
Abstract
Background
Anemia of inflammation (AI) is common in critically ill patients. Although this syndrome negatively impacts the outcome of critical illness, understanding of its pathophysiology is limited. Also, new therapies that increase iron availability for erythropoiesis during AI are upcoming. A model of AI induced by bacterial infections that are relevant for the critically ill is currently not available. This paper describes the development of an animal model for AI that is relevant for critical care research.
Results
In experiments with rats, the rats were inoculated either repeatedly or with a slow release of Streptococcus pneumoniae or Pseudomonas aeruginosa. Rats became ill, but their hemoglobin levels remained stable. The use of a higher dose of bacteria resulted in a lethal model. Then, we turned to a model with longer disease duration, using pigs that were supported by mechanical ventilation after inoculation with P. aeruginosa. The pigs became septic 12 to 24 h after inoculation, with a statistically significant decrease in mean arterial pressure and base excess, while heart rate tended to increase. Pigs needed resuscitation and vasopressor therapy to maintain a mean arterial pressure > 60 mmHg. After 72 h, the pigs developed anemia (baseline 9.9 g/dl vs. 72 h, 7.6 g/dl, p = 0.01), characterized by statistically significant decreased iron levels, decreased transferrin saturation, and increased ferritin. Hepcidin levels tended to increase and transferrin levels tended to decrease.
Conclusions
Using pathogens commonly involved in pulmonary sepsis, AI could not be induced in rats. Conversely, in pigs, P. aeruginosa induced pulmonary sepsis with concomitant AI. This AI model can be applied to study the pathophysiology of AI in the critically ill and to investigate the effectivity and toxicity of new therapies that aim to increase iron availability.
Similar content being viewed by others


Background
Anemia is a hallmark of critical illness, and inflammation is thought to contribute to the development of anemia in the majority of critically ill patients. Thereby, anemia of inflammation (AI) is common in the intensive care unit (ICU) [1]. The pathophysiology of AI is multifactorial and includes a shortened red blood cell (RBC) life span caused by erythrophagocytosis [2], as well as a decreased erythropoiesis [3]. Erythropoiesis is impaired due to inflammatory cytokines that decrease erythroid precursor proliferation and erythropoietin levels [4]. Also, serum iron levels are low in AI due to increased levels of hepcidin, which is the iron-regulating hormone that is produced in response to inflammatory cytokines [5]. Hepcidin causes degradation of the iron exporter ferroportin, resulting in sequestration of iron inside cells and subsequent low plasma iron [6]. Thereby, in contrast to iron deficiency anemia, AI patients do not have a lack of iron, but rather a decreased iron availability.
Regardless of the cause, anemia in the critically ill is associated with adverse outcome [7] and AI occurs early after ICU admission [8]. AI is treated with RBC transfusions. In sepsis, half of the patients require RBC transfusion within the first 24 h of ICU admission [8]. However, RBC transfusion is associated with morbidity and mortality in the critically ill [9], calling for alternative therapies. In the last decade, new therapies to treat AI by targeting the iron metabolism are in development, such as hepcidin inactivators [10,11,12], hepcidin production inhibitors [13, 14], interleukin 6 (IL-6) inhibitor [15], and IL-6 receptor blockers [16,17,18]. These therapies aim to increase the amount of iron available for erythropoiesis, which could potentially reduce the amount of transfusions and improve the outcome of critical illness. However, such interventions may also have drawbacks, such as slow resolution of infection or acquisition of new infections, as bacteria use iron for their growth [19]. Therefore, new therapies for critically ill patients that increase iron availability should preferably be tested in models of AI that are caused by bacterial infections that are relevant for the critically ill. The currently available AI animal models are mainly non-infectious models, including heat-killed Brucella abortus [2], zymosan [20], cytokines [11, 21], or peptidoglycan-polysaccharide [22]. Infectious AI animal models show high variation due to technical difficulties [23, 24], have only mild anemia [25, 26], or use parasitic infections that are not relevant for the critically ill [27,28,29,30]. Taken together, animal models for AI that mimic critical care illness are currently limited. Therefore, through a multi-national European collaboration, we aimed at developing an animal model of AI caused by pulmonary sepsis, evaluating the advantages and disadvantages of models in rats and pigs.
