
Abstract
Background
Elevated atmospheric CO2 has direct and indirect influences on ecosystem processes. The impact of elevated atmospheric CO2 concentration on carbon and nitrogen transformations, together with the microbial community, was evaluated with water hyacinth (Eichhornia crassipes) in an open-top chamber replicated wetland. The responses of nitrogen and carbon pools in water and wetland soil, and microbial community abundance were studied under ambient CO2 and elevated CO2 (ambient + 200 μL L−1).
Results
Total biomass for the whole plant under elevated CO2 increased by an average of 8% (p = 0.022). Wetlands, with water hyacinth, showed a significant increase in total carbon and total organic carbon in water by 7% (p = 0.001) and 21% (p = 0.001), respectively, under elevated CO2 compared to that of ambient CO2. Increase in dissolved carbon in water correlates with the presence of wetland plants since the water hyacinth can directly exchange CO2 from the atmosphere to water by the upper epidermis of leaves. Also, the enrichment CO2 showed an increase in total carbon and total organic carbon concentration in wetland soil by 3% (p = 0.344) and 6% (p = 0.008), respectively. The total nitrogen content in water increased by 26% (p = 0.0001), while total nitrogen in wetland soil pool under CO2 enrichment decreased by 9% (p = 0.011) due to increased soil microbial community abundance, extracted by phospholipid fatty acids, which was 25% larger in amount than that of the ambient treatment.
Conclusion
The study revealed that the elevated CO2 would affect the carbon and nitrogen transformations in wetland plant, water, and soil pool and increase soil microbial community abundance.
Similar content being viewed by others


Introduction
Carbon dioxide (CO2) is the most important greenhouse gas, which has increased by around 2 ppm of the current rate per year. Enriched atmospheric CO2 concentration can both enhance and improve soil microorganism activities through plant photosynthesis, organic substrate quantity, organism’s respiration, and microbial biomass allocation in the ecosystem (Liu et al. 2018). There are many considerable interests in understanding whether the elevated atmospheric CO2 can regulate the carbon sequestration and nitrogen mineralization in soil, and how these changes respond to the ecological environment (Yu and Chen 2019).
Increase in elevated atmospheric CO2 causes an increase in nitrogen immobilization (Chang 2019) and a reduction in nitrification in soils by increasing the allocation of carbon to roots. The increasing CO2 is generally associated with the reduction in nitrate reductase activity, which might reduce the availability of electron acceptors for denitrification (Chang 2019). The rising atmospheric CO2 has direct and indirect influences on nitrogen losses in ecosystems. A study by Conthe and coworkers (2019) revealed that the direct and indirect ways are releasing NO and N2O into the atmosphere and leaching for NO3−-N and denitrification, respectively. Other studies revealed that increasing CO2 (inflow of C to the soil) would change the rates of nitrogen mineralization (Gao et al. 2019). The rate of N mineralization is crucial in the determination of available nutrients for vegetative production (Pang et al. 2019). On the other hand, the status of available nutrients may also indicate N losses or gains from terrestrial ecosystems (Yu and Chen 2019). Alternatively, plants grown under elevated CO2 showed variations in root responses, depending on species and environmental conditions such as temperature, pH, light, water, and nutrient availability (Gao et al. 2019). Furthermore, Zhang et al. (2019a) showed significant effects of soil pH on soil respiration with microbial activity. Thus, it is important to investigate the effects of elevated CO2 on the environment.
The increasing atmospheric CO2 concentrations have the potential alteration of carbon cycling for terrestrial ecosystems (Yu and Chen 2019). Previous works have demonstrated that a rise in atmospheric CO2 can cause a greater exudation of labile C from plant roots. This promotes the growth of microbes (Pang et al. 2019). A report showed that changes in C availability (organic matter) or substrate quality (lower soil C to N ratio), would promote the growth of Gram-negative bacteria (Laut et al. 2020). High plant growth under elevated CO2 is likely to provide more organic substrates for microbial metabolism in soil, and fuel more rapid rates of soil carbon cycling (Yang et al. 2019). Bacteria in the soil can recycle carbon (C), nitrogen (N), sulfur (S), and other chemical elements in the decomposition process. Bacteria contribute to nutrient mineralization in soil (Zhang et al. 2019a). Accordingly, the soil microbial biomass is a more sensitive indicator of soil fertility than the soil organic matter because it responds readily to change the soil’s physical and chemical environment (Brym et al. 2014; Laut et al. 2020). It must be noted that elevated CO2 could lead to an increase in microbial community growth and immobilization of N in soil (Yang et al. 2019; Xing et al. 2020).
In recent years, the phospholipid fatty acids (PLFAs) analysis method has been widely employed and recognized to be a robust technique to study soil microbial community (Fan et al. 2017; Li et al. 2018). In this study, this method was used to measure the composition and abundance of the microbial community in wetland soil under two atmospheric CO2 levels.
