
Abstract
Beta-glucans, which existed in the cell walls of cereals, bacteria, and fungi, comprise a group of β-d-glucose polysaccharides. We investigated the effects of four kinds of beta-glucan, that are derived from barley, yeast, mushroom, and euglena on wound healing. The migration and viability of keratinocyte or fibroblast were analyzed using the in vitro scratch wound healing assay, invasion assay, MTT assay, and in vivo assay. All the beta-glucans had a significant effect on keratinocyte migration at 20 μM and showed no toxicity on dermal fibroblast. Moreover, treatment of keratinocytes with the beta-glucan derived from the mushroom (Schizophyllum commune) promoted in vivo wound closure. The Integrin/FAK/Src pathway is known to affect cell migration by forming lamellipodia. Beta-glucan from S. commune activates the Integrin/FAK/Src signaling pathway in a time-dependent. Reactive oxygen species are associated with fibroblast differentiation to contract dermal layer and synthesize collagens. We found that fibroblast was activated by increasing NOX4 expression. We propose that beta-glucan derived from mushroom is capable of promoting keratinocyte migration via the induction of FAK/Src phosphorylation there by accelerating wound closure and activating dermal fibroblast differentiation through NADPH oxidase for matrix remodeling.
Similar content being viewed by others


Introduction
Beta-glucan is a generic term for β-d-glucose polysaccharides, a major component of the cell walls, and classified into microbial- or plant derived-glucans depending on sources such as, bacteria, fungus, yeast, cereal [1, 2]. In general, beta-glucans have d-glucose units with β-1,3 link, but cereal beta-glucan contain both β-1,3 and β-1,4 backbone bonds. Yeast and fungus β-glucan contains long β-1,6 and short β-1,6 branching respectively, while bacterial beta-glucan have no branching [3] (Fig. 1). Also, different types of beta-glucans exhibit distinct molecular weights, solubility, and viscosity causing diverse physiological functions [4]. It is well known that beta-glucans modulate immune functions such as phagocytosis, have anti-cancer and anti-biotic properties, and lower blood pressure or cholesterol levels [5]. Since beta-glucan also induces the production of growth factors essential for skin, promotes collagen biosynthesis, and maintains skin’s moisture and elasticity, we investigated whether beta-glucan regenerates damaged skin and different types of beta-glucan have different effects.

The structure and description of four kinds of beta-glucans
Regeneration of impaired skin involves complex biological processes, such as re-epithelialization and fibroplasia. During re-epithelialization, epithelial cells surrounding wound margin grow and migrate to form normal epidermis before they start to differentiate for stratification giving rise to the basal, spinous, granular, and cornified layers. Integrin-induced FAK/Src signaling pathway was shown to regulate epithelial cell migration [6]. For fibroplasia, it is essential that dermal fibroblast proliferates and differentiates. Differentiated dermal fibroblast synthesizes extracellular matrix, such as a collagen and elastin, and induce dermal contraction [7].
In this study, we show that four types of beta-glucan promote epithelial migration and dermal activation without any toxic effects on the skin cells. In particular, beta-glucan derived from S. commune was most effective in wound repair. It accelerated re-epithelialization through integrin/FAK/Src signaling pathway and activated dermal transformation via integrin or reactive oxygen species (ROS) production.
Materials and methods
Materials
Beta-glucan derived from black yeast, barley, and euglena gracilis were respectively purchased from TCI chemicals (G0331), SIGMA (G6513), and SIGMA (89862). Beta-glucan derived from S. commune was obtained from QUEGEN biotech. These were dissolved in distilled water and adjusted to pH 7.0.
Cell culture
Human dermal fibroblasts were isolated from human foreskin specimens in this lab, and described in previous paper [8]. Human spontaneously immortalized keratinocyte cell lines (HaCaT) were obtained from the Korea Cell Line Bank. HaCaT keratinocyte and primary human dermal fibroblast were cultured in DMEM (Gibco Co.) supplemented with 10% FBS (Omega Co.) and 1% penicillin/streptomycin. Cells were incubated at 37 °C in 5% CO2.
