
Abstract
Background
The loss of traditional agropastoral systems, with the consequent reduction of foraging habitats and prey availability, is one of the main causes for the fast decline of Lesser Kestrel (Falco naumanni). To promote the conservation of the Lesser Kestrel and their habitats, here we studied the foraging activities patterns of this species during the breeding season.
Methods
Between 2016 and 2017, we captured and tagged 24 individuals with GPS dataloggers of two colonies in Villena (eastern Spain) with the goals of estimating the home range sizes of males and females, evaluating the differences in spatial ecology between two colonies located in different environments: natural and beside a thermosolar power plant, and investigating habitat selection.
Results
Considering the distances before July 15, date until which it can be assured that the chicks remain in the nest in our colonies, there were significant differences with the distances to the nest in relation to the colony of the individuals: Lesser Kestrels from the thermosolar power plant colony had a greater average distance. The average size of home range areas was 13.37 km2 according to 95% kernel, and there were also significant differences in relation to colony: the individuals from the thermosolar power plant colony used a larger area (22.03 ± 4.07 km2) than those from the other colony (9.66 ± 7.68 km2). Birds showed preference for non-irrigated arable lands and pastures.
Conclusions
Despite the differences between the two colonies, the home ranges of both are smaller or similar to those observed in other European colonies. This suggests that Lesser Kestrels continue to have adequate habitats and a good availability of prey. Therefore, the extension and proximity of the plant does not imply a great alteration, which highlights the importance of maintaining the rest of the territory in good conditions to minimize the impact.
Similar content being viewed by others


Background
Lesser Kestrel (Falco naumanni, Fleischer 1818) is one of the smallest European raptors (wing-span 58–72 cm, body mass 120–140 g). It is a trans-Saharan migratory falcon that overwinters in Sahel area (Limiñana et al. 2012) and breeds colonially in cavity sites in old buildings or under roof tiles in western Europe (Negro and Hiraldo 1993). Lesser Kestrel is predominantly insectivorous, feeding mostly on large invertebrates (mainly Orthoptera, Coleoptera and Scolopendridae) (Franco and Andrada 1977; Negro 1997; Rocha 1998; Lepley et al. 2000; Kotsonas et al. 2018) and secondarily on spiders and small mammals and lizards (Cramp and Simmons 1980; Tella et al. 1996; Negro 1997), whose density in steppe habitats seems to be high due to their wide floristic diversity and composition (Wiens 1985, 1989; Tellería et al. 1988; Martínez 1994; Moreira 1999; Clere and Bretagnolle 2001). Lesser Kestrel populations declined strongly during the second half 20th century, especially in its European range (it became extinct in several countries and has practically disappeared in others (Tucker and Heath 1994; Rodríguez et al. 2006)), due to a reduction in the availability of nest sites, increases in the use of agricultural pesticides, changes in land use (Bustamante 1997) and above all, loss of traditional agropastoral systems, that caused a reduction of foraging habitats and prey availability (Tella et al. 1998).
In this species, low breeding success strongly influences population dynamics (Hiraldo et al. 1996) and has been attributed to low hunting performance in intensively used agricultural landscapes (Donázar et al. 1993; Tella et al. 1998). The current situation in many breeding areas of low prey availability for this species may stress the effect of changes in land-use management on Lesser Kestrel behaviour, especially hunting behaviour (García et al. 2006). However, the population has been considered stable for the last two decades, and consequently, it was downlisted from ‘Vulnerable’ to ‘Least Concern’ in 2011 according to IUCN criteria (BirdLife International 2018).
Lesser Kestrel is currently associated with high value habitats of the western Palaearctic, such as Palearctic pseudo-steppes, where the species typically inhabits during the breeding season (Cramp and Simmons 1980). Extensive cereal fields, fallows, pasturelands and field margins in agricultural areas are the main habitats used by Lesser Kestrels for hunting (Donázar et al. 1993; Tella et al. 1998; Gustin et al. 2014; Vlachos et al. 2015). Therefore, monitoring Lesser Kestrels will promote a better understanding of management strategies that could prove useful for preserving European steppe lands (Tella et al. 1998) and, furthermore, will provide insight into the ways in which European agricultural policies could be adapted to significantly reduce the rate of biodiversity loss (European Commission 2006). Although there are many previous studies about space use by Lesser Kestrel (García et al. 2006; Catry et al. 2014; Vlachos et al. 2015; Di Maggio et al. 2016), recently the lack of concordance in their results has been pointed; also, these studies comprise a set of local truths that make it difficult to establish overall species management recommendations (Rodríguez et al. 2014). The size of the home range areas has also been addressed previously, obtaining great differences according to the suitability of the habitat (Tella et al. 1998; Catry et al. 2013; Vlachos et al. 2015).