Previous studies reported mainly on induced CO2 response on the plant nutrient cycling and soil microbial community (Fan et al. 2017). Elevated atmospheric CO2 concentration adversely affects global warming and climate and inversely affects the plant growth responses to photosynthesis, soil carbon sequestrations, and nitrogen cycling. Understanding the effect of elevated CO2 on various wetland systems is paramount. In fact, plants and microbes directly respond to the ecosystems with the increases of elevated atmospheric CO2. The objective of this present study is to examine the interconnected processes in wetland ecosystem for correlation of carbon, nitrogen cycle with water hyacinth (Eichhornia crassipes) in water, wetland soil, and soil microbial community composition between two concentrations of atmospheric CO2.
Also, the field observation of CO2 effect on specific C and N cycling in a natural lake ecosystem is difficult to analyze. This present research contributes to understanding the internal problem of the natural lake ecosystems between two atmospheric CO2 (elevated and ambient) concentrations. Given the research needs, open-top chamber (OTC) for CO2 elevation was used as the imitation of field observation of natural lake ecosystems, in which sediment and wetland plant species were used. Therefore, the carbon (C) and nitrogen (N) transformations, total organic carbon (TOC), inorganic carbon (IC), total carbon (TC), total nitrogen (TN), and nitrogen interactions in water and wetland soil were monitored and the microbial community abundance was explored using the method of PLFAs in this experiment.
Materials and methods
Experimental design and treatments
Open-top chamber (OTC)
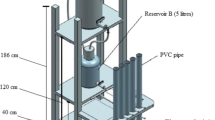
A sketch of an open-top chamber (OTC) used in this study is shown in Fig. 1. OTC was 13.5 m3 in volume and the base was made by a brick frame and it was covered by 0.15-cm-thick glass and placed inside a ventilated greenhouse. One OTC was used for ambient CO2 and another was used for elevated CO2 (ambient + 200 μL L−1) without any input of N treatment. Each OTC was separated into four partitions. Among these four partitions, two of them were cultured with water hyacinth and the remaining two were used as control (without plants and soil). The greenhouse was equipped with a fan that continually blew air into the chamber to adjust the daily room temperature (24–35 °C). Elevated CO2 was pumped into the chamber through a plastic drainage pipe (diameter 0.5 cm), which was laid inside of open-top chamber along the edges. To control CO2 concentration, TES 1370 NDIR CO2 meter was used daily to detect the concentration of CO2 inside two chambers.

The sketch of the open-top chamber indicating the elevating CO2 into the open-top chamber
Experimental site, sediment, and plant sampling
Sediments used in this experiment were collected from Taihu lake, an area of approximately 36895 km2 (31° 25.106′ N, 120° 14.758′ E), in the east of China. An equal height, about 14–15 cm (600 kg in weight), of air-dried sediments was placed into duplicated partitions before loading the culture solution into the open-top chamber.
Water hyacinth was obtained from the lake on the campus of Jilin Agricultural University, Changchun, China. After collection, the plants were washed and air-dried in a shady and cool place. Uniform leaves and healthy plants that weighed approximately 200 g were cultured inside the duplicated partitions appropriately. The cultured solution (tap water being used) of 40 cm in depth, which was nearly equal to 1 l, was poured into the chambers.
Chemical analysis for water and soil
The experiment was run for 120 days. The soil samples were dried naturally and grounded. The grounded sample was sieved through a 2-mm sieve. Soil pH was measured in a ratio of 1:2 representing a mixture of soil to deionized water. A measurement of TN for soil was conducted using a UV spectrometry (SHIMADZU UV-2450) as soon as the samples were pre-filtered with a 0.45 μm glass fiber filter. Water samples were collected and pre-treated by the SHIMADZU TOC-CPN. Water TOC, TC, IC, and TN were determined by using TOC/TN analyzer (SHIMADZU TOC-CPN) and soil TOC, TC, and IC were analyzed by the SSM analyzer (SHIMADZU TOC-CPN), respectively. Gas chromatography-mass spectrometry (GC-MS) QP-2010 Plus with AOC-20i verified the phospholipid fatty acids (PLFAs).
Methodology for GC-MS
Fatty acid methyl esters (FAMEs) were analyzed by GC-MS (QP2010 Plus SHIMADZU), with VF-23 ms fused silica (30 m × 0.25 mm × 0.25 μm) (Varian, Inc.). The methodology for GC-2010 was a 1-μl single injection, − 1 split ratio, split-less mode, with gas Helium as carrier together with pressure 57.4 kPa, and the oven temperature: (1) from 50 (held for 2 min) to 100 °C, at 5 °C/min; (2) from 100 to 200 °C, at 4 °C/min; and (3) from 220 °C (held for 4 min) to 240 °C, at 4 °C/min. The column and temperature program details were ion source temperature of 200 °C, interface temperature of 260 °C, and solvent cut time of 5 min, respectively.