MTT assay
Cells (2 × 104 cells/ml) were seeded in 96-well plates. Next day, cells were treated with four types of beta-glucans (0–40 μg/ml) or S. commune-derived glucan (0–0.4 mg/ml). After incubation for 24 or 48 h with HaCaT or fibroblast, 10 μl of MTT solution (5 mg/ml) was added to each well, and cells were incubated for an additional 4 h at 37 °C. The medium was replaced with 150 μl DMSO, and the plate was incubated for 30 min at room temperature with shaking. Absorbance was measured at 570 nm using a spectrophotometer [9].
Scratch wound healing assay
HaCaT cells were incubated in 48-well plates at a density of 7 × 104 cells/well for 24 h. A scratch was made on the cell monolayer by drawing a pipette tip across the well. The culture medium was supplemented with four types of beta-glucan or S. commune-derived-glucan dose-dependently. At 0 h and 24 h post-treatment, wound closure was captured using an Olympus IX70 microscope equipped with a digital camera at 40× magnification. The distance migrated was measured using the Image J software, and calculated by comparing the initial and final width of the scratch.
Matrigel invasion assay
HaCaT cells (7 × 104 cells/well) were seeded in the insert of a 12-well invasion assay kit (SPL Co.), and the bottom chamber was filled with media. The membrane allows the exchange of media during incubation. After 24 h, media containing S. commune-derived glucan was added to the insert and bottom for 24 h. Cells on the upper side of the insert were removed using a cotton swab, and cells on the lower side of the insert were fixed with 4% formaldehyde. After washing with PBS two times, invaded cells were stained with a 1% crystal violet solution and photographs were acquired at a 4× magnification using an Olympus IX70 microscope. Invasive ability was measured using the ImageJ program.
Western blot
HaCaT or fibroblast cells were seeded at a density of 1 × 105 each on 100 mm cell culture dishes and incubated for 24 h. These were treated with 20 μg/ml of S. commune-derived glucan in a time-dependent manner. Total protein was extracted from treated cells using RIPA buffer, and the protein concentration was determined using the BCA Protein Assay Kit. Equal amounts of protein (~ 20 μg/lane) were analyzed from each sample by resolving with 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). Protein loaded gels were transferred to NC membranes, and these were blocked in 5% skimmed milk with Tris-buffered saline Tween 20 (TBST) buffer, followed by overnight incubation with primary antibodies at 4 °C. Cytokeratin 10 (sc-53252), Cytokeratin 14 (sc-17104), FAK (sc-1688), p-FAK (sc-11765), C-Src (sc-130124), Integrin α3, α5, α6, αv, β1, β5, and GAPDH (sc-25778) antibodies were purchased from Santa Cruz Biotechnology (USA);S100A4 (ab27957) antibody was purchased from ABcam (UK); Slug (#9585), E-cadherin (#3195), and p-Src (#2101) antibodies were purchased from Cell Signaling Technology (USA). NOX4 (NB110-58849) and MMP-1 (#444209) antibodies were purchased from Novus Biologicals and Calbiochem, respectively. We used HRP-conjugated anti-mouse (K0211589, KOMABIOTECH), anti-rabbit (K021178, KOMABIOTECH), and anti-goat (AP1079P, Millipore) as secondary antibodies. Proteins were detected using chemiluminescent reagent, ECL solution (W6002, Biosesang).
In vivo experiments
Six-week old male ICR mice (n = 6 for each group) were chosen for the experiment. All procedures were approved by the Animal Care and Use Committee at Jeju National University (permission number 2015-0033). First, the fur was removed with an electronic hair clipper and removal cream. Then, wounds were made on the middle of the back using a 6-mm biopsy punch. Wounds of mice in the experimental group were treated daily with 200 μl of beta-glucans (200 μg/ml) dissolved in butylene glycol for 2 weeks. Madecassol® was used as a positive control and applied to wound sites in the same manner as beta-glucans. Images were captured at 0, 3, 7, 14 days after wounding to evaluate wound closure and the rate was calculated as a relative percentage of the original wound area, using the ImageJ program. At 7 days post-wound, skin tissue was isolated and fixed using a 4% formaldehyde solution. Paraffin-embedded tissues were cut into 4-mm sections and stained using Hematoxylin–Eosin.