Methods
Rat experiments
Studies were approved by the Institutional Animal Care and Use Committee of the Amsterdam University Medical Centers, located at the Academic Medical Center, Amsterdam, Netherlands. All animal procedures were performed in compliance with Institutional Standards for Human Care and Use of Animal Laboratory Animals.
Experimental protocol
For all experiments, the bacterial inoculum was prepared as follows: an overnight bacterial culture was diluted in fresh medium. The bacteria were cultured to logarithmic growth phase at 37 °C. Then, the culture was centrifuged and the pellet was washed and resuspended in sterile saline. This bacterial suspension was diluted to the desired inoculum concentration, based on the optical density at 600 nm. The inoculum concentration was verified by the culture of 10-fold serial dilutions of the inoculum on agar plates.
Male Sprague-Dawley rats (Envigo, The Netherlands) were anesthetized with 3% isoflurane. After baseline blood sampling via a catheter in the tail vein, rats were intratracheally inoculated with a high dose of 108 colony-forming units (CFU) of a log-phase culture of Streptococcus pneumoniae serotype 3 (ATCC 6303; Rockville, MD, USA) in a volume of 150 μL using a miniature nebulizer. A second group of rats was inoculated repeatedly, at day 0 and day 4, with 107 CFU of a log-phase culture of S. pneumoniae in a volume of 150 μL. A third group of rats was inoculated with 106 CFU of a log-phase culture of S. pneumoniae embedded in agar beads in a volume of 150 μL, from which bacteria are slowly released [31] (Fig. 1). These S. pneumoniae agar beads were prepared as described in the Additional file 1. Finally, additional rats were inoculated with high dose Pseudomonas aeruginosa (PA103, 109 CFU; kindly provided by Iglewski Laboratory, Rochester, NY, USA) in a volume of 150 μL either in a bolus solution or via slow release, embedded in agar beads.

Streptococcus pneumoniae embedded in an agar bead. Richardson staining, × 400 magnification. Agar beads with Streptococcus pneumonia have a diameter of 50–200 μm. The size of these S. pneumonia beads is similar to the Pseudomonas aeruginosa agar beads in the original protocol [31].
All rats were weighed daily. Supplemental fluid bolus (10 ml/kg Ringers Lactate) was given every 24 h intraperitoneally in case of severe illness, as indicated by > 10% weight loss compared to the previous day. Blood samples of 500 μl were taken via the tail vein at baseline and at day 3, 7, and 10 to measure the hemoglobin (Hb) level. No control group was used in these experiments. Hb levels were compared with the baseline measurement. After 7 or 14 days, rats were anesthetized by intraperitoneal injection of 90 mg/kg ketamine (Dechra, The Netherlands), 0.25 mg/kg dexmedetomidine (Orion Pharrma, Finland), and 0.5 mg/kg atropine (Centrafarm, The Netherlands) to bleed them via the inferior caval vein. The blood was collected into EDTA anti-coagulated tubes
Sample analysis
Hb levels were measured using the ScilVet abc + (scil animal care company GmbH, Germany). Of each rat, the two middles lobes of the right lung were homogenized in 1 ml sterile PBS. Serial 10-fold dilutions were plated on blood agar plates and incubated at 37 °C with 5% CO2. The number of CFUs was counted the next day.
Pig experiment
This study was approved by the Institutional Review Board and Animal Ethics Committee of the University of Barcelona, Barcelona, Spain. The animal procedures were performed according to local Spanish guidelines for the use and care of animals.
