar to that described for stump-tail macaques (M. arctoides) (Brereton 1994).
Present observations do indicate that there is positive relation seen with the baculum and glans penis length in rodents but no relationship with relative testis mass. In contrast, there is no evidence to support an association between baculum length and relative testis mass in either bats or primates (Ramm 2007). Suggesting that the influence of genital length at least in some mammals as a postcopulatory sexual selection. This study tried to explain some of the aspects of the penis and baculum in these mammals; however, it seems that we remain generally ignorant of the function; therefore, the nature of the selection pressures on most aspects of penile morphology (Harcourt 1995).
The specific anatomical designs may provide explanations for the individual requirements for the specific physiologic functions that differ from species to species. Although there is no baculum in the human glans, a strong equivalent distal ligament is arranged centrally and acts as a supporting trunk for the glans penis (Hsu et al. 2005). In humans, rare congenital os penis has been reported in human males (Hoeg 1986; Hsu et al. 2005; Brereton 1994; Yilmaz et al. 2013) (assumed to represent atavism but no real proof) or calcification can also occur in Peyronie’s disease (etiology of the diseases not clear) signifying penile pathology occurring at a younger age (Levine et al. 2003; Kalokairinou et al. 2012). Thus, the appearance of the baculum or os penis is a pathological occurrence in human. Under this condition with its influence on penile reconstruction surgery (where surgical management is the only option) (Zaid et al. 2014), the outcome could be an interesting observation. However, the baculum in non-human primates like macaques are consistent and exhibit wide varieties in the penis structure (Dixson 1987).
Conclusions
The presence and absence of the os penis vary in taxa within mammals; thus, understanding the adaptive value of the mammalian penis bone is challenging. The specific functional basis through which males benefit from increased genital length needed further study. It is mandatory to understand the sexual physiology and anatomy of each experimental animal model and the essential differences between the models from the human condition for prospective new drug discoveries and translation (Wu and Kovac 2016). Especially, studies related to erectile function using animal models and a thought on the baculum would be a useful and interesting addition to a study.
References
Brereton, A. R. (1994). Copulatory behavior in a free-ranging population of stumptail macaques (Macaca arctoides) in Mexico. Primates, 35, 113–122.
Bretan Jr., P. N. (1989). History of the prosthetic treatment of impotence. Urol Clin North Amer, 16, 1–5.
Cold, C., & Tarara, R. (1997). Penile and clitoral prepuce mucocutaneous receptors in Macaca mulatta. Vet Pathol, 34, 506.
Darwin, C. R. (1971). The descent of man and selection in relation to sex. London: Murray.
Dean, R. C., & Lue, T. F. (2005). Physiology of penile erection and pathophysiology of erectile dysfunction. Urol Clin N Am, 32, 379-v.
Diamond, J. (1997). Why sex is fun: the evolution of human sexuality. London: Weidenfeld and Nicolson.
Dixson, A.,. F. (1995). Baculum length and copulatory behaviour in carnivores and pinnipeds (grand order ferae). J Zool (Lond), 235, 67–76.
Dixson, A. F. (1987). Observations on the evolution of the genitalia and copulatory behaviour in male primates. J Zool (Lond), 213, 423–443.
Dixson, A. F. (2012). Primate sexuality: comparative studies of the prosimians, monkeys, apes, and humans. Oxford: Oxford University Press.
Fooden, J. (1971). Male external genitalia and systematic relationships of the Japanese macaque (Macaca fuscata BLYTH, 1875). Primates, 12, 305–311.
Glucksmann, A., & Cherry, C. P. (1972). The hormonal induction of an os clitoridis in the neonatal and adult rat. J Anat, 112, 223–231.
Goran, A. (1997). The evolution of animal genitalia: distinguishing between hypotheses by single species studies. Biol J Linn Soc, 60, 365–379.
Gunst, N., Leca, L. B., & Vasey, P. (2013). Development of sexual and socio-sexual behaviours in free-ranging juvenile male Japanese macaques (Macaca fuscata). Behaviour, 150, 1225–1254.
Hamilton, W. A. (1936). A study of the baculum in the genera Perognathus and Dipodomys. J Mammal, 17, 145–156.
Hamilton, W. J. (1946). A study of the baculum in some north American Microtinae. J Mammal, 27, 378–387.
Harcourt, A. H. (1995). Sexual selection and sperm competition in primates, what are male genitalia good for? Evol Anthropol, 4, 121–129.