Statistical analysis
GraphPad Prism 6 software was used for statistical analysis. Data are expressed as mean value with standard errors from three independent experiments. Significance between more than two groups was analyzed using one-way ANOVA, and the Student t test was assessed to compare two groups. A P value < 0.05 considered statistically significant.
Results
Four types of beta-glucan promote epithelial cell migration
First, we prepared four kinds of beta-glucan solutions (pH 7.0) (Fig. 1). To analyze whether four types of beta-glucan, derived from S. commune, black yeast, barley, and euglena, affect the skin cell viability and migration, and have different properties, we treated HaCaT keratinocytes or dermal fibroblasts with various beta-glucan concentration (0–40 μg/ml). We found that all the beta-glucans display no toxicity for HaCaT and fibroblast cells using an MTT assay (Fig. 2a, c). But, migrations of HaCaT keratinocyte were increased upon stimulation with beta-glucans and peaked at 20 μg/ml for all the tested beta-glucans (Fig. 2b). HaCaT cell lysates were treated with 20 μg/ml of beta-glucans to investigate the molecular mechanism of beta-glucan action. It was found that phosphorylation of FAK (Y397) and Slug expression sere significantly increased by S. commune-derived-beta-glucan (Fig. 2d). As the FAK signaling pathway and transition to mesenchymal cell are major factors of wound healing, these data indicate that the four types of beta-glucan have positive effects on wound repair. Especially, S. commune, beta-glucan is more effective than others.

The comparision of four types of beta-glucans in structure and effect on skin cells. a Viabilty of HaCaT and c fibroblast treated with beta-glucans derived from S. commune, black yeast, barely, and euglena. The cells (2 × 104 cells/ml) were seeded and treated with 0–40 μg/ml beta-glucan for 24 h. Viability of cells was measured using the MTT solution (5 mg/ml). b Scratch wound healing assay with HaCaT treated as described in a after 24 h. HaCaT cells (7 × 104 cells/well) were seeded on 48 well plate and a scratch was made on the cell monolayer by drawing a pipette tip across the well, and then treated with 0–40 μg/ml. Migration distance was measured using the ImageJ program. *P < 0.05 as compared to the control. d Western blotting for HaCaT cells treated with four types of beta-glucans. Cell lyastes were isolated using RIPA solution and protein concentration was measured by BCA assay. Phospho-FAK, E-cadherin, and slug expression were measured and their levels were normalized to GAPDH. The values indicate intensities of protein expression with respect to that of the loading control
Beta-glucan derived from the mushroom (S. commune) is most effective on epithelial cell motility
Based on the previous results, we selected the S. commune beta-glucan for further treatments. It displayed no cytotoxicity for the HaCaT and fibroblast cells (Fig. 3a, d), but migration of HaCaT cells was significantly increased at 20 μg/ml treatments (Fig. 3b). Invasion ability of HaCaT cells was enhanced by beta-glucan (S. commune) and peaked at 20 μg/ml (Fig. 3c). Both the migration or invasion assays are important methods to detect whether beta-glucans have an effect on wound repair. However, it is known that these assays have different actions. Invasion assay includes protrusion formation and matrix degradation and it is analyzed by staining the cells penetrating through pores. Whereas, migration assay is the way to measure the distance over which the cells migrated. Therefore, we used 20 μg/ml of the S. commune beta-glucan for further experiments. Interestingly, the migration was decreased at 80 μg/ml and rebounded again at 200–400 μg/ml. Because the beta-glucan sample contains lots of factors that occur various biological reactions and we are not yet analyze and characterize the components of beta-glucan, we guess it came from complicated reaction (Fig. 3b). More research is needed.