Microbial phospholipid fatty acids (PLFAs) profile
The extraction of lipids from fresh sediment in fractionation and quantification was described by Guo et al. (2019). After 5 g of fresh sediment was randomly picked out, visible fine roots were firstly removed and gravimetric water content was then measured at 105 °C, the samples were moved to the glass tube (Teflon-lined screw cap). The samples were fractionated using silicic acid chromatography. Also, a stream of nitrogen gas was used to dry the phospholipid fraction and later stored at − 20 °C. The resultant polar lipid fractions were then subjected to mild alkaline methanolysis, and the resultant fatty acid methyl esters (FAMEs) were extracted with two 2-ml aliquots of chloroform. The sample was dried under nitrogen gas at room temperature. The resultant FAMEs were re-liquefied in 1 ml hexane containing an internal standard (19:0 FAME at 40 ng/ml) to transfer to GC vials of (GC-MS) for the analysis of microbial community abundance.
The nomenclature for microbial PLFAs
Based on Frostegård and Bååth (1996), the designation for the systematic names of FAMEs was A:BωC, where
-
A—the total number of carbon atoms
-
B—the number of double bonds
-
ω—position of the first double bond
-
C—the number of carbon atoms from the aliphatic end
Suffixes “c” and “t” refer to the “cis” and “trans” conformations, respectively.
Methyl branching, at the “iso and anteiso” positions from the methyl end of the molecule, and methyl branching at the 10th carbon atom from the carboxyl end of the molecule are designated by the prefixes “i”, “a”, and “10Me” respectively. The prefix “cy” denotes cyclopropane fatty acids.
Plant relative CO2 treatment effect
The relative CO2 treatment effects (%) were calculated by Eq. 1:
The floating aquatic plant (water hyacinth) was harvested at the end of the growing period, i.e., (treatment for 4 months) to measure the whole-plant biomass under two different CO2 concentrations.
Statistical and data analyses
Treatment effects (the CO2 effect) on data analyses were analyzed using one-way ANOVAs and CO2 effects were also analyzed with paired-samples t test with using SPSS Statistics 17.0 (SPSS Inc., USA), respectively. The relation of samples (R2) was assessed statistically by the linear regression analysis of SPSS Statistics 17.0. All determinations of chemical analysis for plant, water and soil samples were performed in triplicate analyses with one blank from duplicate treatments in OTCs. The measurement of plant growth was expressed as mean of duplicate treatments in OTCs. The data are presented as mean ± standard error, p, and F value. Statistical significance was accepted at α = 0.05.
Results
Effects of elevated CO2 on plant biomass
After four months of incubation, whole-plant biomass showed an 8% increase (p = 0.022) under elevated CO2 compred to ambient CO2 (Fig. 2). Water TC with plant and water TC as control (without plant) changed (in time) for elevated chamber and ambient chamber during treatment CO2 (Fig. 3). The presence of plants influenced the CO2 effects in the experiment. From the study (Fig. 3), the mean water TC with the plants was increased by 21% in time with 10.52 ± 0.3 mg/L with plants as compared to 8.69 ± 0.2 mg/L without plants (control) under the same CO2 concentration. However, there were no significant differences (p > 0.05) for water TC as control (1%) with 8.69 ± 0.2 mg/L in elevated treatment as compared to 8.57 ± 0.3 mg/L in ambient treatment.

Effects of elevated CO2 concentration on whole-plant biomass accumulation. The average percent showed the CO2 effect. Mean plant biomass (n = 2 times observation with duplicates) affected by enrichment CO2 and S.E. located on the length of value

a Water TC (with plants) under elevated treatment, b water TC (without plants) under elevated treatment, c water TC (with plants) under ambient treatment, and d water TC (without plants) under ambient treatment (in time) during treatment. Each value indicated mean and standard error (S.E.). There were no significant differences between the two treatments (p ≥ 0.05)
Carbon content in water and soil with plants
There was an increase of 7% (p = 0.001, F = 34.229) for mean water TC with the plants with 10.52 ± 0.3 mg/L under CO2 treatment and 9.83 ± 0.2 mg/L under ambient treatment (Fig. 4a). There was a significant increase of 21% (p = 0.000, F = 137.587) for water TOC with 5.56 ± 0.3 mg/L in the elevated treatment as compared to the 4.58 ± 1.0 mg/L in the ambient treatment (Fig. 4b). But water IC concentration was lower in elevated CO2 treatment than ambient treatment. There was a decrease in IC concentration with time by 5% (p = 0.540, F = 0.422). The concentration was 4.96 ± 0.7 mg/L in the elevated treatment as compared to 5.24 ± 0.5 mg/L in the ambient treatment correspondingly (Fig. 4c).