Experimental protocol
A porcine ventilator-associated pulmonary sepsis model was used, as has been described before [32]. Five female large-white Landrace pigs of 31 ± 1.3 kg (range 29–32 kg) were used. Pigs were premedicated with intramuscular 2 mg/kg azaperone. Then, animals were induced with 2–2.5 mg/kg of propofol, orotracheally intubated with a 7.5-mm I.D. endotracheal tube and mechanically ventilated with a SERVO-i (Maquet, Wayne, NJ, USA). Pigs were ventilated in volume-control setting, with a tidal volume of 10 ml/kg, inspiratory fraction of oxygen 40%, and without positive end-expiratory pressure. Respiratory rate was adjusted every 6 h to maintain normocapnia (40–45 mmHg PaCO2). An arterial line was inserted into the femoral artery to monitor systemic arterial pressure and to collect blood samples. A central venous catheter was surgically placed in the jugular vein for intravenously fluid and drug administration. A no. 8 Foley catheter was placed into the bladder through surgical mini-pelvectomy. Ceftriaxon (1 g) was administrated intravenously 30 min before intubation and 50 mg/kg was given every 12 h to prevent pneumonia caused by oropharyngeal flora. Following surgical preparation and stabilization—approximately 4 h after intubation—the pigs were bronchoscopically inoculated with 15 mL of 108 CFU of a log-phase culture of Pseudomonas aeruginosa (subtype ATCC 27853, ceftriaxone resistant) into each lobe. The inoculum was prepared as described above. Ringer lactate and 0.9% NaCl solution were infused to maintain fluid balance. The animals received 2 l of 0.9% NaCl daily. Also, boluses of 20 ml/kg were given when the pig was thought to be fluid responsive. The fluid therapy targeted urine output ≥ 0.5 m/kg/h. Blood was drawn via the arterial line into EDTA anti-coagulated and serum separating tubes (BD™ Vacutainer™ SST™ II Advance Tubes). Samples were centrifuged for 10 min, at 1750×g and the serum was stored at − 80 °C. Blood samples were taken at baseline (just prior to inoculation) and every 12 h thereafter, until 72 h. Bronchoalveolar lavage (BAL) was performed bronchoscopically with 40 ml saline in the right medium lobe at baseline, 24 and 72 h thereafter. The animals were euthanized 72 h after intubation using intravenous overdosing propofol, potassium.
Blood sampling and analysis
Full blood count and hemoglobin was assessed every 24 h through a hematocytometer (Siemens Advia 2021i, Erlangen, Germany). At the same time points, serum iron was measured using a colorimetric assay (Sekisui Diagnostics, Lexington, MA). Total iron-binding capacity (TIBC) was measured using a colorimetric assay (Pointe Scientific). Transferrin levels were calculated using the formula TIBC (μmol/L)/25.1. Transferrin saturation was calculated with the formula serum iron/(25.1 × total transferrin) × 100%. Ferritin (LsBio), hepcidin (LsBio), and interleukin 6 (IL-6) levels (Millipore Iberica, S.A., Madrid, Spain) were measured by enzyme-linked immunosorbent assay kits. Lung tissue was homogenized and cultured on agar plates at 37 °C with 5% CO2. The number of CFUs was counted the next day.
Statistical analysis
The data is expressed as median ± IQR. Differences between time points were analyzed using a paired samples t test or a Wilcoxon signed-ranks test, for normally and not normally distributed parameters, respectively. Statistical significance was considered to be at p ≤ 0.05. All statistical analyses were performed using IBM SPSS Statistics 24.
Results
Rat experiments
Streptococcus pneumoniae pneumonia in rats does not cause AI
Rats infected with a bolus dose of 108 CFU S. Pneumoniae appeared very ill, with 10% weight loss and a ruffled fur at 24 h after inoculation. The animals died the second day, prior to the development of anemia. Rats inoculated with 107 CFU S. Pneumoniae did not show a decrease in weight. At day 4, these rats were inoculated again with 107 CFU S. Pneumoniae, and from then on, the animals started to lose weight (Fig. 2a). At day 7, the rats appeared ill, with lethargic behavior and ruffled fur. The animals showed macroscopic pneumonia, and there were still bacteria present in the lungs, indicating that the infection was not cleared (Fig. 2b). However, Hb levels were not decreased at 7 days compared to baseline (Fig. 2c). Next, an approach was taken using bacteria embedded in beads, with the aim to induce a slow releasing reservoir resulting in prolonged unresolved infection. Rats inoculated with 106 CFU S. pneumoniae embedded in agar beads did not lose weight during the experiment, but even seemed to gain weight (Fig. 2d). At 14 days, macroscopic lesions were visible in the lungs and there were still bacteria present in the lungs (Fig. 2e). However, Hb levels did not decrease during the 14 days experiment (Fig. 2c). In addition, rats inoculated with 109 CFU P. aeruginosa or 109 CFU P. aeruginosa embedded in agar beads did not show any decrease in Hb level either (data not shown).