The stimulation effect of beta-glucan derived from S. commune on keratinocyte migration. a Viability of HaCaT and d fibroblast treated with beta-glucans derived from S. commune (0–400 μg/ml). The cells (2 × 104 cells/ml) were seeded and treated with 0–40 μg/ml for 24 h. Viability of cells was measured using the MTT solution (5 mg/ml). b Scratch wound healing assay with HaCaT treated as described in a after 24 h. HaCaT cells (7 × 104 cells/well) were seeded on a 48 well plate and a scratch was made on the cell monolayer by drawing a pipette tip across the well. Migration distance was measured using the ImageJ program. *P < 0.05 as compared to the control. c Transwell invasion assay with HaCaT cells treated with beta-glucans derived from S. commune. Invasive ability was measured by fixing with 4% of formaldehyde and staining with a 1% crystal violet solution. The photographs were acquired at × 4 magnification using a microscope. *P < 0.05 as compared to the control
Mechanisms of action of the S. commune beta-glucan on epithelial and dermal fibroblast cells
To investigate the migration mechanism of HaCaT cell treated with beta-glucan, we performed time-dependent treatments. Integrin is well known as a cell surface adhesion receptor, which acts upstream of the FAK/Src pathway. It has 24 subtypes which are formed by different combinations of one of the 18 α subunits and one of the 8 β subunits, which are non-covalently assembled together to form heterodimer [10]. Integrin α3β1 mediates keratinocyte polarization during re-epithelialization and it is also known to control TGF-β related responses [11]. Integrin α5β1 promotes keratinocyte migration [12] and αvβ1 integrins function to regulate keratinocyte adhesion during epithelial regeneration [13]. We analyzed expression of integrin subunits in the beta-glucan treated HaCaT keratinocyte and found that integrin α3β1 and α5β1 were induced by the S. commune beta-glucan. However, the expressions of integrin α6 and αv were not affected by beta-glucan stimulation (Fig. 4a).

The action mechanisms of beta-glucan (S. commune) on keratinocyte and fibroblat. a–c Western blotting for HaCaT cells treated with the S. commune beta-glucan in a time-dependent manner. Cell lysates were isolated using RIPA solution and protein concentration was measured by BCA assay. The expression levels were normalized to GAPDH. The values indicate intensities of protein expression with respect to that of the loading control. d, e Western blotting for HaCaT cells treated with the S. commune beta-glucan in a time-dependent manner. Cell lysates were isolated using RIPA solution and protein concentration was measured by BCA assay. The expression levels were normalized to GAPDH. The values indicate intensities of protein expression with respect to that of the loading control
Focal adhesion kinase (FAK) and steroid receptor coactivator (c-Src) mediate cytoskeletal dynamics and cell motility by controlling actin polymerization [9]. Phosphorylated FAK at tyrosine 397 (Y397) recruit Src and activates it byphosphorylation at tyrosine 416 (Y416) [14]. Upon the beta-glucan treatment, phosphorylation of FAK/Src was increased until 24 h expression of cytokeratin 14 was also peaked at 6 h and then decreased. Following the cytokeratin 14 down-regulation at 6 h, the intensity of cytokeratin 10 was significantly increased (Fig. 4b). Notably, the stratification in epidermis is a crucial factor of regeneration. When keratinocyte in the basal layer fully grow, they begin to differentiate rather than proliferation to spinouse, granular, and stratum layers [15]. The expression of cytoketain 10 indicates differentiation and cytokeratin 14 is used for marking proliferation of the epithelium.
Besides Integrin/FAK/Src, epithelial-mesenchymal transition (EMT) or matrix metalloproteinase (MMP) are known to be associated with cell migration [16]. However, there are no detectable changes in these pathways such as down-regulation of E-cadherin and up-regulation of Slug or MMP-1 (Fig. 4c). These data indicate the improvement of keratinocyte stimulated by the S. commune beta-glucan through Integrin α3β1, α5β1/FAK/Src activation.
