
Abstract
Disentangling the processes that drive population, community and whole forest structure and dynamics is a challenge. It becomes a grand challenge in the tropics where there are a large number of species, small population sizes, less research infrastructure, and a relatively smaller number of researchers compared to the temperate zone. Tackling this grand challenge, we argue, requires detailed knowledge of the functioning of individuals and species. To this end, researchers frequently employ plant functional traits to study tree populations and communities. Here, we review this approach by first providing a basic background regarding the major trait axes generally of interest. We then discuss how these axes may be or have been applied from ecosystem to community and population studies. In doing so, we highlight where the functional trait research program has failed in tropical tree ecology and where it can be improved or strengthened. Finally, we provide a perspective regarding how functional trait and emerging ‘omics approaches can be integrated to address large questions facing the field. Our intention throughout is to provide an entryway into this literature for an early career researcher rather than a comprehensive review of all possible studies that have taken place in tropical forests.
Similar content being viewed by others

Background
Tropical forests are known for their vast complexity, which has long intrigued plant ecologists and evolutionary biologists alike (e.g., von Humboldt and Bonpland 1807; Wallace 1878; Dobzhansky 1950; Gentry 1988). This complexity ultimately arises from ecological interactions that drive evolutionary outcomes and vice versa. These ecological interactions are governed by how phenotypes and the abiotic and biotic environment interact. Thus, phenotypic differences between individuals and between species are of central importance for disentangling and understanding the drivers of tropical diversity (Swenson 2013). Measures of functional diversity are often employed to represent phenotypic differences across spatial, temporal, taxonomic, and size scales in tropical forests. Ultimately, our interest in functional diversity, in this review, is how it is related to demographic outcomes, which alter tree dynamics across scales from populations, to communities, to ecosystems (Ackerly et al. 2000; McGill et al. 2006; Baraloto et al. 2010). Calow (1987) wrote over three decades ago that to study functional ecology, physiological measurements that influence or can be influenced by demography and genetics would be needed. Today, functional trait measurements are often employed to investigate demographic drivers and their implications for diversity and dynamics of tropical trees (Poorter et al. 2008; Paine et al. 2011; Iida et al. 2014a; Liu et al. 2016). This link between functional traits and demography is fundamental to many areas of plant ecology including climate change and ecosystem services, conservation, community dynamics, and even plant ecological genomics (Swenson and Jones 2017; Garnier et al. 2018). Despite the great strides made in each of these areas for understanding tropical tree dynamics, the link between plant function, demography, and dynamics is still elusive.
Introduction
As first coined by Calow (1987) and Keddy (1992), functional traits are morphological-physiological-phenological traits measured at the individual level that directly impact fitness. Despite the large number of traits that could impact plant fitness or performance, the functional trait research program typically focuses on a set of commonly and easily measured traits (e.g., Westoby 1998; Westoby et al. 2002; Westoby and Wright 2006). It is thought that these traits represent fundamental tradeoff axes that indicate life history strategies, the abiotic and biotic interactions of individuals and species, and their overall fitness or performance given the abiotic and biotic contexts (Ackerly 2003; Swenson 2012; Adler et al. 2014; Funk et al. 2017; Amahowe et al. 2018). To begin the review, we will focus on the most commonly measured functional traits (i.e., the wood economics spectrum, the leaf economics spectrum, and seed economics spectrum) and later in the text we will expand our discussion of functional diversity to include a larger variety of traits and functions.
The wood economics spectrum (Chave et al. 2009), like other trait spectra, depicts where a species falls along a continuum of allocation strategies that reflect life history tradeoffs. Specifically, high investment per unit wood volume (i.e., higher wood density) is negatively correlated with volumetric growth and mortality rates. Similarly, low wood density is positively correlated with volumetric growth and mortality rates (Enquist et al. 1999; Chave et al. 2009; Wright et al. 2010; Swenson 2012). Wood density is also essential for robust estimates of carbon storage, which is key in conservation strategies and dynamic global vegetation modeling (Chave et al. 2009). In addition to these well-characterized aspects of the wood economics spectrum, wood density measurements have also been successfully correlated with stem breakage and boring insect resistance; events that are becoming more common with climate change (Zimmerman et al. 1994; van Gelder et al. 2006; Swenson 2012). For these reasons, wood density is perhaps one of the most consistently useful functional traits when assessing tree demography and forest dynamics in the tropics and elsewhere.