For all these reasons, to promote the conservation of the Lesser Kestrel and their habitats, our goal was to study the spatial ecology of the species during the breeding season in an area that meets special conditions. On the one hand, it is a reintroduced population located in the distribution limit of the species (to the east) and, on the other hand, the habitat is strongly transformed during several years (changes in land use, mainly, substitution of cereal lands for vine crops and construction of new infrastructures that have reduced the suitable habitat for the species). Therefore, the main objectives of the present study were (1) to estimate the home range sizes of males and females during the breeding season; (2) evaluate the differences in spatial ecology between two colonies located in different environments: natural and beside a thermosolar power plant and (3) to estimate foraging habitat selection.
Methods
Study area
Fieldwork was carried out in the agricultural pseudo-steppes of ‘Los Alhorines’ (Villena, southeastern Spain). Here there is a 6500 ha Special Protection Area (SPA) created for the benefit of its important steppe bird populations (mainly Lesser Kestrel, Little Bustard Tetrax tetrax, Great Bustard Otis tarda, Montagu’s Harriers Circus pygargus, Black-bellied Sandgrouse Pterocles orientalis and Pin-tailed Sandgrouse P. alchata).
The climate is typically Continental-Mediterranean with relatively cold wet winters and dry hot summers; the average annual temperature is 15.3 °C and the rainfall 421 mm (Climate Data 2018). The study area is dominated by a mosaic of dry crops, including extensive dry cereal (wheat, and especially barley), fallows of variable ages, stubbles plots (grazed by livestock, usually sheeps), olive groves, vineyards, sunflower and a few patches of dry annual legume crops and irrigated crops (mainly maize). This valley has been historically transformed into a large pseudo-steppe devoted to dry cereal farming using traditional practices, such as the maintenance of fallows, little use of fertilizers and biocides, and the existence of numerous field margins. However, current vineyards are being developed to replace the cereal fallow system with intensive cultures. Nowadays, cereal, ploughed fallow and vineyards constituted the main crops. In addition, two human constructions stand out in the landscape: a thermosolar power plant of 1.1 km2 completed in 2013 and a reservoir with capacity for more than 20 hm3 (now dry due to filtrations).
Lesser Kestrel population was established in the study area after a reintroduction project carried out between 1997 and 2002 (Alberdi 2006). In 2000, three pairs bred for the first time in our study area, and currently the total population reaches 70–80 pairs. Most pairs breed naturally in the roofs of abandoned or semi-abandoned farmhouses, and some in nest-boxes arranged in different points. The number of breeding pairs per house ranges along the years; the largest colony usually hosts 12–16 pairs each year, while there are a lot of smaller ones (between 5–8 colonies per year), some of them with only 1–2 pairs. The number of farmhouses occupied by breeding Lesser Kestrels varies between years, but in the study area, there are a total of 27 different Lesser Kestrel colonies.
Data collection
Lesser Kestrels were captured in two different colonies between 2016 and 2017. One of them is located in an abandoned country house beside the thermosolar power plant (the distance between this house and the solar panels is 60 m), with cereal crops around but also near a highway, a prison and a high-speed train line; in this colony, around 14 Lesser Kestrels pairs breed each year in natural nests under the tiles. The other one, with 6–8 couples each breading season, is located in three towers with nest-boxes arranged for the breeding of this species; the surroundings of this colony are composed by different farmlands patches (with a high percentage of fallows, cereal crops, natural meadows, uncultivated lands and some vineyards and almond groves). The distance between the two colonies is about 6.5 km. In the breeding seasons of 2016 and 2017 a total of 24 adult Lesser Kestrels were captured, 8 from the thermosolar power plant colony and 16 from the towers with nest-boxes (14 males and 10 females, see Table 1 for more details). Birds were trapped on nest-boxes or using a dho-gaza net with a stuffed Eagle Owl (Bubo bubo) used as a decoy, placed close to their nests (25–50 m). To prevent nest abandonment, birds were only trapped in the late stage of incubation or when they had small nestlings (from 20 May to 20 June, approximately). Birds were tagged using 5-g datalogger (nanoFix-GEO + RF tags from PathTrack Ltd., Leeds, United Kingdom). Tags were programmed to collect GPS locations every 15 min from 06:00 h to 20:00 h (local hour) during May, June and July, and every 30 min with no off periods during August and September. Datalogger tags were affixed to the back of birds using a Teflon harness, a non-abrasive material (e.g. Kenward 2001), which was sew with a cotton thread in a single point to enable its liberation when the thread is wear out (both the tag and the harness). Weight of datalogger was below the recommended 5% of bird’s body mass (Kenward 2001; mean percentage ± SD = 4.04 ± 0.42%, n = 20, range = 3.38–4.95%). Birds were released in a maximum of 40 min after capture and they were visually tracked to make sure that they resumed their normal activities. We did not observe any adverse effect of the devices on behaviour or reproduction for tagged birds that remained in the study area.