ROS mainly function in a normal wound healing response, especially in dermal fibroblasts. Furthermore, NADPH oxidase (NOX) family is an important source of ROS generation [17]. For example, Sampson et al. demonstrated the ROS signaling by NOX4, which induces fibroblast transformation of myofibroblast by increasing phosphorylation of JNK. NOX4 coordinated cytoskeletal remodeling and differentiation [18]. The S. commune beta-glucan mostly promoted NOX4 synthesis at 24 h treatment (Fig. 4d).
Integrins are also required for fibroblast infiltration into wound site. Integrin αvβ5, one of the integrin subunits, participates in fibroblast transformation to myofibroblast, which is marked with FSP-1, leading to dermal contraction. Upon a time-dependent treatment of fibroblasts with the S. commune, the expression of integrin αvβ5 was increased and peaked at 48 h (Fig. 4e). As a result, the S. commune beta-glucan accelerated dermal differentiation via integrin αvβ5 and ROS production.
Wound healing effect of beta-glucans in the in vivo excisional wound model
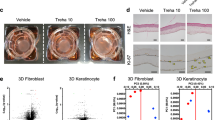
The fast wound closure occurs by epithelial regeneration following dermal remodeling. In an in vivo experiment, we made a wound using biopsy punch and measured the margin area at different time points. At day 3 after wounding, there is no significant difference between groups. However, the wound closure of S. commune-treated-group was accelerated in proliferative phase (Fig. 5a, b). Scab, formed during hemostasis, is a fibrin clot. As epithelial cell grows and migrates, scab loss occurs and it was used to compare the healing capacity [19]. Except for the control and black yeast-treated groups, scab began to fall apart from wound at day 8 after wounding. Scab loss in the S. commune-treated-group was twice as much compared to other groups (Fig. 5c). After completion of wound healing, we stained using hematoxylin and eosin using isolated skin tissue. As shown in Fig. 5d, re-epithelialization is in progress under scars at day 7. The S. commune-treated-group showed elongated and thinly-formed epithelium compared to other groups. These results indicate that beta-glucan derived from S. commune has a beneficial wound healing effect.

The wound healing effect of beta-glucans in an in vivo excisional model. a, b Wound closure in an excisional wound model for 14 days. Photos were taken every 3 days and wound area was measured using the ImageJ program. ‘Madecassol’ was used as a positive control. c The number of scab loss at day 8 after wounding. The number of scab uncombined mouse were counted and quantifiably shown as a graph (n = 6 each group). d Hematoxylin and eosin staining of skin tissue isolated from the wound site on day 7. The photographs were acquired at ×100 magnification using a microscope. Re-epithelialization was measured using the ImageJ program and showed as a graph. Dotted line; a boundary line of wound site, GT granulation tissue. *P < 0.05 as compared to the control
Discussion
Our study demonstrates that beta-glucan from S. commune accelerates cutaneous wound healing. Beta-glucans have a common backbone, β-1,3-glucan, which is a key factor in determining their function. The length and number of branched chains is also important for specific binding to cells of the immune system [20]. Beta-glucans are also known to promote resistance to viral, bacterial, and fungal diseases, as well as exhibit antitumor effects. In particular, beta-glucans derived from mushrooms have multiple short branches. Additionally, it is known that mushroom extract contains other effective ingredients of potential nutritional value (Fig. 1) [21]. However, the wound healing effect of beta-glucans and their roles in repairing skin injury have not been comprehensively studied.
Because of their wound healing properties, beta-glucan have been used in the development of bioartificial skin, considering their ability to bind to gelatin and collagen. It is known that wound dressing with beta-glucan enhance wound healing and reduce pain. Furthermore, beta-glucans increase proliferation, produce diverse cytokines, and release ROS in dermal fibroblast cells, leading to neutrophil infiltration and angiogenesis [22]. Here, we found that S. commune beta-glucan promotes dermal contraction by inducing the expression of NOX4 (which contributes to ROS generation) and Integrin αvβ5 (Fig. 4).