The leaf economics spectrum (LES) characterizes a tradeoff between photosynthetic rates and leaf lifespans where leaves with high photosynthetic rates tend to have cheaply constructed leaves with short lifespans and leaves with low photosynthetic rates tend to have leaves with high construction costs and long leaf lifespans (Reich et al. 1997; Wright et al. 2004). The traits used to characterize the LES are, typically, specific leaf area (SLA), leaf nitrogen content, leaf phosphorus content, and leaf lifespan. The SLA of a leaf is the area of one side of a fresh leaf divided by its oven-dry mass (m2 kg−1). It is also often represented as leaf mass per area (LMA) in the literature, which is simply 1/SLA (Cornelissen et al. 2003). Trees with high SLA, nitrogen, and phosphorus on one end of the spectrum are known to photosynthesize at a faster rate but have shorter leaf life spans, thus “cheap” investment in tissue but fast returns (Wright et al. 2004; Chave et al. 2009; Swenson 2012; Funk et al. 2017). The other end of the spectrum represents longer-lived, slower-growing trees with low SLA, nitrogen, and phosphorus (Wright et al. 2004; Donovan et al. 2011). Some work has shown that the wood economics spectrum is orthogonal to the leaf economics spectrum (Baraloto et al. 2010; Dwyer and Laughlin 2017; Amahowe et al. 2018). This contrasts with other work demonstrating a tight coordination between leaf and wood hydraulic traits (e.g., Santiago et al. 2004). There are multiple potential reasons for these discrepancies, but the standard practice should be to measure both axes.
Lastly, seed mass is a commonly measured functional trait in tropical trees that forms the basis of the seed economics spectrum. Specifically, seed mass represents where a species falls along a continuum of making many small seeds or very few large seeds. Small seeds are less well provisioned (i.e., less endosperm) making them unlikely to establish in resource limited environments (e.g., shaded understories). However, they are more likely to reach high resource environments (e.g., canopy gaps) and typically have faster growth rates. Conversely, large seeds are more well provisioned making them more likely to establish in resource poor environments, while experiencing low growth and mortality rates (e.g., Grubb 1977). Thus, seed mass should be a leading indicator of plant life history strategies (e.g., Venable 1996; Moles et al. 2004). Indeed, seed mass is often one of the more informative traits tropical tree ecologists have studied (e.g., Wright et al. 2010), but it remains one of the hardest to measure traits due to difficulty in collecting seeds from many species with low population sizes and infrequent flowering events.
At present, measuring functional traits is the standard approach when investigating tree demography (Poorter et al. 2008; Wright et al. 2010; Paine et al. 2015). Along with the large functional trait literature, more detailed investigations of physiology exist and were more common place in the context of tree demographic modeling in the 1980s and 1990s. For example, the most detailed compilation of work on the functional biology of tropical trees, focused only on interactions between the environment and a single or a handful of species at a time, was edited by Mulkey et al. (1996). Many of the studies in that volume focused on gas exchange or other metabolic fluxes as ecophysiological traits of interest, which are difficult to obtain in the field for large numbers of individuals (Swenson 2013). During the early 1990s, studies were also investigating aspects of plant function along environmental gradients (Ellsworth and Reich 1992; Kitajima 1992; Osunkoya et al. 1994). For example, Kitajima (1996) focused on the effects of seed mass for survival and quantified early forms of what are now popularized functional traits along a shade tolerant to light demanding spectrum. Other studies were finding that seedlings with high specific leaf area had shorter leaf life spans, faster leaf turnover rates, and high photosynthetic rates (Bongers and Popma 1990; Kitajima 1994). Thus, while most of the current focus on the functional biology of tropical trees circles around a few easily measured traits, there is a rich body of literature on more detailed physiological measurements. In the following, we will discuss the functional trait literature, but push the reader to consider enhancing functional trait research with more detailed physiological and genomic measurements of the functional diversity of tropical trees.
Below, we highlight three main areas where functional traits and their linkages to tree demography and forest dynamics have had large impacts: climate and ecosystem change, community dynamics, and ecological genomics. We will guide the reader through trait-based research in these areas, give examples of detailed analyses, and highlight research shortcomings and future progressions.