With 24 tagged Lesser Kestrels, good quality data were obtained from 20 individuals (Table 1), with an average of 48 ± 27 tracking days during the breeding season and 1432 ± 801 locations/individual. For the other 4 individuals, data have been obtained for a very few days (#16413 and #16416 only 1 day being tracked, and #16667 and #16675 2 and 5 days respectively) due to device failure, death of the individual or abandonment of the study area. Data of these individuals were not used in the analyses to avoid an underestimation of the breeding area. The data of the tags were collected through mobile UHF data reception stations. For the analysis we only have considered the dates in which the individuals remain in the breeding area, discarding the locations corresponding to the premigratory movements on account of their leaving from the breeding area (García 2000).
Distance to the nest and home range
We calculated the distance to the nest position of every recorded location of all Lesser Kestrels. Besides considering the whole study period, we have used July 15 (date from which it can be assured that the chicks have become independent in our colonies and therefore the parents do not need to feed them) as separation in dates to evaluate possible differences in the travelled distances. The home range of each individual was estimated using fixed kernel method (Worton 1989), including all GPS locations available for the study period. We calculated the 95%, 75% and 50% fixed kernels using the Animal Movement extension for ArcView 3.2 (Hooge and Eichenlaub 1997) and the least squares cross-validation procedure to calculate the smoothing parameter H (Silverman 1986). Fixed kernel estimators are widely used in the literature and allow comparison with similar studies (Worton 1989; Mellone et al. 2011; Limiñana et al. 2012; Gustin et al. 2017). We also calculated the 100% minimum convex polygon (MCP) for each breeding area of each Lesser Kestrel. The real size of home ranges was estimated transforming the polygons to an Equal-Area Cylindrical projection.
Habitat selection
To describe the habitat use within the home ranges, we used the CORINE 2012 land cover map provided by the European Environment Agency (https://www.eea.europa.eu/data-and-maps/data/clc-2012-raster). To estimate the habitat availability within the areas of the two colonies and to evaluate whether Lesser Kestrels are found in certain habitats more frequently than expected by their availability we performed a habitat selection analysis. We calculated the percentage of every habitat type within the combined MCP, which represents the maximum potential area. Data of each colony were treated separately, so we evaluated the differences in habitat availability between the two colonies using the Chi squared test in contingency tables. We generated 40,000 random points within these combined MCPs and we assigned the corresponding habitat type to every random point and to every location recorded during the breeding season. To determinate habitat preferences we used Monte Carlo simulations (Manly 1997; Soutullo et al. 2008; Limiñana et al. 2011; López-López et al. 2016), comparing the frequency of real observed locations with the expected frequencies according to random locations. These expected frequencies were calculated by sampling the same number of real locations from the generated random points; this process was repeated 1000 times using the “shuffle rows” option in Excel’s PopTools add-in (Hood 2010). With Monte Carlo analysis the observed values (selected as dependent range) were compared against the 1000 replicated of expected values (selected as test values). These analyses were conducted using Excel’s PopTools and were performed separately for each colony. Comparisons were two-tailed and significance level was established at p < 0.05.
We grouped the land cover classes (“CLC”) with assigned values into eleven categories to facilitate the interpretation of the results: artificial surfaces (CLC codes: 112, 121, 131 and 133), non-irrigated arable land (CLC code: 211), permanently irrigated land (CLC code 212), vineyards (CLC code: 221), arboreal crops (CLC codes: 222 and 223), pastures (CLC code: 231), heterogeneous agricultural areas (CLC codes: 242 and 243), forests (CLC codes: 312 and 313), scrub and/or herbaceous vegetation associations (CLC codes: 321, 323 and 324), sparsely vegetated areas (CLC code: 333) and water bodies (CLC code: 512). We also calculated the number of locations that were within the ‘Los Alhorines’ SPA.