There are four different pattern recognition receptors (PRRs) for beta-glucans in immune system cells, complement receptor 3 (CR3), scavenger receptor, lactosylceramide, and dectin-1 [23]. The dectin-1 beta-glucan receptor, has two isoforms the full-length isoform A and the stalk region-lacking isoform B. A previous study by Linda et al., showed that dectin-1 isoform B is expressed in HaCaT keratinocytes and possesses functions which are different from the ones observed in the immune system cells. These functions include promoting proliferation and migration, which are major processes during re-epithelialization [22]. Spleen tyrosine kinase (Syk) is a mediator of dectin-1 activation and is phosphorylated by beta-glucans [24]. Syk is phosphorylated by active Src, thereby induces cell spreading. On the contrary, active Syk promotes Integrin αV translocation to the membrane of epithelial cells and control migration through FAK and Src [25].
As shown by these reports, several receptors can recognize beta-glucans epithelial cells, with beneficial effects for wound repair. We found that S. commune beta-glucan induces FAK/Src pathway activation via integrin up-regulation in HaCaT keratinocyte cells. In addition to the Integrin/FAK/Src pahtway, there are several indications of other action mechanisms by which S. commune beta-glucan may promote wound healing. One possibility is that dectin-1 and integrins stimulated by beta-glucans phosphorylate Src or Syk, forming a positive loop between them that can promotes cell migration. Further experiments are needed to prove this hypothesis.
We demonstrated the cutaneous wound healing effects of four types of beta-glucans, and found that S. commune beta-glucan is especially efficacious in wound repair due to its ability to promote re-epithelialization and dermal contraction by activation of Integrin/FAK/Src in the epidermis as well as by dermal transformation via NOX4 expression and integrin up-regulation. These findings suggest that beta-glucan from S. commune is a potential therapeutic agent for skin injury.
References
Lattimer JM, Haub MD (2010) Effects of dietary fiber and its components on metabolic health. Nutrients 2:1266–1289
Yun CH, Estrada A, Van Kessel A, Park BC, Laarveld B (2003) Beta-glucan, extracted from oat, enhances disease resistance against bacterial and parasitic infections. FEMS Immunol Med Microbiol 35:67–75
Volman JJ, Ramakers JD, Plat J (2008) Dietary modulation of immune function by beta-glucans. Physiol Behav 94:276–284
Lee SH, Jang GY, Hwang IG, Kim HY, Woo KS, Kim KJ, Lee MJ, Kim TJ, Lee J, Jeong HS (2015) Physicochemical properties of beta-glucan from acid hydrolyzed barley. Prev Nutr Food Sci 20:110–118
Rahar S, Swami G, Nagpal N, Nagpal MA, Singh GS (2011) Preparation, characterization, and biological properties of beta-glucans. J Adv Pharm Technol Res 2:94–103
Zhao XK, Cheng Y, Liang Cheng M, Yu L, Mu M, Li H, Liu Y, Zhang B, Yao Y, Guo H et al (2016) Focal adhesion kinase regulates fibroblast migration via integrin beta-1 and plays a central role in fibrosis. Sci Rep 6:19276
Darby IA, Laverdet B, Bonte F, Desmouliere A (2014) Fibroblasts and myofibroblasts in wound healing. Clin Cosmet Investig Dermatol 7:301–311
Kim YM, Huh J, Lim Y, Cho M (2015) Soy isoflavone glycitin (4′-hydroxy-6-methoxyisoflavone-7-d-glucoside) promotes human dermal fibroblast cell proliferation and migration via TGF-β signaling. Phytother Res 29:757–769
Sophors P, Kim YM, Seo GY, Huh JS, Lim Y, Koh DS, Cho M (2016) A synthetic isoflavone, DCMF, promotes human keratinocyte migration by activating Src/FAK signaling pathway. Biochem Biophys Res Commun 472:332–338