Failed attempts to establish rat AI model using S. pneumoniae. Daily weight (a), lung CFU/ml after 7 days (b), and Hb levels (c) of rats inoculated with a log-phase culture of 107 CFU S. pneumoniae at day 0 and day 4. The daily weight (d), lung CFU/ml (e), and Hb levels (f) of rats inoculated with a log-phase culture of 106 CFU S. pneumoniae embedded in agar beads. N = 4 and data are expressed as mean + SD (n = 4). ***P < 0.001, **P < 0.01, *P < 0.05
Pig experiments
Pseudomonas aeruginosa causes pneumonia and shock in pigs
Between 12 and 24 h after inoculation, pigs presented signs of severe respiratory infection. Table 1 shows the hemodynamic parameters of the pigs at several time points during the experiment. The mean arterial pressure decreased over time, from 80 mmHg (76–85) at baseline to 67 mmHg (63–71) at 24 h (p = 0.02), while the heart rate (p = 0.07) and noradrenaline infusion rate (p = 0.07) tended to increase in the first 24 h. Arterial base excess decreased between baseline and 12 h, from 7.6 mmol/L (5.7–9.7) to 2.8 mmol/L (− 1.1–6.4) (p = 0.03) (Table 1). Also, platelets decreased in the first 24 h from 338 × 109/L (330–568) to 205 × 109/L (123–219) (p = 0.004) and remained low throughout the experiment. White blood cell (WBC) counts tended to increase over time, from 8.8 × 109/L (6.5–11.6) at baseline to 23.8 × 109/L (12.8–29.9) at 48 h (p = 0.07). CFU counts increased during the experiment, from 0 at baseline until 4.6 log CFU/ml (3.7–5.9) (p = 0.04) in BAL fluid at 72 h (Fig. 3).

Characteristics of a pig pneumosepsis model. Hb levels (a), WBC counts (b), platelet counts (c), and P. aeruginosa concentration in the BAL fluid (d) in pigs inoculated with 15 mL of 108 CFU of a log-phase culture of P. aeruginosa into each lung lobe. N = 5 and data are expressed as mean + SD (n = 5). ***P < 0.001, **P < 0.01, *P < 0.05
Pseudomonas aeruginosa pneumonia causes AI in pigs
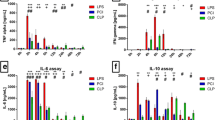
Hb levels at baseline were 9.9 g/dl (9.2–10.7) and decreased significantly over time to 7.6 g/dl (7.1–7.7) after 72 h (p = 0.01) (Fig. 3). Serum iron level at baseline was already lower than normal for pigs (9.5 μmol/L (6.8–14.4)) and decreased significantly throughout the experiment to 3.5 μmol/L (1.9–4.1) after 72 h (p = 0.01). Also, transferrin saturation decreased significantly over time from 10.4% (8.6–13.0) at baseline to 4.7% (2.9–6.0) after 72 h (p < 0.01). Transferrin levels tended to decrease during the experiment from 3.6 g/L (3.0–4.7) at baseline to 2.6 g/L (2.5–2.8) at T = 72 h (p = 0.08). Ferritin levels increased significantly from 25.2 ng/ml (21.1–36.4) at baseline to 188.6 ng/ml (178.0–207.8) after 72 h (p = 0.04). Hepcidin levels slightly increased during the experiment from 151.2 ng/ml (110.1–185.5) at baseline to 359.9 ng/ml (138.8–476.2) after 48 h (p = 0.08). Finally, IL-6 increased significantly from 31 pg/ml (17–241) to 2698 pg/ml (1713–4180) at T = 24 h (p = 0.04) (Fig. 4).

Iron parameters in pigs with pneumosepsis. Levels of iron (a), ferritin (b), transferrin (c), transferrin saturation (d), hepcidin (e), and IL-6 (f) in pigs inoculated with 15 mL of 108 CFU of a log-phase culture of P. aeruginosa into each lung lobe. N = 5 and data are expressed as mean + SD (n = 5). ***P < 0.001, **P < 0.01, *P < 0.05. Dotted lines represent reference levels
Discussion
Initially, the use of rodents was evaluated for the development of this AI model, because of the lower costs and because of ample experience with these animals. However, efforts to develop a rat model for AI due to S. pneumoniae or P. aeruginosa pneumonia were unsuccessful. The rats did not survive inoculation of high dose bacteria, whereas a lower, repeated dose of bacteria or slow-release infection with bacteria embedded in agar beads did not result in AI, despite the presence of unresolved infection at day 14. This suggests that it is not feasible to induce AI due to prolonged severe infection in rats with the use of pathogens commonly encountered in the critically ill. Consequently, we switched to an existing pig model. This model is a model of longer lasting pneumosepsis induced by P. aeruginosa and has been detailed elsewhere [32]. However, the anemia status in this model has never been described.
