
Abstract
Background
The flavonoid prunin is a flavanone glycoside found in cherry trees, including the flowering cherry Prunus yedoensis Matsumura (Rosacea). Although this compound has been studied for its antioxidant, anti-bacterial, and blood-sugar-lowering effects, no studies address its use in cosmetics. This study investigates whether prunin exhibits anti-inflammatory effects in cells exposed to ultraviolet B (UVB) radiation.
Methods
The effects of prunin were assessed by measuring cell viability using the water-soluble tetrazolium salt-1 assay and free radical damage using the dichlorofluorescein diacetate assay as well as by quantitative real-time PCR.
Results
UVB-induced decrease in cell viability diminished by pretreatment with prunin in a concentration-dependent manner. Intracellular reactive oxygen species (ROS) quantitative analysis revealed that the expression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), which is associated with the inflammatory response, and mRNA expression of interleukin-6 (IL-6), interleukin-8 (IL-8), cyclooxygenase-2 (COX2), tumor necrosis factor-alpha (TNF-α), and protease-activated receptor (PAR2) decreased with prunin pretreatment in a concentration-dependent manner.
Conclusions
Prunin increased the survival rate of UVB-treated human keratinocyte HaCaT cells. Prunin protected the HaCaT cells, eliminated ROS, and demonstrated anti-inflammatory effects. Thus, prunin is worthy of investigation for use as a cosmetic ingredient that protects the skin and has anti-inflammatory effects.
Similar content being viewed by others

Background
The skin is put under oxidative stress when UV radiation induces the formation of reactive oxygen species (ROS), which damage DNA, lipids, and proteins (Devasagayam and Kamat 1994). Ultraviolet light acts as an external stress-stimulating factor, which increases the expression and activity of 11β-hydroxysteroid ehydrogenase type 1 (11β-HSD1), an enzyme that converts ROS and inactive cortisone to active cortisol in keratinocytes (Itoi et al. 2013).
Increased oxidative stress in the skin leads to inflammatory responses such as erythema, edema, and fever. These responses attempt to restore and regenerate wounds caused by chemical, physical, and bacterial assaults (Greaves and Sondergaard 1970; Hruza and Pentland 1993). During an inflammatory response, immune cells such as monocytes and macrophages are stimulated to express inflammation-related genes through the activation of transcription factors. Macrophages express the transcription factor nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) in response to stimuli such as lipopolysaccharide and interferon-γ. NF-κB induces inflammatory responses by producing nitric oxide synthase, nitric oxide, prostaglandins, and cyclooxygenase-2 (COX2) (Nathan 1987). NF-κB is usually bound to inhibitor kappa B (IκB), which maintains the inactive state. When exposed to external stimuli such as UV light, IκB is rapidly phosphorylated and degraded (Maziere et al. 1999; Nomoto et al. 2001; Gomez-Nicola et al. 2010; Baeuerle and Henkel 1994). After NF-κB separates from IκB and becomes activated, it migrates to the nucleus, where it increases the expression of genes that induce inflammatory responses, including interleukin-6 (IL-6), interleukin-8 (IL-8), tumor necrosis factor-alpha (TNF-α), and COX2 (Baeuerle and Baltimore 1988). In addition, a variety of proteases are expressed in the skin. These proteases are involved in maintaining homeostasis through immune response, inflammation, cytokine expression, vascular function, tissue recovery, host defense, and apoptosis (Sharlow et al. 2000).
Protease-activated receptor 2 (PAR2) plays an important role in inflammation. In 1995, we observed PAR-2 expression in Santulli in vitro. D ‘Andrea observed PAR-2 expression and formed surface, superficial, and granular layers. PAR-2 and PAR-4 indicate that trypsin activates mast cell tryptase PAR-2 (Santulli et al. 1995; Coughlin 1999; D’Andrea et al. 1998). In addition, trypsin, a mast cell tryptase or SLGRL-NH2 (synthetic peptide) also activates PAR-2 and causes extensive inflammation (Kong et al. 1997).