a–c Mean concentration (mg/L) of water TOC, TC, and IC. The average percent showed the CO2 effect. The length of each value was mean (mean of three replicate) indicated in elevated CO2 (black) and ambient CO2 (white) and their standard error (S.E.) (n = 8 times observation with duplicates). Statistical significance, α = 0.05
In this experiment, there is a significant difference in the soil carbon changes between the two treatments. In the high CO2 treatment, soil TC concentration was 3% (p = 0.344, F = 1.057) higher relative to the ambient treatment (Fig. 5a), and soil TOC concentration was 6% (p = 0.008, F = 2.573) greater as indicated in Fig. 5b. The mean content of TC, under elevated CO2, was 13.42 ± 0.34 mg/g against 12.91 ± 0.52 mg/g under ambient CO2. Similarly, the mean content of TOC, under elevated CO2, was 10.74 ± 1.21 mg/g against 10.12 ± 0.89 mg/g under ambient CO2. Soil IC declined by 3% (p = 0.257, F = 1.570) (Fig. 5c). The mean content of IC, under elevated CO2, was 2.68 ± 0.23 mg/g against 2.79 ± 0.25 mg/g under ambient CO2.

a–c Mean concentration (mg/L) of soil TOC, TC, and IC for two treatments. The average percent showed the CO2 effect. The length of each value was mean (mean of three replicate) indicated in elevated CO2 (black) and ambient CO2 (white) and their standard error (S.E.) (n = 8 times observation with duplicates). Statistical significance, α = 0.05
Total nitrogen in water and soil with plants
There was a substantial increase of 26% (p = 0.000, F = 245.281) in the mean water TN concentration with 1.39 ± 0.3 mg/L in the elevated treatment as compared to 1.10 ± 0.07 mg/L in the ambient treatment at the end of the experiment (Fig. 6a). The mean N concentration for wetland soil under CO2 enrichment was 9% decrease (p = 0.011, F = 13.405), which was lower than in ambient treatment with 0.55 ± 0.36 mg/g dried soil for higher CO2 treatment and 0.60 ± 0.07 mg/g dried soil for ambient treatment from initial to final extraction time, respectively (Fig. 6b). The total N loss of the water and soil pool under elevated chamber was 9% with 110 ± 3.3 mg/g dried soil CO2 under elevated chamber paralleled to 121 ± 4.7 mg/g dried soil CO2 under ambient chamber (Fig. 7). This result was consistently maintained throughout the experimental process.

a, b Mean concentration of water TN and soil TN for two treatments during the experimental periods. The average percent showed the CO2 effect. The length of each value was mean (n = 8 times observation with duplicates) indicated in elevated CO2 (black) and ambient CO2 (white) and standard error (S.E.) located on the length of values

Average total N content was lost by CO2 enrichment between two treatments from beginning to end of the experiment. The average percent showed different TN content between the two chambers. The length of each value was mean indicated in elevated CO2 (black) and ambient CO2 (white) and standard error (S.E.) located on the length of values
Soil microbial community abundance
General background for taxonomic and functional biomarkers
Microbes can be distinguished into four groups: one with a high proportion of methyl branched-chain or monounsaturated FAME’s, i.e., Gram-positive bacteria; the second, with a large proportion of cyclopropane fatty acids, i.e., Gram-negative bacteria; the third, with fungal indicators of PLFAs: C18:1ω9t (saprotrophic fungi), C18:3ω3c and C18:2ω6c and C16:1ω5c (mycorrhizal (AM) fungi); and lastly, C20:4ω6c was a species of protozoa as shown in Table 1 (Zhang et al. 2019b; Tzanakakis et al. 2019; Gorman-Lewis et al. 2019).
In this study, Gram-positive bacterial PLFAs or nitrifiers (ammonia-oxidizing bacteria) were iC14:0, C16:0, C17:0, C18:0, and C20:0 as indicated in Table 1. Among them, methane utilizing bacteria were C16:0 and C18:0. Gram-negative bacteria or microeukaryotes PLFAs were cyc17:0 (sulfate-reducing bacteria), 10meC16:0 (actinomycetes), C16:1ω5c, C16:1ω7c, and C18:1ω9t. Moreover, C18:3ω3c and C18:2ω6c were fungi.
Reaction of microbial PLFAs during CO2 run
Figure 8 showed each and the total content of microbial PLFAs for sediments under elevated CO2 and ambient CO2. In CO2 treatment, total microbial community abundance was 45.4 nmol/g (p = 0.023) whereas 36.2 nmol/g in ambient conditions. These microbial PLFAs showed a 25% significant difference between elevated and ambient conditions during the course of the CO2 enrichment.