Statistical analysis
To evaluate the effect of the colony of origin and the sex in the spatial parameters (distance to the nest and home ranges) we used Linear Mixed Models (LMMs). The distances to the nest and the different measurements of the home range areas (kernel estimations and MCP) were included as dependent variables and a model was developed for each one. In the case of the distances we used the whole dataset of locations and for the home range areas we used the corresponding for each individual. The variables “colony” and “sex” were considered as fixed factors, and the “individual” and “year” as random factors.
All statistical analyses (including t-Student tests to evaluate differences between some parameters, descriptive statistics and Chi squared tests) were performed with IBM SPSS Statistics ver. 22.0. Significance level was established at p < 0.05 and the results are presented as mean ± standard deviation (SD).
Results
Data have been obtained for 20 individual breeding areas, with a total of 28,645 locations (10,577 in the thermosolar power plant colony and 18,068 in the towers). The tracking days for the individuals of the thermosolar power plant colony were on average 59 ± 30 with 2003 ± 1099 locations/individual, and 42 ± 25 days with 1310 ± 769 locations/individual for the individuals of the other colony. There were no significant differences for the number of tracking days (t = 1.326, p = 0.202) and for the locations per individual (t = 1.629, p = 0.121) between colonies.
The average distance to the nest obtained for each individual is shown in Table 1, with maximum distances that ranged between 28.17 km (one male from the thermosolar power plant colony) and 5.37 km (one male from the colony of the towers). Some 39.73% of obtained locations were at a distance < 1 km from the nest (25.36% of locations at < 500 m from the nest); between 1–3 km there were 27.46% of them and between 3–10 km there were 31.75%. Only 1.06% of locations were recorded > 10 km from the nest. If we consider separately the locations from the thermosolar power plant colony and from the colony of the towers with nest-boxes, significant differences are observed between the frequencies of each colony (χ2 = 1801.94, df = 3, p = 0.001). Individuals from the thermosolar power plant had a higher frequency of locations between 3 and 10 km, whereas in the other colony the higher frequency was at < 1 km (Fig. 1). However, according to the results of the LMM, distances to the nest during the study period did not show significant differences in relation to the colony (Table 2). Likewise, no differences were found in relation to sex (Table 2).

Frequency of locations in relation to the distance from the nest. The results are indicated for the two studied colonies
Considering the distances before July 15, there were significant differences with the distances to the nest in relation to the colony of the individuals according to the results of the LMM (Table 2), having a greater average distance the Lesser Kestrels from the thermosolar power plant (2.79 ± 2.69 km against 1.71 ± 2.29 km of the individuals from the other colony). For distances after July 15, there were no significant differences neither in relation to the colony nor sex for any of the two time periods (Table 2).
The average size of home range areas was 13.37 km2 according to 95% kernel, 2.98 km2 for 75% kernel, 1.28 km2 for 50% kernel and 122.91 km2 for MCP (Table 1 shows the results for each individual). Although according to the LMMs, there were no significant differences for the 75% and 50% kernels areas and MCP in relation to colony and to sex, the differences were significant for the 95% kernel areas in relation to colony (Table 2): the individuals from the thermosolar power plant colony used a larger area (22.03 ± 4.07 km2; Fig. 2a, b) than those from the other colony (9.66 ± 7.68 km2; Fig. 2c, d). The largest area (according to 95% kernel) registered was 27.11 km2 (one male from the thermosolar power plant) and the smallest was 1.67 km2 (one female from the towers with nest-boxes).

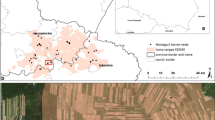
Home ranges of Lesser Kestrels tracked by GPS satellite telemetry during the breeding season. The minimum convex polygon and the 95% kernel are shown. Upper panels: individuals from the thermosolar power plant colony: a males, b females. Lower panels: individuals from the towers colony: c males, d females. The habitat type was obtained from CORINE 2012 land cover map
According to the land cover within the combined MCP, there were differences in the habitat availability between the two colonies (\(\chi_{10}^{2}\) = 3154.24, p < 0.001), although the differences were in the frequencies of each habitat type and not in the categories (Table 3). In both colonies, the non-irrigated arable lands was the most abundant habitat, which was used more frequently than expected from their availability by birds from the thermosolar power plant and from the colony of the towers (71.35% and 51.99% of locations respectively; Table 3). Lesser Kestrels also showed preference for pastures and for the surfaces where the colony was located: an artificial area (the farmhouse) and scrub and/or herbaceous vegetation associations (in this category are included the slopes of dry reservoir that were revegetated especially to imitate areas of scrub and fallow). In contrast, vineyards and arboreal vegetation were avoided (Table 3). Only 13.2% of all location were within the ‘Los Alhorines’ SPA.