Prunin is a flavonoid, which is a family of polyphenolic compounds with its name derived from the Greek word flavus that means yellow. This plant pigment possesses flavone as its basic structure. Found in relatively small amounts in animals, prunin is mainly present in vegetables, fruits, flowers, leaves, stems, roots, and fruits (Middleton et al. 2000). Prunin is a flavanone glycoside with the molecular formula C21H22O10 and a molecular weight of 434.39. Prunin was isolated from P. davidiana Fr. (Rosacea) stems (Jung et al. 2017). The stems of P. davidiana Fr. (Rosacea) are used in Korean traditional medicine for treating neuritis and rheumatism (Choi et al. 1991). These stems contain antiviral agents and a variety of flavonoids that possess antioxidant, anti-inflammatory, hypocholesterolemic, and cardioprotective properties, and the DNA-binding activity (Jung et al. 2003; Yousuf et al. 2013; Zhang et al. 2008). Previous studies on prunin report that it has antioxidant and anti-inflammation properties, decreases blood glucose and insulin levels, and improves blood circulation (Céliz et al. 2013; Céliz et al. 2010; Céliz et al. 2011; Choi et al. 2006; Kimihisa et al. 2010). In this study, we investigated the effects of prunin treatment on ultraviolet (UVB)-induced cell damage and inflammation.
Methods
Cell culture
The human keratinocyte HaCaT cells used in this experiment were purchased from ATCC (USA). To incubate HaCaT cell strains, Dulbecco’s Modified Eagle Medium (Hyclone, USA) containing 10% fetal bovine serum (Hyclone) and 1% penicillin/streptomycin (penicillin 100 IU/ml, streptomycin 100 μg/ml; Invitrogen, USA) was used. The cells were incubated at 37 °C with 5% CO2.
Sample treatment
Prunin was purchased from Sigma-Aldrich (USA) in purified (> 95%) powder, which was dissolved in dimethyl sulfoxide (Sigma-Aldrich) at an appropriate concentration for the experiment. HaCaT (1 × 106cell/well) was incubated in the culture dish for 24 h, and prunin was subsequently added at an appropriate concentration and preprocessed for 6 additional hours. A UVB lamp (UVP, USA) was used to irradiate the cells with UVB. The wavelength of UVB irradiation was measured with a fiberoptic Spectrometer System USB2000 (Ocean Optics, USA). After the irradiation, PBS was removed, and a new medium was added, followed by additional incubation of 24 h prior to use in the experiment.
Cell viability estimation
The principles of the WST-1 assay were used for the experiment on cell survival. Overall, 100 μL HaCaT (3 × 103cells/well) was inoculated on a 96-well plate and incubated for 24 h. Afterward, prunin was added at 1, 5, 10, 20, and 40 μM and 40 mJ/cm2 and UVB was irradiated to these cells, which were subsequently incubated for additional 24 h. In addition, 10 μL of EZ-Cytox cell viability assay kit reagent (ITSBio, Korea) was added to the incubated cells for 1 h, and the absorbance was measured using a microplate reader (Bio-Rad, USA) at 490 nm.
Quantitative analysis of intracellular ROS
In total, 10 μM dichlorofluorescein diacetate was added as a dye to measure intracellular ROS. The cells were harvested after incubation for 30 min. After adding PBS to release the cells, a flow cytometer (BD Biosciences, USA) was used to measure the changes in ROS. To verify the removal effects of prunin on ROS, N-acetyl-L-cysteine (NAC) was used as a positive control group that acts as a ROS scavenger and measured after the same treatment as prunin.
Quantitative real-time PCR
The cells obtained from the cell culture were extracted with Trizol reagent (Invitrogen, USA). The extracted RNA was quantified using a Nanodrop (Maestrogen, USA), and only RNA with a purity of 260 nm/280 nm (ratio 1.8) was used for the experiment.
In total, 10 μl of DNA was prepared by adding 1 μg RNA, 0.5 ng oligo dT18, and DEPC water in a PCR tube and processing it for 10 min at 70 °C to induce RNA denaturation. Subsequently, M-MLV reverse transcriptase (Enzynomics, Korea) was used to induce a reaction at 37 °C for 1 h to synthesize cDNA.
For qRT-PCR, 0.2 μM primers, 50 mM KCl, 20 mM Tris/HCl pH 8.4, 0.8 mM dNTP, 0.5 U Extaq DNA polymerase, 3 MgCl2, and 1X SYBR green (Invitrogen) were mixed in a PCR tube to produce a reagent. PCR was performed with Linegene K (BioER, China) through 40 cycles (3 min per cycle) of denaturation, denaturation (94 °C, 30 s), annealing (58 °C, 30 s), and polymerization (72 °C, 30 s) at 94 °C. The significance of PCR was validated using a melting curve. The gene expressions were compared for analysis by normalizing the β-actin expression. The primers used for this experiment are shown in Table 1.
Statistical analysis
Each experiment in this study was performed three times or more under the same conditions, and the results are expressed as mean and standard deviation. For each experiment, the Student’s t test was used to analyze all findings, with a p value of 0.05 or below considered as statistically significant.