Comparison of concentrations (nmol/g) of different microbial PLFAs. The length of each value indicated less and greater quantitative result was mean than ambient treatment for during enrichment CO2 (white) and S.E. There were significant differences between two treatments at (p ≤ 0.05). Values are means ± S.E
The Gram-negative and -positive bacterial PLFAs contents could be observed significantly in CO2 enrichment conditions. The total Gram-positive bacteria were 3.26% (F = 8.261, p = 0.045), Gram-negative bacteria were 136.27% (F = 22.261, p = 0.0001), total fungi were 46.23% (F = 5.855, p = 0.137), and protozoa were 21.00% than ambient conditions, respectively (Table 1).
Discussion
Effects of elevated atmospheric CO2 concentration on plant biomass and C content in soil and water
Numerous studies approved that elevated CO2 could increase the biomass of plants. In this study, we showed an increase in plant biomass of water hyacinth by 8% (p = 0.022). Song et al. (2009) suggested that increased atmospheric CO2 consistently enhanced the growth of water hyacinth. Water TC increased by 7% (p = 0.001) and water TOC increased by 21% (p = 0.000) by CO2 effect. The increasing of water TOC may depend on plant that absorbs CO2 from the atmosphere to water and convert inorganic CO2 to labile organic C, TOC in water because the free-floating aquatic plants can directly exchange CO2 with the atmosphere through the upper epidermis of leaves (Satake and Shimura 1983). Soil TC increased by 3% (p = 0.344) and soil TOC increased by 6% (p = 0.008) by CO2 effect. According to Walter and Heiman (2000), the transport pathway of C from water to sediment increased through the vascular tissue of the plant below the water tables; thus, labile organic C in soil increased with the increasing CO2. Moreover, concentrations of water and soil IC decreased by 5% (p = 0.540) and 3% (p = 0.257), respectively. The plant may have used IC for plant respiration and photosynthesis effects on its growth and many aspects of plant physiology. Another explanation may be the severely reduced supply of IC due to slower CO2 diffusion rates through underwater photosynthesis (Madsen and Sandjensen 1994).
Generally, an increased TOC concentration in water and soil was due to the plant biomass enhancement because water TC without plant did not significantly increase even under elevated CO2 concentration. CO2 did not viscously dissolve in water when the pool did not have plants under elevated CO2. Water TC concentration without plant exposed to high CO2 levels slightly increased by 1.4% (p = 0.046) compared with that exposed to ambient CO2 and it was statistically significant. Water TOC concentration with no plant under high CO2 levels decreased by 1.1% (p = 0.011) and water IC increased by 4.1% (p = 0.173), respectively. Absorption of CO2 from the atmosphere to water may depend on CO2 exchange in plants. According to Maltais-Landry et al. (2009), planted wetlands may sequester 15 times more C than unplanted wetland systems. Thus, the increase in C content in wetlands can be attributed to the increment in plant growth.
In short, wetland plants play an important role in nutrients cycling due to nitrogen uptake, storage, and release processes in the wetland ecosystem (Wang et al. 2013). Wetland plants that will be affected by future CO2 concentrations will likely follow a common pattern of carbon storage in water and soil as well. Commonly, elevated CO2 induces growth which increases wetland plants relative to the equal measurement of ambient CO2 and the following results synchronize with nitrogen dynamics and changes in microbial activity (Yu and Chen 2019; Kelly et al. 2013). The wetland ecosystem can direct the net balance between carbon gains and losses. The C storage occurred due to the combined effects of the increased plant productivity resulting from increased CO2 and drastic changes in plant residues. The comparison of water TC, with plants, to water TC as a control (without plants), in the elevated chamber, showed that the water TC with plants increased in time (21%) than that of the control. By contrast, exposure of water TC without plants to high CO2 had no significant effect as compared to ambient treatment of water TC without plants, which increased by 1%.
Effects of elevated atmospheric CO2 concentration on N cycling (soil/water) and microbial community abundance
Several research works have elucidated that effect of elevated CO2 on dissolved organic C and its relationship with soil microorganisms (Zhao et al. 2009; Jinbo et al. 2007; Hungate 1999). Soil organic C is an important labile C fraction because it is the main energy source for microorganisms to increase N mineralization rate in freshwater marshes (Zhao et al. 2009; Jinbo et al. 2007; Hungate 1999). Dissolved organic C has also been proposed as an indicator of the C available to soil microorganisms (Mikan et al. 2000; Kang et al. 2005; Smolander and Kitunen 2002; Sowerby et al. 2000). In this study, under elevated CO2, TN in water increased by 26% (p = 0.0001) compared to ambient CO2. The increase may be attributed to the different microbial populations and functions since soil microbial community could substantially transform N into gasses (called nitrous oxide and nitrogen gas) in wet soil which dissolve in water. Moreover, sediment is a net sink of N, which is the net transfer of N from the water column to sediment or from sediment to water.