Harcourt, A. H., & Gardiner, J. (1994). Sexual selection and genital anatomy of male primates. Proc R Soc Lond, 255, 47–53.
Herdina, A. N., Herzig-Straschil, B., Hilgers, H., Metscher, B. D., & Plenk, H. (2010). Histomorphology of the penis bone (Baculum) in the gray long-eared bat Plecotus austriacus (Chiroptera, Vespertilionidae). Anat Rec, 293, 1248–1258.
Herdina, A. N., Kelly, D. A., Jahelková, H., Lina, P. H. C., Horáček, I., & Metscher, B. D. (2015). Testing hypotheses of bat baculum function with 3D models derived from microCT. J Anat, 226, 229–235.
Hoeg, O. M. (1986). Human penile ossification. Scand J Urol Nephrol, 20, 231–232.
Hooper, E. T. (1958). The male phallus in mice of the genus Peromyscus. Muc Pubi Mus Zool Univ Mich, 105, 1–23.
Hsu, G. L., Lin, C. W., Hsieh, C. H., Hsieh, J. T., Chen, S. C., Kuo, T. F., … Tseng, G. F. (2005). Distal ligament in human glans: a comparative study of penile architecture. J Androl, 26, 624–628.
Kalokairinou, K., Konstantinidis, C., Domazou, M., Kalogeropoulos, T., Kosmidis, P., & Gekas, A. (2012). US imaging in Peyronie’s disease. J Clini Imag Sci, 2, 63.
Kelly, D. A. (2000). Anatomy of the baculum–corpus cavernosum interface in the Norway rat (Rattus norvegicus), and implications for force transfer during copulation. J Morphol, 244, 69–77.
Klaczko, J., Ingram, T., & Losos, J. (2015). Genitals evolve faster than other traits in Anolis lizards. J Zool, 295, 44–48.
Krutsch, P. H., & Crichton, E. G. (1987). Reproductive biology of the male little mastiff bat Mormopterus planiceps (Chiroptera: Molossidae), in Southeast Australia. Am J Anat, 178, 352–368.
Levine, L. A., Estrada, C. R., Storm, D. W., & Matkov, T. G. (2003). Peyronie disease in younger men characteristics and treatment results. J Androl, 24, 27–32.
Lidicker, W. Z. (1968). A phylogeny of New Guinea rodent genera based on phallic morphology. J Mammal, 49, 609–643.
McClintock, M. K., Anisko, J. J., & Adler, N. T. (1982). Group mating among Norway rats. II. The social dynamics of copulations: competition, cooperation and mate choice. Anim Behav, 30, 410–425.
Monica, C., & Scalici, M. (2017). Baculum (Os penis). In The International Encyclopedia of Primatology. John Wiley & Sons, Inc.. https://doi.org/10.1002/9781119179313.wbprim0214.
Murakami, R. (1987). A histological study of the development of the penis of wild – type and androgen intensive mice. J Anat, 153, 223–231.
Murakami, R., & Mizuno, T. (1986). Proximal-distal sequence of development of the skeletal tissues in the penis of rat and the inductive effect of epithelium. Embryol Exp Morphol, 92, 133–143.
Parag, A., Bennett, N. C., Faulkes, C. G., & Bateman, P. W. (2006). Penile morphology of African mole rats (Bathyergidae): structural modification in relation to mode of ovulation and degree of sociality. J Zool, 270, 323–329.
Patterson, B. D. (1983). Baculum-body size relationships as evidence for a selective continuum on baculum morphology. J Mammal, 64, 496–499.
Patterson, B. D., & Thaeler, C. S. (1982). The mammalian baculum: hypotheses on the nature of baculum variability. J Mammal, 63, 1–15.
Prakash, S., Prithiviraj, E., Suresh, S., Lakshmi, N. V., Ganesh, M. K., Anuradha, M., … Dinesh, P. (2014). Morphological diversity of sperm: a mini review. Iran J Reprod Med, 12, 239–242.
Prakash, S., Suresh, S., & Prithiviraj, E. (2009). Anatomical aspects of bonnet monkey (Macaca radiata) male reproductive system. Anat Sci Int, 84, 53–60.
Ramm, S. A. (2007). Sexual selection and genital evolution in mammals: a phylogenetic analysis of Baculum length. Am Nat, 19, 169.
Rasmussen, K. K., Vilmann, H., & Juhl, M. (1986). Os penis of the rat. V. - the distal cartilage process. Acta Anat, 125, 208–212.