Discussion
Two are the main results of the study that we comment here: the home range size of Lesser Kestrel in this colonies of eastern Spain and the possible effects that the anthropization (the thermosolar power plant) of the landscape could cause in the spatial ecology of this species, since the type of habitat that surrounds the colonies of Lesser Kestrel affects its home range and this is of great importance for its conservation (Tella et al. 1998; De Frutos et al. 2010). The largest home ranges used by birds from the thermosolar power plant colony, could be due to the influence that the infrastructure has on this colony, in combination with the anthropized environment where the plant is and the differences in habitat cover, with less availability of suitable patches for foraging. When they are in a favourable habitat, the flight distances are lower (Bustamante 1997; Tella et al. 1998; Catry et al. 2013), and in our study the frequency of locations at greater distances from the nest is higher in the thermosolar power plant colony, with also greater distances than those of the individuals from the other colony (located in a natural landscape) before July 15, when they need more quantity of preys to feed the chicks. Coinciding with Vlachos et al. (2015), we have not observed differences in the foraging areas and distances to the nest between sexes, although other studies have found a spatial segregation, with males foraging closer to the colony than females (Hernández-Pliego et al. 2017).
Despite these differences between the colony located beside the thermosolar power plant and the other in a natural environment, both the home ranges of one and another are smaller than or similar to those observed in other European colonies. Using the same methodology and considering the 95% kernel as a representative approach to area size, Gustin et al. (2017) recorded in Italy home ranges of 41.4 and 46.5 km2 in two colonies with a partially suitable landscape composed of non-irrigated arable lands and pseudo-steppes. There are other studies that use different methods to estimate the home ranges, so they are not directly comparable, but they give an idea of the size of the areas. Vlachos et al. (2015) observed home ranges over 25 km2 for males and 17 km2 (95% MCP) for females in an intensively cultivated area of central Greece. In Portugal the estimated home ranges in pseudo-steppes characterized by traditional extensive cereal cultivation were lower than 20 km2, whereas they were significant larger, around 144 km2, in unsuitable habitat with abandoned cultivations and forested areas (Catry et al. 2013). These differences according to the landscape have been also reported in colonies in Spain, with home ranges of 12.3 km2 in traditional agro-grazing systems and 63.6 km2 in an intensively cultivated area (Tella et al. 1998). The estimated home range sizes in our study area (on average 22.03 and 9.66 km2) suggest that despite the impact and proximity of the thermosolar power plant, Lesser Kestrels continue to have adequate habitats and a good availability of prey, showing the possible coexistence of new human infrastructures and Lesser Kestrels. As a compensatory measure of the environmental impact caused by the thermosolar power plant, the owner company bought the surrounding territories to guarantee that they maintained the traditional cultivation of cereals and fallows, thus maintaining the habitat used by the species. However, this positive situation may change with the replacement of grain fields and changes in land use that are occurring in the region, and the consequent reduction of suitable habitats. The Lesser Kestrels of these colonies preferably use non-irrigated arable land and pastures, where they find a large number of insects, but many of these are being replaced by vineyards. These changes in land use alter the habitat and the types of prey that can be found (Ursúa et al. 2005; García et al. 2006; Catry et al. 2012, 2014) and in addition, the vineyards imply an added danger due to the increasingly used cultivation technique in espalier, with wires to hold the vines against which the Lesser Kestrels and other birds can collide. Therefore, managing these lands around the colonies is of great importance for the conservation of this species (Donázar et al. 1993; Bustamante 1997; De Frutos et al. 2010).
Birds of prey usually select the more profitable areas as foraging habitats based on the availability and/or accessibility of their main prey items (see Village 1982; Cody 1985). For aerial hunters, such as Lesser Kestrels, the access to prey may be affected by vegetative cover of the habitat (Shrubb 1980; Bechard 1982; Toland 1987; Smallwood 1988). Consequently, capture success by aerial hunters should be favored in sites in which access to prey depends not only on its abundance but also on certain vegetation structure parameters (Rodríguez et al. 2014), apart from the type of hunting behaviour preceding the attack (Vlachos et al. 2003). Therefore, the combination of both factors (prey abundance and vegetation structure) may determine the preference for certain types of habitat. This highlights the importance of conserving the suitable habitats for the species in an increasingly anthropized landscape (Franco and Sutherland 2004; García et al. 2006).