Similarly, decreasing in soil TN concentrations (9%, p = 0.011) may also depend on the soil microbial community abundance because increasing C input to the soil under elevated CO2 could lead to increased microbial growth, immobilization of N, and thereby reduction of dissolved N in soil (Nord et al. 2015; Niklaus et al. 1998). Increased N demand by heterotrophic microorganisms (ammonium used bacteria) would decrease ammonium concentrations decreasing nitrate (oxidation from ammonium) availability for nitrifiers (nitrate used bacteria) and this may follow decrease in soil nitrate concentrations (Hungate 1999). The ammonia-oxidizing bacterial community in the sediment surface can impact the transport of nitrogen to overlying water (Satoh et al. 2007). Sediments are a major site of the nitrification-denitrification by main microbes that can remove the labile N from the sediments to overlying water bodies and thereby reduction in N concentration in soil (Satoh et al. 2007).
In this study, as C17:0, ammonia-oxidizing bacteria became dominant in elevated CO2 soil, its populations and functions also stimulate the N cycle (Satoh et al. 2007). Ammonia-oxidizing bacteria convert ammonia into nitrous oxide, thereby supporting higher rates of denitrification and the main removal pathway for N to water. When fungi are inhibited in soil, denitrifiers or ammonia-oxidizing bacteria produced a substantially high amount of nitrous oxide and increased nitrate availability in soil (Hu et al. 2001; Balser and Firestone 2005). Anaerobic sulfate-reducing bacteria (cycC17:0) is also associated with N2O production. C16:1ω5c, a mycorrhizal fungi biomarker, dominated elevated CO2 soil in the present study. Mycorrhizal fungi increased the community metabolic efficiency and enhanced C storage as well as increases fungal PLFAs, and C18:2ω6c which may be connected with the decreased total and available soil N, and hence, denitrification occurs (Hu et al. 2001; Balser and Firestone 2005). N2O production by denitrification is especially enhanced at high C content due to the energy source for microbes to enhance the heterotrophic nitrification process (Hu et al. 2001; Balser and Firestone 2005).
Moreover, the increased soil labile TOC at elevated CO2, when it occurs, enhances higher microorganisms resulting in decreased soil N concentrations (Barnard et al. 2005). Gram-negative bacteria, which lead to further C and N recycling, were higher in abundance (by 136.27%, p = 0.011) under elevated treatment in this study and contributed to soil C storage as well as lead to more available N in the soil (Balser and Firestone 2005; Fraterrigo et al. 2006). Lipson and Näsholm (2001) mentioned that dominant Gram-negative bacteria are associated with nitrous oxide production (denitrification). Soil C sufficiently supports denitrifier populations resulting in denitrification (Wrage et al. 2001).
The results from this study also agreed with other studies (Wrage et al. 2001; Jensen and Andersen 1992; James et al. 1997). The diffusive NH4+ transformation by ammonia used bacteria from aquatic sediments contribute a significant amount of N to the water column. Although excessive external nutrient loads have reduced, internal nutrient loads can have significant influence on the water quality of shallow lakes (Wrage et al. 2001; Jensen and Andersen 1992; James et al. 1997). Since soil N concentration was decreased (9%, p = 0.011) by microbial activity, such decreased soil N reflected the increased overlying water concentration of N (26%, p = 0.0001). Therefore, the contribution of biochemical pathways of internal cycles of C and N to water is governed by the compositions of the bacterial populations and the functions of the resident soil. The results showed that under higher CO2 this is a significant effect on the microbial community composition and cascading impact on the wetland ecosystem of plant, water, and soil of C and N cycling during the growing season.
Conclusions
The study showed conclusively that the effect of CO2 enrichment is likely to increase plant biomass, and positively affect the microbial community abundance in the soil of wetlands. Besides, C availability is one of the main driving forces to increase dissolved inorganic nitrogen in water which would closely relate to soil mineralization and microbial uptake of C by a significant increase of the microbial community abundance under elevated CO2. The concentration of TOC concentrations in water with plants increased significantly under elevated CO2 that consequently indicated the significant increase in concentrations of TOC in the soil. This suggests that the upper epidermis of leaves and roots of plants can directly exchange CO2 (IC) for labile TOC from the atmosphere to water and soil. Under elevated CO2 with plants, significant increase in the TC and TOC concentrations to soil may increases microbial-C availability thereby stimulating increased demand for N, increasing microbial NH4+ immobilization (consumption) and decreasing net NH4+ mineralization (production) in soil. The information garnered from this study complements past studies and thereby provides valuable information to stakeholders and researchers on understanding the impact of CO2 elevation on the ecosystem.