Results
Cytotoxic and protective effects of prunin
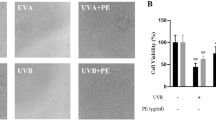
The water-soluble tetrazolium salt-1 assay was performed to determine the cytotoxicity of prunin in human keratinocyte HaCaT cells. Treatment of the cells with prunin at concentrations of 1, 5, 10, 20, and 40 μM revealed the following survival rates: 1 μM, 100%; 5 μM, 102%; 10 μM, 102%; 20 μM 93%, and 40 μM, 75%. These results suggest that prunin is cytotoxic at concentrations ≥ 40 μM (Fig. 1a). To examine the cytoprotective effects of prunin, damaged HaCaT cells were treated with prunin (5 or 10 μM) and irradiated with 40 mJ/cm2 UVB. Compared to the cell survival rate of 53% in the control group, the survival rate of prunin-treated cells increased to 71% (5 μM) and 96% (10 μM). The cell survival rate was increased by prunin treatment in a concentration-dependent manner (Fig. 1b).

Cell viability on prunin in HaCaT keratinocytes. a The cellular toxicity of prunin on HaCaT keratinocytes (*p < .05). b The effect of prunin on cell viability in UVB-irradiated HaCaT keratinocytes (**p < .01, ***p < .001)
ROS elimination effect of prunin in HaCaT cells irradiated with UVB
The cell-permeable reagent 2′,7′-dichlorofluorescin diacetate was used to investigate the effect of prunin on ROS in UVB-irradiated HaCaT cells. This reagent fluorescent stains intracellular ROS, which were then analyzed by flow cytometry. ROS increased from 1 to 7.28 in cells exposed to 40 mJ/cm2 UVB. Treatment of cells with prunin at concentrations of 5 and 10 μM before irradiation resulted in a decrease in ROS to 4.32 and 1.57, respectively. As a positive control, treatment with 10 mM NAC resulted in a decrease to 1.35, a result similar to that after treatment with 10 μM prunin (Fig. 2).

The ROS scavenging effect of prunin in UVB-irradiated HaCaT keratinocytes (***p < .001)
Anti-inflammatory effect of prunin on UVB-irradiated HaCaT cells
The expression level of IL-6, IL-8, COX2, TNF-α, and PAR2 was confirmed by quantitative real-time PCR to assess the effect of prunin on UVB-induced inflammation in HaCaT cells. First, we examined the effect of prunin on the UVB-induced increase in IL-6 mRNA (5.17). Cells treated with prunin at concentrations of 5 and 10 μM before UVB exposure exhibited a decrease in IL-6 mRNA to 3.55 and 1.40, respectively. Prunin treatment decreased IL-6 mRNA expression in a concentration-dependent manner (Fig. 3a). The IL-8 mRNA expression of 3.17 after UVB exposure decreased to 2.43 and 1.78 with prunin treatment at 5 and 10 μM, respectively. Prunin decreased IL-8 mRNA expression in a concentration-dependent manner (Fig. 3b). COX2 mRNA, encoding a protein that induces inflammation, increased to 8.14 after UVB exposure. Pretreatment with 5 or 10 μM prunin decreased COX2 mRNA expression to 4.55 and 1.23, respectively. Prunin decreased COX2 mRNA expression in a concentration-dependent manner (Fig. 3c). TNF-α mRNA expression increased to 5.04 after UVB exposure. Pretreatment with 5 or 10 μM prunin decreased TNF-α mRNA expression to 3.26 and 1.26, respectively. Prunin decreased TNF-α mRNA expression in a concentration-dependent manner (Fig. 3d). PAR2 mRNA increased to 3.80 upon UVB exposure. Pretreatment with 5 or 10 μM prunin resulted in a decrease in PAR2 mRNA expression to 1.83 and 1.06, respectively. Prunin decreased PAR2 mRNA expression in a concentration-dependent manner (Fig. 3e).

The effect of prunin on inflammation in UVB-irradiated HaCaT. a The effect of prunin on expression of IL-6 mRNA in UVB-irradiated HaCaT keratinocytes (**p < .01, ***p < .001). b The effect of prunin on expression of IL-8 mRNA in UVB-irradiated HaCaT keratinocytes (**p < .01, ***p < .001). c The effect of prunin on expression of COX-2 mRNA in UVB-irradiated HaCaT keratinocytes (***p < .001). d The effect of prunin on expression of TNF-α mRNA in UVB-irradiated HaCaT keratinocytes (**p < .01, ***p < .001). e The effect of prunin on expression of PAR2 mRNA in UVB-irradiated HaCaT keratinocytes (***p < .001)
Discussion
Intracellular damage by oxidative stress causes cell cycle arrest and cellular aging and increases the production of ROS (Cerella et al. 2009). Intracellular ROS can damage DNA and mitochondria, resulting in abnormalities in energy metabolism along with protein oxidation (Lang et al. 2018). In addition, ROS activates NF-κB, which promotes the expression of genes that induce inflammatory responses (Nathan 1987; Baeuerle and Baltimore 1988).