The small percentage of locations within the ‘Los Alhorines’ SPA indicates that it is necessary to modify its extension. The criteria used to delimit the boundaries of SPAs are frequently unclear and sometimes potentially inappropriate, using for example the nest distribution with a certain boundary around the nest (Guixé and Arroyo 2011). However, management for conservation of a species should take into account its foraging needs as well as its nesting habitat (Martin and Possingham 2005).
Conclusions
In summary, although there are differences in the home ranges between the two colonies, the sizes are smaller than or similar to those observed in other European colonies. This suggests that Lesser Kestrels continue to have adequate habitats and a good availability of prey, despite the proximity and extension of the thermosolar power plant. In conclusion, we can say that the impact of an infrastructure can be minimized if special measures are taken to maintain the rest of the territory in good condition, as it has been done here. In many cases the impact of the work itself is seen in a restricted area and the environment is neglected, producing negative effects that could be alleviated.
References
Alberdi M. Actuaciones del plan de acción para la conservación de las aves de las estepas cerealistas de la Comunidad Valenciana. Informe inédito. Equipo de Seguimiento de Fauna–VAERSA. Valencia: Consellerıa de Territorio y Vivienda; 2006.
Bechard MJ. Effect of vegetative cover on foraging site selection by Swainson’s Hawk. Condor. 1982;84:153–9.
BirdLife International. Species factsheet: Falco naumanni. 2018. http://www.birdlife.org. Accessed 05 Mar 2018.
Bustamante J. Predictive models for Lesser Kestrel Falco naumanni distribution, abundance and extinction in southern Spain. Biol Conserv. 1997;80:153–60.
Catry I, Franco AMA, Sutherland WJ. Landscape and weather determinants of prey availability: implications for the Lesser Kestrel Falco naumanni. Ibis. 2012;154:111–23.
Catry I, Franco AMA, Rocha P, Alcazar R, Reis S, Cordeiro A, Ventim R, Teodósio J, Moreira F. Foraging habitat quality constrains effectiveness of artificial nest site provisioning in reversing population declines in a colonial cavity nester. PLoS ONE. 2013;8:e58320.
Catry I, Franco AMA, Moreira F. Easy but ephemeral food: exploring the trade-offs of agricultural practices in the foraging decisions of Lesser Kestrels on farmland. Bird Study. 2014;61:447–56.
Climate Data. Villena. 2018. https://es.climate-data.org/europe/espana/comunidad-valenciana/villena-57249/. Accessed 25 Oct 2018.
Cody M. Habitat selection in birds. London: Academic Press; 1985.
Cramp S, Simmons KEL. Handbook of the birds of Europe, the Middle East and North Africa; The birds of the Western Palaearctic. New York: Oxford University Press; 1980.
Clere E, Bretagnolle V. Food availability for birds in farmland habitats: biomass and diversity of arthropods by pitfall trapping technique. Rev Ecol. 2001;56:275–97.
De Frutos A, Olea PP, Mateo-Tomas P, Purroy FJ. The role of fallow in habitat use by the Lesser Kestrel during the post-fledging period: inferring potential conservation implications from the abolition of obligatory set-aside. Eur J Wildl Res. 2010;56:503–11.
Di Maggio R, Campobello D, Tavecchia G, Sarà M. Habitat- and density-dependent demography of a colonial raptor in Mediterranean agro-ecosystems. Biol Conserv. 2016;193:116–23.
Donázar JA, Negro JJ, Hiraldo F. Foraging habitat selection, land-use changes and population decline in the Lesser Kestrel Falco naumanni. J Appl Ecol. 1993;30:515–22.
European Commission. Halting the loss of biodiversity by 2010 and beyond: sustaining ecosystem services for human well-being. 2006. http://ec.europa.eu/environment/nature/biodiversity/comm2006/bap_2006.htm. Accessed 05 Mar 2018.
Franco A, Andrada J. Alimentación y selección de presa en Falco naumanni. Ardeola. 1977;23:137–87.
Franco A, Sutherland WJ. Modelling the foraging habitat selection of Lesser Kestrels: conservation implications of European Agricultural Policies. Biol Conserv. 2004;120:63–74.
García J. Dispersión premigratoria del cernícalo primilla Falco naumanni en España. Ardeola. 2000;47:197–202.