Availability of data and materials
Not applicable
Abbreviations
- CO2 :
-
Carbon dioxide
- C:
-
Carbon
- N:
-
Nitrogen
- PLFAs:
-
Phospholipid fatty acids
- OTC:
-
Open-top chamber
References
Balser TC, Firestone MK (2005) Linking microbial community composition and soil processes in a California annual grassland and mixed-conifer forest. Biogeochemistry 73(2):395–415
Barnard R, Leadley PW, Lensi R, Barthes L (2005) Plant, soil microbial and soil inorganic nitrogen responses to elevated CO2: a study in microcosms of Holcus lanatus. Acta Oecol 27(3):171–178
Brym A, Paerl HW, Montgomery MT, Handsel LT, Ziervogel K, Osburn CL (2014) Optical and chemical characterization of base-extracted particulate organic matter in coastal marine environments. Mar Chem 162:96–113
Chang CM (2019) Rationalization and prediction of the impact of different metals and root exudates on carbon dioxide emission from soil. Sci Total Environ 691:348–359
Conthe M et al (2019) Denitrification as an N2O sink. Water Res 151:381–387
Fan F, Zhang B, Morrill PL (2017) Phospholipid fatty acid (PLFA) analysis for profiling microbial communities in offshore produced water. Mar Pollut Bull 122(1–2):194–206
Fraterrigo JM, Balser TC, Turner MG (2006) Microbial community variation and its relationship with nitrogen mineralization in historically altered forests. Ecology 87(3):570–579
Frostegård A, Bååth E (1996) The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol Fertil Soils 22(1–2):59–65
Gao J, Song Z, Liu Y (2019) Response mechanisms of leaf nutrients of endangered plant (Acer catalpifolium) to environmental factors varied at different growth stages. Glob Ecol Conserv 17:e00521
Gorman-Lewis D, Martens-Habbena W, Stahl DA (2019) Cu(II) adsorption onto ammonia-oxidizing bacteria and archaea. Geochim Cosmochim Acta 255:127–143
Guo W, Jia G, Ye F, Xiao H, Zhang Z (2019) Lipid biomarkers in suspended particulate matter and surface sediments in the Pearl River estuary, a subtropical estuary in southern China. Sci Total Environ 646:416–426
Hu S, Chapin FS, Firestone MK, Field CB, Chiariello NR (2001) Nitrogen limitation of microbial decomposition in a grassland under elevated CO2. Nature 409(6817):188–191
Hungate BA (1999) Ecosystem responses to rising atmospheric CO2. In: Luo Y, Mooney HA, eds. Carbon Dioxide and Environmental Stress. Academic Press, San Diego, pp 265–285
James RT, Martin J, Wool T, Wang PF (1997) A sediment resuspension and water quality model of Lake Okeechobee. J Am Water Resour Assoc 33(3):661–678
Jensen HS, Andersen FO (1992) Importance of temperature, nitrate, and pH for phosphate release from aerobic sediments of four shallow, eutrophic lakes. Limnol Oceanogr 37(3):577–589
Jinbo Z, Changchun S, Shenmin W (2007) Dynamics of soil organic carbon and its fractions after abandonment of cultivated wetlands in Northeast China. Soil Tillage Res 96(1–2):350–360
Kang H, Kim SY, Fenner N, Freeman C (2005) Shifts of soil enzyme activities in wetlands exposed to elevated CO2. Sci Total Environ 337(1–3):207–212
Kelly JJ, Peterson E, Winkelman J, Walter TJ, Rier ST, Tuchman NC (2013) Elevated atmospheric CO2 impacts abundance and diversity of nitrogen cycling dunctional genes in soil. Microb Ecol 65(2):394–404
Laut L et al (2020) Organic matter compounds as a tool for trophic state characterization in a hypersaline environment: Araruama lagoon, Brazil. J S Am Earth Sci 97:102403
Li X, Fan F, Zhang B, Zhang K, Chen B (2018) Biosurfactant enhanced soil bioremediation of petroleum hydrocarbons: design of experiments (DOE) based system optimization and phospholipid fatty acid (PLFA) based microbial community analysis. Int Biodeterior Biodegrad 132:216–225
Lipson D, Näsholm T (2001) The unexpected versatility of plants: organic nitrogen use and availability in terrestrial ecosystems. Oecologia 128(3):305–316
Liu J, Appiah-Sefah G, Apreku TO (2018) Effects of elevated atmospheric CO2 and nitrogen fertilization on nitrogen cycling in experimental riparian wetlands. Water Sci Eng 11(1):39–45
Madsen TV, Sandjensen K (1994) The interactive effects of light and inorganic carbon on aquatic plant growth. Plant Cell Environ 17(8):955–962
Maltais-Landry G, Maranger R, Brisson J, Chazarenc F (2009) Greenhouse gas production and efficiency of planted and artificially aerated constructed wetlands. Environ Pollut 157(3):748–754
Mikan CJ, Zak DR, Kubiske ME, Pregitzer KS (2000) Combined effects of atmospheric CO2 and N availability on the belowground carbon and nitrogen dynamics of aspen mesocosms. Oecologia 124(3):432–445