We found that prior to illness induction, the pigs have low iron and ferritin levels, compatible with iron deficiency at baseline. However, the pigs developed anemia over time during the sepsis. The decrease in Hb was due to AI, exemplified by hypoferremia, decreased transferrin saturation, and increased ferritin levels, associated with an increased IL-6 and hepcidin levels. Also, serum transferrin levels tended to decrease due to the inflammation. Taken together, these pigs show all the characteristics of AI [33]. The main strength of this model is that it is very similar to the ICU setting since the pigs had severe pneumosepsis, requiring mechanical ventilation and hemodynamic support. The animals developed kidney injury and acute respiratory distress syndrome showing that this is a model for septic shock with multi-organ failure [32, 34, 35]. Other strengths of this model are the comparability to AI in humans and the possibility of a long-term study. Future animal studies could use this pig AI model to pre-test the efficacy and toxicity before the start of trials with new agents for AI in the critically ill. Also, as the cardiac output of pigs almost equals that of humans [36], dose-finding medication studies can be done in this model as well. However, as the pigs that we used had a mild iron-deficiency at baseline, we would recommend to correct this iron deficiency by an intramuscular iron injection for studies to AI [37]. Further, compared to patients on the ICU that receive mechanical ventilation for weeks, this model is still a short-term model.
Conclusions
This study describes an AI model relevant for critical care. This model can be used to study the pathophysiology of AI during critically illness as well as the effect of new therapies for AI that aim to increase iron availability. Using this model, both the effect on anemia as well as the effect on immune host response, bacterial resolution, and organ failure can be studied.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- AI:
-
Anemia of inflammation
- BAL:
-
Bronchoalveolar lavage
- BE:
-
Base excess
- CFU:
-
Colony-forming unit
- Hb:
-
Hemoglobin
- HR:
-
Heart rate
- ICU:
-
Intensive care unit
- IL-6:
-
Interleukin 6
- MAP:
-
Mean arterial pressure
- RBC:
-
Red blood cell
- TIBC:
-
Total iron-binding capacity
References
Prakash D (2012) Anemia in the ICU: anemia of chronic disease versus anemia of acute illness. Crit Care Clin 28:333–343. https://doi.org/10.1016/j.ccc.2012.04.012
Kim A, Fung E, Parikh SG, et al (2014) A mouse model of anemia of in flammation: complex pathogenesis with partial dependence on hepcidin. Blood 123:1129–1137. https://doi.org/10.1182/blood-2013-08-521419.A.K.
Sihler KC, Napolitano LM (2008) Anemia of inflammation in critically ill patients. J Intensive Care Med 23:295–302. https://doi.org/10.1177/0885066608320836
Ge C, Ma L (1946) The anemia of infection; hypoferremia, hypercupremia, and alterations in porphyrin metabolism in patients. J Clin Invest 25:65–80. https://doi.org/10.1172/JCI101690
Park CH, Valore EV, Waring a J, Ganz T (2001) Hepcidin, a urinary antimicrobial peptide synthesized in the liver. J Biol Chem 276:7806–7810. https://doi.org/10.1074/jbc.M008922200
Nemeth E, Tuttle MS, Powelson J et al (2004) Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science (80- ) 306:2090–2093. https://doi.org/10.1126/science.1103786
Hayden SJ, Albert TJ, Watkins TR, Swenson ER (2012) Anemia in critical illness: Insights into etiology, consequences, and management. Am J Respir Crit Care Med 185:1049–1057. https://doi.org/10.1164/rccm.201110-1915CI
Holst LB, Haase N, Wetterslev J et al (2014) Lower versus higher hemoglobin threshold for transfusion in septic shock. N Engl J Med 141001051021009. https://doi.org/10.1056/NEJMoa1406617