García J, Morales MB, Martinez J, Iglesias L, De La Morena E, Suárez F, Viñuela J. Foraging activity and use of space by Lesser Kestrel Falco naumanni in relation to agrarian management in central Spain. Bird Conserv Int. 2006;16:83–95.
Guixé D, Arroyo B. Appropriateness of special protection areas for wideranging species: the importance of scale and protecting foraging, not just nesting habitats. Anim Conserv. 2011;14:391–9.
Gustin M, Ferrarini A, Giglio G, Pellegrino SC, Frassanito A. Detected foraging strategies and consequent conservation policies of the Lesser Kestrel Falco naumanni in Southern Italy. Proc Int Acad Ecol Environ Sci. 2014;4:148–61.
Gustin M, Giglio G, Pellegrino SC, Frassanito A, Ferrarini A. Space use and flight attributes of breeding Lesser Kestrels Falco naumanni revealed by GPS tracking. Bird Study. 2017;64:274–7.
Hernández-Pliego J, Rodríguez C, Bustamante J. A few long versus many short foraging trips: different foraging strategies of Lesser Kestrel sexes during breeding. Mov Ecol. 2017;5:8.
Hiraldo F, Negro JJ, Donazar JA, Gaona P. A demographic model for a population of the endangered Lesser Kestrel in southern Spain. J Appl Ecol. 1996;33:1085–93.
Hood GM. PopTools version 3.2.5. 2010. http://www.poptools.org.
Hooge PN, Eichenlaub B. Animal movement extension for ArcView. U.S.A.: Alaska Science Center, Biological Science Office, U.S. Geological Survey, Anchorage, AK; 1997.
Kenward RE. A manual for wildlife radio tagging. London: Academic Press; 2001.
Kotsonas E, Bakaloudis D, Papakosta M, Goutner V, Chatzinikos E, Vlachos C. Assessment of nestling diet and provisioning rate by two methods in the Lesser Kestrel Falco naumanni. Acta Ornithol. 2018;52:149–56.
Lepley M, Brun L, Foucart A, Pilard P. Régime et comportament alimentaires du Faucon Crécerellette Falco naumanni, en Crau en période de reproduction et post-reproduction. Alauda. 2000;68:177–84.
Limiñana R, Soutullo A, Arroyo B, Urios V. Protected areas do not fulfil the wintering habitat needs of the trans-Saharan migratory Montagu’s harrier. Biol Conserv. 2011;145:62–9.
Limiñana R, Romero M, Mellone U, Urios V. Mapping the migratory routes and wintering areas of Lesser Kestrels Falco naumanni: new insights from satellite telemetry. Ibis. 2012;154:389–99.
López-López P, de La Puente J, Mellone U, Bermejo A, Urios V. Spatial ecology and habitat use of adult Booted Eagles (Aquila pennata) during the breeding season: implications for conservation. J Ornithol. 2016;157:981–93.
Manly BFJ. Randomization, bootstrap and Monte Carlo methods in biology. 2nd ed. London: Chapman & Hall; 1997.
Martin TG, Possingham HP. Predicting the impact of livestock grazing on birds using foraging height data. J Appl Ecol. 2005;42:400–8.
Martínez C. Habitat selection by the Little Bustard Tetrax tetrax in cultivated areas of central Spain. Biol Conserv. 1994;67:125–8.
Mellone U, Yáñez B, Limiñana R, Muñoz AR, Pavón D, González JM, Urios V, Ferrer M. Summer staging areas of non-breeding short-toed snake eagles. Bird Study. 2011;58:516–21.
Moreira F. Relationships between vegetation structure and breeding bird densities in fallow cereal steppes in Castro Verde, Portugal. Bird Study. 1999;46:309–18.
Negro JJ. Falco naumanni Lesser Kestrel. In: Snow DW, Perrins CM, editors. The birds of Western Palearctic: update. Oxford: Oxford University Press; 1997. p. 49–56.
Negro JJ, Hiraldo F. Nest-site selection and breeding success in the Lesser Kestrel Falco naumanni. Bird Study. 1993;40:115–9.
Rocha PA. Dieta e comportamento alimentar do Peneireiro-de-dorso-liso Falco naumanni. Airo. 1998;9:40–7.
Rodríguez C, Johst K, Bustamante J. How do crop types influence breeding success in Lesser Kestrels through prey quality and availability? A modelling approach. J Appl Ecol. 2006;43:587–97.
Rodríguez C, Tapia L, Ribeiro E, Bustamante J. Crop vegetation structure is more important than crop type in determining where Lesser Kestrels forage. Bird Conserv Int. 2014;24:438–52.