Romer, A. S. (1970). The vertebrate body. Philadelphia: Saunders.
Sachs, B. D., Glater, G. B., & O’Hanlon, J. K. (1984). Morphology of the erect glans penis under various gonadal hormone conditions. Anat Rec, 210, 45–52.
Sattar, A. A., Salpigides, G., Vanderhaeghen, J. J., Schulman, C. C., & Wespes, E. (1995). Cavernous oxygen tension and smooth muscle fibers: relation and function. J Urol, 154, 1736–1739.
Schultz, N. G., Lough-Stevens, M., Abreu, E., Orr, T., & Dean, M. D. (2016). The Baculum was gained and lost multiple times during mammalian evolution. Integr Comp Biol, 56, 644–656.
Short, R. V. (1981). Sexual selection in man and the great apes. In C. E. Graham (Ed.), Reproductive biology of the great apes: comparative and biomedical perspectives. New York: Academic Press.
Simmons, L. W., & Firman, R. C. (2014). Experimental evidence for the evolution of the mammalian baculum by sexual selection. Evolution, 68, 276–283.
Stockley, P., Ramm, S. A., Sherborne, A. L., Thom, M. D., Paterson, S., & Hurst, J. L. (2013). Baculum morphology predicts reproductive success of male house mice under sexual selection. BMC Biol, 11, 66.
Suresh, S., & Prakash, S. (2011). Effect of Mucuna pruriens (Linn.) on oxidative stress-induced structural alteration of corpus cavernosum in streptozotocin-induced diabetic rat. J Sex Med, 8, 1943–1956.
Suresh, S., & Prakash, S. (2012). Effect of Mucuna pruriens (Linn.) on sexual behavior and sperm parameters in streptozotocin-induced diabetic male rat. J Sex Med, 9, 3066–3078.
Vilmann, A., & Vilmann, H. (1983). Os penis of the rat. IV. - proximal growth cartilage. Acta Anat, 117, 136–144.
Vilmann, H. (1982). Os penis of the rat. III. - formation and growth of the bone. Acta Morphol Neerl Scand, 20, 309–318.
Weihe, W. H. (1987). The laboratory rat. The UFAW handbook on the care and management of laboratory animals, (6th ed., ). Harlow: Longman.
Williams-Ashman, H. G. (1990). Enigmatic features of penile development and functions. Perspect Biol Med, 33, 335–374.
Wu, C., & Kovac, J. R. (2016). Models for erectile dysfunction and their importance to novel drug discovery. Expert Opin Drug Discovery, 11, 185–196.
Yamamoto, M. (1987). Histology of the os penis of the rat. I. - light microscopic study of the distal segment and possible origin of its osteocytes. Tokyo Med J, 10, 303–313.
Yilmaz, I. E., Barazani, Y., & Tareen, B. (2013). Penile ossification: a traumatic event or evolutionary throwback? Case report and review of the literature. Cana Urol Assoc, 7, 112–114.
Yonezawa, T., Higashi, M., Yoshioka, K., & Mutoh, K. (2011). Distribution of aromatase and sex steroid receptors in the baculum during the rat life cycle: effects of estrogen during the early development of the baculum. Biol Reprod, 85, 105–112.
Yoshida, H., & Huggins, C. B. (1980). Induction of ossification in the clitoris of neonatal female rats by administration of androgens. Endocrinology, 106, 1956–1959.
Zaid, U. B., Alwaal, A., Zhang, X., & Lue, T. F. (2014). Surgical management of Peyronie’s disease. Curr Urol Rep, 15, 446.
Funding
No special fund is received for the research. The work was carried out as a faculty individual research activity of the University of Madras through the basic annual budget head.
Availability of data and materials
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
Author information
Authors and Affiliations
Contributions
SP contributed to the conceptualization, study design, data processing, and writing the manuscript. MI contributed to the methodology, data collection, and input for drafting the manuscript. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Protocol for the research project has been approved by the Institute Animal Ethics Committee (IAEC) of Dr. ALM PGBIMS, University of Madras.
This article does not contain any studies with human subjects performed by any of the authors.
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Prakash, S., Ibrahim, M. The penile baculum in monkey (Macaca radiata), rat (Rattus norvegicus), and bat (Pipistrellus mimus) and its functional significance—a mini review. JoBAZ 79, 38 (2018). https://doi.org/10.1186/s41936-018-0052-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41936-018-0052-4