Napolitano LM, Kurek S, Luchette Fa, et al (2009) Clinical practice guideline: red blood cell transfusion in adult trauma and critical care. J Trauma 67:1439–1442. https://doi.org/10.1097/TA.0b013e3181ba7074
Sasu BJ, Cooke KS, Arvedson TL et al (2010) Antihepcidin antibody treatment modulates iron metabolism and is effective in a mouse model of inflammation-induced anemia. Blood 115:3616–3624. https://doi.org/10.1182/blood-2009-09-245977
Schwoebel F, van Eijk LT, Zboralski D et al (2013) The effects of the anti-hepcidin Spiegelmer NOX-H94 on inflammation-induced anemia in cynomolgus monkeys. Blood 121:2311–2315. https://doi.org/10.1182/blood-2012-09-456756
Van Eijk LT, John ASE, Schwoebel F et al (2014) Effect of the antihepcidin Spiegelmer lexaptepid on inflammation-induced decrease in serum iron in humans. Blood 124:2643–2646. https://doi.org/10.1182/blood-2014-03-559484
Poli M, Girelli D, Campostrini N et al (2010) Heparin: a potent inhibitor of hepcidin expression in vitro and in vivo. Blood 117:997–1004. https://doi.org/10.1182/blood-2010-06-289082
Steinbicker AU, Sachidanandan C, Vonner AJ et al (2011) Inhibition of bone morphogenetic protein signaling attenuates anemia associated with inflammation. Blood 117:4915–4924. https://doi.org/10.1182/blood-2010-10-313064
Schipperus M, Rijnbeek B, Reddy M et al (2009) CNTO328 (Anti-IL-6 mAb) treatment is associated with an increase in hemoglobin (Hb) and decrease in hepcidin levels in renal cell carcinoma (RCC). (Abstract). Blood 114:1551
Song SNJ, Tomosugi N, Kawabata H et al (2010) Down-regulation of hepcidin resulting from long-term treatment with an anti-IL-6 receptor antibody (tocilizumab) improves anemia of inflammation in multicentric Castleman disease. Blood 116:3627–3634. https://doi.org/10.1182/blood-2010-03-271791
Song S-NJ, Iwahashi M, Tomosugi N et al (2013) Comparative evaluation of the effects of treatment with tocilizumab and TNF-a inhibitors on serum hepcidin, anemia response and disease activity in rheumatoid arthritis patients. Arthritis Res Ther 15(1). https://doi.org/10.1186/ar4323
Isaacs JD, Harari O, Kobold U et al (2013) Effect of tocilizumab on haematological markers implicates interleukin-6 signalling in the anaemia of rheumatoid arthritis. Arthritis Res Ther 15:R204. https://doi.org/10.1186/ar4397
Patruta S, Walter HH (1999) Iron and infection ¨. Kidney Int 55
Lasocki S, Millot S, Andrieu V et al (2008) Phlebotomies or erythropoietin injections allow mobilization of iron stores in a mouse model mimicking intensive care anemia. Crit Care Med 36:2388–2394. https://doi.org/10.1097/CCM.0b013e31818103b9
Libregts SF, Gutiérrez L, De BAM et al (2011) Chronic IFN- γ production in mice induces anemia by reducing erythrocyte life span and inhibiting erythropoiesis through an IRF-1 / PU . 1 axis. Blood 118:2578–2588. https://doi.org/10.1182/blood-2010-10-315218
Sartor RB, Anderle SK, Rifai N et al (1989) Protracted anemia associated with chronic, relapsing systemic inflammation induced by arthropathic peptidoglycan-polysaccharide polymers in rats. Infect Immun 57:1177–1185
Schubert T, Echtenacher B, Hofstädter F, Männel DN (2003) TNF-independent development of transient anemia of chronic disease in a mouse model of protracted septic peritonitis. Lab Investig 83:1743–1750. https://doi.org/10.1097/01.LAB.0000101693.12149.2C
Schubert TE, Echtenacher B, Hofstädter F, Männel DN (2005) Failure of interferon-gamma and tumor necrosis factor in mediating anemia of chronic disease in a mouse model of protracted septic peritonitis. Int. J. Mol. Med. 16:753–758
Gallimore B, Gagnon RF, Subang R, Richards GK (1991) Natural history of chronic staphylococcus epidermidis foreign body infection in a mouse model. J Infect Dis 164:1220–1223
Ganz SR and T (2010) Animal models of anemia of inflammation. Semin Hematol 46:351–357. https://doi.org/10.1053/j.seminhematol.2009.06.003.Animal