We observed here that exposing HaCaT cells to 40 mJ/cm2 UVB leads to an increase in ROS. Figure 2 shows that prunin inhibits this UVB-induced increase in ROS in HaCaT cells in a concentration-dependent manner, inhibits the increased cellular aging caused by UVB, and is effective for ROS elimination. In addition, we observed that exposure of these cells to 40 mJ/cm2 UVB causes a rapid increase in the activity of NF-κB, a proinflammatory transcription factor. Prunin pretreatment inhibited this increase in activity in a concentration-dependent manner. In other words, prunin inhibits the activation of NF-κB in the nucleus, thereby decreasing the expression of several inflammation-related substances. In particular, the expression levels of IL-8, IL-6, COX2, TNF-α, and PAR2, which are induced upon activation of NF-κB, decreased with increasing concentrations of prunin. These results suggest that prunin inhibits UVB-induced inflammation in HaCaT cells and therefore is effective for anti-inflammatory skin.
Conclusions
Prunin pretreatment protects HaCaT cells against UVB-induced damage by preventing the generation of ROS and decreasing the expression of IL-6, IL-8, COX2, TNF-α, and PAR2 in a concentration-dependent manner. Prunin improves the survival rate of UVB-damaged HaCaT cells and blocks the NF-κB pathway through ROS elimination, thereby inhibiting inflammatory responses. In summary, prunin has cytoprotective and anti-inflammatory effects in HaCaT cells.
Abbreviations
- 11β-HSD1:
-
11β-hydroxysteroid ehydrogenase type 1
- COX2:
-
Cyclooxygenase-2
- IL-6:
-
Interleukin-6
- IL-8:
-
Interleukin-8
- IκB:
-
Inhibitor kappa B
- NAC:
-
N-acetyl-L-cysteine
- NF-κB:
-
Nuclear factor kappa-light-chain-enhancer of activated B cells
- PAR2:
-
Protease-activated receptor 2
- ROS:
-
Reactive oxygen species
- TNF-α:
-
Tumor necrosis factor-alpha
- UVB:
-
Ultraviolet
References
Baeuerle PA, Baltimore D. I kappa B: a specific inhibitor of the NF-kappa B transcription factor. Science. 1988;242(4878):540–6.
Baeuerle PA, Henkel T. Function and activation of NF-kappa B in the immune system. Annu Rev Immunol. 1994;12(1):41–79.
Céliz G, Audisio MC, Daz M. Antimicrobial properties of prunin, a citric flavanone glucoside, and its prunin 6″-O-lauroyl ester. J Appl Microbiol. 2010;109(4):1450–7.
Céliz G, Daz M, Audisio MC. Antibacterial activity of naringin derivatives against pathogenic strains. J Appl Microbiol. 2011;111(3):731–8.
Céliz G, Alfaro FF, Cecilia C, Daz M, Verstraeten SV. Prunin- and hesperetin glucoside-alkyl (C4-C18) esters interaction with Jurkat cells plasma membrane: consequences on membrane physical properties and antioxidant capacity. Food Chem Toxicol. 2013;55:411–23.
Cerella C, Coppola S, Maresca V, De Nicola M, Radogna F, Ghibelli L. Multiple mechanisms for hydrogen peroxide-induced apoptosis. Ann N Y Acad Sci. 2009;1171:559–63.
Choi JS, Yokozawa T, Oura H. Antihyperlipidemic effect of flavonoids from Prunus davidiana. J Nat Prod. 1991;54(1):218–24.
Choi JS, Yokozawa T, Oura H. Improvement of hyperglycemia and hyperlipemia in streptozotocin-diabetic rats by a methanolic extract of Prunus davidiana stems and its main component, prunin. Planta Med. 2006;57(3):208–19.
Coughlin SR. How the protease thromgin talks to cells. Proc Natl Acad Sci U S A. 1999;96(20):11023–7.
D’Andrea MR, Derian CK, Leturcq D, Baker SM, Grunmark A, Ling P. Characterization of protease-activatedreceptor-2 immunoreactivity in normal human tissues. J Histochem Cytochem. 1998;46:157–64.