Shrubb M. Farming influences on the food and hunting of kestrels. Bird Study. 1980;27:109–15.
Silverman BW. Density estimation for statistics and data analysis. London: Chapman and Hall; 1986.
Smallwood JA. The relationship of vegetative cover to daily rhythms of prey consumption by american kestrels wintering in southcentral Florida. J Raptor Res. 1988;22:77–80.
Soutullo A, Urios V, Ferrer M, López-López P. Habitat use by juvenile Golden Eagles Aquila chrysaetos in Spain. Bird Study. 2008;55:236–40.
Tella JL, Hiraldo F, Donázar JA, Negro JJ. Costs and benefits of urban nesting in the Lesser Kestrel. In: Bird D, Varland D, Negro JJ, editors. Raptors in human landscapes: adaptions to built and cultivated environment. London: Academic Press; 1996. p. 53–60.
Tella JL, Forero MG, Hiraldo F, Donázar JA. Conflicts between Lesser Kestrel conservation and European agricultural policies as identified by habitat use analyses. Conserv Biol. 1998;12:593–604.
Tellería JL, Santos T, Álvarez G, Sáez-Royuela C. Avifauna de los campos de cereales del interior de España. In: Bernis F, editor. Aves de los medios urbano y agrícola en las mesetas españolas. Madrid: SEO; 1988. p. 173–319.
Toland BR. The effect of vegetative cover on foraging strategies, hunting success and nesting distribution of American kestrels in central Missouri. J Raptor Res. 1987;21:14–20.
Tucker GM, Heath MF. Birds in Europe: their conservation status. Conservation Series 3. Cambridge: BirdLife International; 1994.
Ursúa E, Serrano D, Tella JL. Does land irrigation actually reduce foraging habitat for breeding Lesser Kestrels? The role of crop types. Biol Conserv. 2005;122:643–8.
Village A. The home range and density of kestrels in relation to vole abundance. J Anim Ecol. 1982;51:413–28.
Vlachos CG, Bakaloudis DE, Chatzinikos E, Papadopoulos T, Tsalagas D. Aerial hunting behaviour of the Lesser Kestrel Falco naumanni during the breeding season in Thessaly (Greece). Acta Ornithol. 2003;38:129–34.
Vlachos CG, Bakaloudis DE, Kitikidou K, Goutner V, Bontzorlos V, Papakosta MA, Chatzinikos E. Home range and foraging habitat selection by breeding Lesser Kestrels (Falco naumanni) in Greece. J Nat Hist. 2015;49:371–81.
Wiens JA. Habitat selection in variable environments: shrub-steppe birds. In: Cody ML, editor. Habitat selection in birds. Orlando: Academic Press; 1985. p. 227–51.
Wiens JA. The ecology of bird communities. Cambridge: Cambridge University Press; 1989.
Worton BJ. Kernel methods for estimating the utilization distribution in home-range studies. Ecology. 1989;70:164–8.
Authors’ contributions
VU designed the experiment and MR performed the fieldwork and collected the data. JV and MR performed data analyses. All authors contributed to writing. All authors read and approved the final manuscript.
Acknowledgements
We are grateful to Toni Pérez for his help during all the fieldwork and along all the time. We are thankful to Servicio de Biodiversidad of Conselleria de Agricultura, Medio Ambiente, Cambio Climático y Desarrollo Rural (Generalitat Valenciana), especially to Juan Jiménez and Juan Antonio Gómez for giving all the necessary permissions. We are also thankful to Ruben Limiñana for his advices. This paper is part of the PhD thesis of M. Romero at the University of Alicante.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets analysed during the current study are available from the Movebank database (Lesser Kestrel project).
Consent for publication
Not applicable.
Ethics approval
This study was approved by the Regional Ministry of Agriculture, Environment, Climate Change and Rural Development of Comunidad Valenciana, and the permissions for the capture and tagging were also granted by this organization. Birds were handled by qualified personnel and anesthesia was not necessary.
Funding
Funding for the project on studying the spatial ecology of Lesser Kestrel was provided by Enerstar S.A. (http://www.enerstar.es). JV is supported by a FPU grant of Spanish Ministry of Education (reference FPU014/04671). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Vidal-Mateo, J., Romero, M. & Urios, V. How can the home range of the Lesser Kestrel be affected by a large civil infrastructure?. Avian Res 10, 10 (2019). https://doi.org/10.1186/s40657-019-0149-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40657-019-0149-6