Emeribe AO, Anosa VO (1991) Haematology of experimental Trypanosoma brucei gambiense infection. Rev Elev Med Vet Pays Trop 44:53–57
Stijlemans B, Vankrunkelsven A, Brys L et al (2008) Role of iron homeostasis in trypanosomiasis-associated anemia. Immunobiology 213:823–835. https://doi.org/10.1016/j.imbio.2008.07.023
Thawani N, Tam M, Stevenson MM (2009) STAT6-mediated suppression of erythropoiesis in an experimental model of malarial anemia. Haematologica 94:195–204. https://doi.org/10.3324/haematol.13422
Lamb TJ, Langhorne J (2008) The severity of malarial anaemia in Plasmodium chabaudi infections of BALB/c mice is determined independently of the number of circulating parasites. Malar J 7:1–9. https://doi.org/10.1186/1475-2875-7-68
Facchini M, De Fino I, Riva C, Bragonzi A (2014) Long term chronic Pseudomonas aeruginosa airway infection in mice. J Vis Exp:1–10. https://doi.org/10.3791/51019
Gianluigi Li Bassi, M.D., Ph.D., Montserrat Rigol, D.V.M., Ph.D., Joan-Daniel Marti, R.P.T. PD, Lina Saucedo, M.D., Otavio T. Ranzani, M.D., Ignasi Roca, Ph.D., Maria Cabanas MD, Laura Muñoz, B.Sc., Valeria Giunta, M.D., Nestor Luque, M.D., Mariano Rinaudo MD, et al (2014) A novel porcine model of ventilator-associated pneumonia caused by oropharyngeal challenge with Pseudomonas aeruginosa. Anesthesiology 1205–1215
Nemeth E, Ganz T (2014) Anemia of inflammation. Hematol Oncol Clin North Am 28:671–681. https://doi.org/10.1016/j.hoc.2014.04.005
Sibila O, Agustí C, Torres a, et al (2007) Experimental Pseudomonas aeruginosa pneumonia: evaluation of the associated inflammatory response. Eur Respir J 30:1167–1172. https://doi.org/10.1183/09031936.00053607
Luna CM, Baquero S, Gando S et al (2007) Experimental severe Pseudomonas aeruginosa pneumonia and antibiotic therapy in piglets receiving mechanical ventilation. Chest 132:523–531. https://doi.org/10.1378/chest.07-0185
Lelovas PP, Kostomitsopoulos NG, Xanthos TT (2014) A comparative anatomic and physiologic overview of the porcine heart. J Am Assoc Lab Anim Sci 53:432–438
Starzyński RR, Laarakkers CMM, Tjalsma H et al (2013) Iron supplementation in suckling piglets: how to correct iron deficiency anemia without affecting plasma hepcidin levels. PLoS One 8:e64022. https://doi.org/10.1371/journal.pone.0064022
Acknowledgements
Not applicable.
About this supplement
This article has been published as part of Intensive Care Medicine Experimental Volume 7 Supplement 1 2019: Proceedings from the Third International Symposium on Acute Pulmonary Injury and Translational Research (INSPIRES III). The full contents of the supplement are available at https://icm-experimental.springeropen.com/articles/supplements/volume-7-supplement-1.
Funding
This manuscript was not sponsored for publication.
Author information
Authors and Affiliations
Contributions
The study design was contributed by MB, SZ, AT, GLB, and NJ. MB, AT, GLB, EA, AM, TS, FAI, and PP contributed to the data. MB, RB, AT, GLB, and NJ analyzed and interpreted the data. MB, RB, GLB, and NJ wrote the manuscript. All authors reviewed the manuscript and approved the final submitted version.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
These studies were approved by the Institutional Animal Care and Use Committee of the Academic Medical Center, University of Amsterdam, Amsterdam, Netherlands, and the Institutional Review Board and Animal Ethics Committee of the University of Barcelona, Barcelona, Spain.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional file
Additional file 1:
Protocol for embedding Streptococcus pneumonia in agar beads. (DOCX 14 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Boshuizen, M., van Bruggen, R., Zaat, S.A. et al. Development of a model for anemia of inflammation that is relevant to critical care. ICMx 7 (Suppl 1), 47 (2019). https://doi.org/10.1186/s40635-019-0261-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40635-019-0261-2