Devasagayam TP, Kamat JP. Biological significance of oxygen. Indian J Exp Biol. 1994;67:319–25.
Gomez-Nicola D, Valle-Argos B, Nieto-Sampedro M. Blockade of IL-15 activity inhibits microglial activation through the NF-kappaB, p38, and ERK1/2 pathways, reducing cytokine and chemokine release. Glia. 2010;58(3):264–76.
Greaves MW, Sondergaard J. Pharmacological agents released in ultraviolet inflammation studies by continuous skin perfusion. J Invest Dermatol. 1970;54:365–7.
Hruza L, Pentland AP. Mechanisms of UV-induced inflammation. J Invest Dermatol. 1993;100:35–41.
Itoi S, Terao M, Murota H, Katayama. 11β-Hydroxysteroid dehydrogenase 1 contributes to the pro-inflammatory response of keratinocytes. Biochem Biophys Res Commun. 2013;440(2):265.
Jung HA, Jung MJ, Kim JY, Chung HY, Choi JS. Inhibitory activity of flavonoids from Prunus davidiana and other flavonoids on total ROS and hydroxyl radical generation. Arch Pharm Res. 2003;26(10):809–15.
Jung HA, Ali MY, Bhakta HK, Min B-S, Choi JS. Prunin is a highly potent flavonoid from Prunus davidiana stems that inhibits protein tyrosine phosphatase 1B and stimulates glucose uptake in insulin-resistant HepG2 cells. Arch Pharm Res. 2017;40:37–48.
Kimihisa I, Megumi M, Shunsuke N, Kazuya M, Hideaki M. Effects of unripe Citrus hassaku fruits extract and its flavanone glycosides on blood fluidity. Biol Pharm Bull. 2010;33(4):659–64.
Kong W, Mcconalogue K, Khitin LM, Hollenberg MD, Payan DG, Bohm SK, Bunnett NW. Luminal trypsin may regulate enterocytes through proteinase-activated receptor-2. Proc Natl Acad Sci U S A. 1997;94:8884–9.
Lang JY, Ma K, Guo JX, Sun H. Oxidative stress induces B lymphocyte DNA damage and apoptosis by upregulating p66shc. Eur Rev Med Pharmacol Sci. 2018;22:1051–60.
Maziere C, Conte MA, Degonville J, Ali D, Maziere JC. Cellular enrichment with polyunsaturated fatty acids induces an oxidative stressand activates the transcription factors AP-1 and NF-kappaB. Biochem Biophys Res Commun. 1999;265(1):116–22.
Middleton E, Kandaswami C, Theoharides TC. The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease, and cancer. Pharmacol Reu. 2000;52(4):673–751.
Nathan CF. Secretory products of macrophages. J Clin Invest. 1987;79:319–26.
Nomoto Y, Yamamoto M, Fukushima T, Kimura H, Ohshima K, Tomonaga M. Expression of nuclear factor kappaB and tumor necrosis factoralpha in the mouse brain after experimental thermal ablation injury. Neurosurgery. 2001;48:158–66.
Santulli RJ, Derian CK, Darrow AL, Tomko KA, Eckardt AJ, Seiberg M. Evidence for the presence of a protease-activated receptor distinct from the thrombin receptor in human keratinocytes. Proc Natl Acad Sci U S A. 1995;92:9151–5.
Sharlow ER, Paine CS, Babiarz L, Eisinger M, Shapiro S, Seiberg M. The protease-activated receptor-2 upregulates keratinocyte phagocytosis. J Cell Sci. 2000;113:3093–101.
Yousuf S, Sudha N, Murugesan G, Enoch IVMV. Isolation of prunin from the fruit shell of Bixa orellana and the effect of bcyclodextrin on its binding with calf thymus DNA. Carbohydr Res. 2013;365:46–51.
Zhang L, Liu W, Hu T, Du L, Luo C, Chen K, Shen X, Jiang H. Structural basis for catalytic and inhibitory mechanisms of beta-hydroxyacyl-acyl carrier protein dehydratase (FabZ). J Biol Chem. 2008;283(9):5370–9.
Acknowledgements
Not applicable
Funding
Not applicable
Availability of data and materials
Not applicable
Author information
Authors and Affiliations
Contributions
EJN and JYR did all of research background such as experiments, data collecting, and statistical analysis as well as draft manuscript. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Na, E.J., Ryu, J.Y. Anti-inflammatory effects of prunin on UVB-irradiated human keratinocytes. biomed dermatol 2, 14 (2018). https://doi.org/10.1186/s41702-018-0024-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41702-018-0024-9