Niklaus PA, Spinnler D, Körner C (1998) Soil moisture dynamics of calcareous grassland under elevated CO2. Oecologia 117(1–2):201–208
Nord EA, Jaramillo RE, Lynch JP (2015) Response to elevated CO2 in the temperate C3 grass Festuca arundinaceae across a wide range of soils. Front Plant Sci 6:96
Pang D et al (2019) Responses of soil labile organic carbon fractions and stocks to different vegetation restoration strategies in degraded karst ecosystems of Southwest China. Ecol Eng 138:391–402
Satake K, Shimura S (1983) Carbon dioxide assimilation from air and water by duckweed Spirodela polyrrhiza (L.) Schleid. Hydrobiologia 107(1):51–55
Satoh H, Nakamura Y, Okabe S (2007) Influences of infaunal burrows on the community structure and activity of ammonia-oxidizing bacteria in intertidal sediments. Appl Environ Microbiol 73(4):1341–1348
Smolander A, Kitunen V (2002) Soil microbial activities and characteristics of dissolved organic C and N in relation to tree species. Soil Biol Biochem 34(5):651–660
Song L, Wu J, Li C, Li F, Peng S, Chen B (2009) Different responses of invasive and native species to elevated CO2 concentration. Acta Oecol 35(1):128–135
Sowerby A, Blum H, Gray TRG, Ball AS (2000) The decomposition of Lolium perenne in soils exposed to elevated CO2: comparisons of mass loss of litter with soil respiration and soil microbial biomass. Soil Biol Biochem 32(10):1359–1366
Tzanakakis VA, Taylor AE, Bakken LR, Bottomley PJ, Myrold DD, Dörsch P (2019) Relative activity of ammonia oxidizing archaea and bacteria determine nitrification-dependent N2O emissions in Oregon forest soils. Soil Biol Biochem 139:107612
Walter BP, Heimann M (2000) A process-based, climate-sensitive model to derive methane emissions from natural wetlands: application to five wetland sites, sensitivity to model parameters, and climate. Glob Biogeochem Cycles 14(3):745–765
Wang J, Zhu T, Ni H, Zhong H, Fu X, Wang J (2013) Effects of elevated CO2 and nitrogen deposition on ecosystem carbon fluxes on the Sanjiang plain wetland in Northeast China. PLoS One 8:e66563
Wrage N, Velthof GL, Van Beusichem ML, Oenema O (2001) Role of nitrifier denitrification in the production of nitrous oxide. Soil Biol Biochem 33:1723–1732
Xing W, Wang Y, Hao T, He Z, Jia F, Yao H (2020) pH control and microbial community analysis with HCl or CO2 addition in H-2-based autotrophic denitrification. Water Res 168:115200
Yang S et al (2019) Long-term elevated CO2 shifts composition of soil microbial communities in a Californian annual grassland, reducing growth and N utilization potentials. Sci Total Environ 652:1474–1481
Yu T, Chen Y (2019) Effects of elevated carbon dioxide on environmental microbes and its mechanisms: a review. Sci Total Environ 655:865–879
Zhang H, Wang L, Li Y, Wang P, Wang C (2019a) Background nutrients and bacterial community evolution determine 13C-17β-estradiol mineralization in lake sediment microcosms. Sci Total Environ 651:2304–2311
Zhang W, Zhang H, Jian S, Liu N (2019b) Tree plantations influence the abundance of ammonia-oxidizing bacteria in the soils of a coral island. Appl Soil Ecol 138:220–222
Zhao G, Liu J, Wang Y, Dou J, Dong X (2009) Effects of elevated CO2 concentration and nitrogen supply on biomass and active carbon of freshwater marsh after two growing seasons in Sanjiang plain, Northeast China. J Environ Sci 21(10):1393–1399
Acknowledgements
The research was supported by the Natural Science Fund Project of Jilin Provincial Department of Science and Technology in 2020, the Jilin Agricultural University National Undergraduate Entrepreneurship Program in 2018, and the Jilin Agricultural University National Undergraduate Innovation Program in 2019.
Funding
Not applicable
Author information
Authors and Affiliations
Contributions
Conceptualization: D.J., L.C., and N.X. Data collection and analysis: D.J., D.A.K., N.X., E.N., and W.K.D. Writing—original draft preparation: D.J., D.A.K., N.X., E.N., and W.K.D. Writing—review and editing: D.A.K., L.C., W.K.D., and E.N. Supervision: L.C. and W.K.D. Funding acquisition: L.C. All authors have read and agreed to the published version of the manuscript.
Authors’ information
Not applicable
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare no conflicts of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Jiang, D., Chen, L., Xia, N. et al. Elevated atmospheric CO2 impact on carbon and nitrogen transformations and microbial community in replicated wetland. Ecol Process 9, 57 (2020). https://doi.org/10.1186/s13717-020-00267-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13717-020-00267-0