Lambert AW, Ozturk S, Thiagalingam S (2012) Integrin signaling in mammary epithelial cells and breast cancer. ISRN Oncol 2012:493283
Reynolds LE, Conti FJ, Silva R, Robinson SD, Iyer V, Rudling R, Cross B, Nye E, Hart IR, Dipersio CM, Hodivala-Dike KM (2008) Alpha3beta1 integrin-controlled Smad7 regulates reepithelialization during wound healing in mice. J Clin invest 118:965–974
Chen WW, Tjin MS, Chua AWC, Lee ST, Tay CY, Fong E (2017) Probing the role of integrins in keratinocyte migration using bioengineered extracellular matrix mimics. ACS Appl Mater Interfaces 9:36483–36492
Koivisto L, Heino J, Hakkinen L, Larjava H (2014) Integrins in wound healing. Adv Wound Care (New Rochelle) 3:762–783
Baillat G, Siret C, Delamarre E, Luis J (2008) Early adhesion induces interaction of FAK and Fyn in lipid domains and activates raft-dependent Akt signaling in SW480 colon cancer cells. Biochim Biophys Acta 1783:2323–2331
Fuchs E (1994) Epidermal differentiation and keratin gene expression. Princess Takamatsu Symp 24:290–302
Radisky ES, Radisky DC (2010) Matrix metalloproteinase-induced epithelial-mesenchymal transition in breast cancer. J Mammary Gland Biol Neoplasia 15:201–212
Seo GY, Lim Y, Koh D, Huh JS, Hyun C, Kim YM, Cho M (2017) TMF and glycitin act synergistically on keratinocytes and fibroblasts to promote wound healing and anti-scarring activity. Exp Mol Med 49:e302
Sampson N, Koziel R, Zenzmaier C, Bubendorf L, Plas E, Jansen-Durr P, Berger P (2011) ROS signaling by NOX4 drives fibroblast-to-myofibroblast differentiation in the diseased prostatic stroma. Mol Endocrinol 25:503–515
Eming SA, Martin P, Tomic-Canic M (2014) Wound repair and regeneration: mechanisms, signaling, and translation. Sci Transl Med 6:265–266
Chan GC, Chan WK, Sze DM (2009) The effects of beta-glucan on human immune and cancer cells. J Hematol Oncol 2:25
Friedman M (2016) Mushroom polysaccharides: chemistry and antiobesity, antidiabetes, anticancer, and antibiotic properties in cells, rodents, and humans. Foods. https://doi.org/10.3390/foods5040080
Murphy EA, Davis JM, Carmichael MD (2010) Immune modulating effects of beta-glucan. Curr Opin Clin Nutr Metab Care 13:656–661
van den Berg LM, Zijlstra-Willems EM, Richters CD, Ulrich MM, Geijtenbeek TB (2014) Dectin-1 activation induces proliferation and migration of human keratinocytes enhancing wound re-epithelialization. Cell Immunol 289:49–54
Cohen-Kedar S, Baram L, Elad H, Brazowski E, Guzner-Gur H, Dotan I (2014) Human intestinal epithelial cells respond to beta-glucans via Dectin-1 and Syk. Eur J Immunol 44:3729–3740
Fotheringham JA, Coalson NE, Raab-Traub N (2012) Epstein-Barr virus latent membrane protein-2A induces ITAM/Syk- and Akt-dependent epithelial migration through alphaV-integrin membrane translocation. J Virol 86:10308–11032
Authors’ contributions
GS did most of experiments except animal tissue histology. CH did histology. SC prepare and supply beta-glucan. YMK and MC plan and supervise whole experiments and edit and correct writing. All authors read and approved the final manuscript.
Acknowledgements
This work was supported by the Korea Industrial Complex Corporation through the Project Fostering the Industrial Cluster (No. RKK17008).
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Seo, G., Hyun, C., Choi, S. et al. The wound healing effect of four types of beta-glucan. Appl Biol Chem 62, 20 (2019). https://doi.org/10.1186/s13765-019-0428-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13765-019-0428-2