Climate and ecosystem change
The relationship between functional traits and the demographic rates (i.e., growth rate, survival, reproduction) of tropical trees is dependent on the environmental context. Thus, any change in the environment will alter the demographic rates of trees and the functional composition of forests and forest dynamics. The environment is currently changing at unprecedented rates and extreme events such as drought, fire, insect and pathogen outbreaks, and hurricanes that greatly impact forest composition and dynamics are becoming a routine (Bigler et al. 2007; Jentsch et al. 2007; Chave et al. 2009; Choat et al. 2012; Esquivel-Muelbert et al. 2017; McDowell et al. 2018). With the increase in major events, researchers are looking for ways to better predict the future climate and the fate of tropical forests. A knowledge of how traits relate to demographic rates and how these relationships are modified by the environmental context is, therefore, essential for making such predictions.
Vegetation modeling
As the effects of climate change become more apparent, researchers are striving to predict future climatic and vegetation patterns. Dynamic global vegetation models (DGVMs) are often central to these predictions. Current DGVMs vary in their detail regarding the functional diversity in tropical forests with some including only one to a few functional types rather than a continuous distribution of functions (Díaz and Cabido 1997; Baraloto et al. 2010; Xu et al. 2016; Butler et al. 2017; Maréchaux and Chave 2017). While some inclusion of functional types is better than excluding plant function all together, such models fail to capture trait variation and should break apart these functional types into specific functional trait measurements for each species to incorporate more axes of plant function and continuous variation in traits (Wright et al. 2005; Baraloto et al. 2010; Funk et al. 2017). The challenge going forward is to develop a new generation of DGVMs that can deal with this continuous variation and models that can utilize both widely measured functional trait information (e.g., Fyllas et al. 2012, 2014; Maréchaux and Chave 2017) and less commonly measured physiological (Xu et al. 2016) and functional genomic measurements (Swenson 2012; Swenson and Jones 2017).
Along with predicting future climate, many of these models have the potential to predict relative abundance of species in designated areas (Laughlin and Laughlin 2013; Funk et al. 2017). This information would give insight into the resistance and resilience of different species (i.e., their capacity to maintain populations to alternations in the environment) to changes in climate and local adaptation capacity of species. Climate-based species distribution models currently predict the extensive redistributions of some tree species over the next century (Aitken et al. 2008). However, many of the predictions made by these models do not account for the actual ability of species to migrate or locally adapt (Aitken et al. 2008). Improved accuracy of these models can be gained with a better understanding of differential demography of species that incorporates phenotypic variation and abiotic and biotic interactions (Díaz and Cabido 1997).
Longitudinal studies
The parameterization of DGVMs can benefit greatly from long-term forest monitoring where tree demographic rates are tracked in detail. In those instances where traits can be linked to species (e.g., Iida et al. 2014a), or even individuals (e.g., Liu et al. 2016; Umaña et al. 2017a; Umaña et al. 2018), this information should and can be fed back into the DGVM parameterization process. Unfortunately, such long-term studies are often lacking. Those that do exist often observe individuals at 5-year intervals (e.g., Kraft et al. 2010; Iida et al. 2016), which does not allow the research to identify the impacts of acute and extreme events. Thus, going forward, observational studies of individuals on, at most, yearly time scales coupled with, preferably, individual level trait measurements would be a necessary next step for delineating how traits and demographic rates are related as the environment changes. One potential outcome of such work could be that individual level trait or rate measurements are not necessary for powerful modeling of future forest states, but individual level studies should first be conducted to remove this possibility or at least to provide more detailed prior distributions.
Community dynamics
Community ecology has taken the greatest advantage of the functional traits research program thus far. Specifically, functional traits have been used by community ecologists to elucidate and decipher patterns of species richness, abundance, and demography over space and time (e.g., Kitajima 1994; Mulkey et al. 1996; Swenson and Enquist 2007; Kraft et al. 2008; Poorter et al. 2008; Swenson and Enquist 2009; Wright et al. 2010; Katabuchi et al. 2012; Paine et al. 2012; Baraloto et al. 2012; Umaña et al. 2016, 2017). For example, Swenson and Enquist (2007) published one of the first analyses of community-wide diversity of a functional trait, wood density, in co-occurring tropical tree species demonstrating that assemblages are more diverse in this trait than expected given their high species richness. From here, a multitude of studies investigating environmental filtering (e.g., Swenson and Enquist 2009; Kraft and Ackerly 2010; Shipley 2010; Katabuchi et al. 2012; Kunstler et al. 2012) versus niche partitioning (Baraloto et al. 2006; Kraft et al. 2008; Paine et al. 2011; Lasky et al. 2015) using trait dispersion patterns were published. While important, these early studies typically focused on static snapshots of a forest composition and did not include temporal dynamics or demographic rates. More recently, this has changed and a greater focus has been placed on how traits are related to demography and ultimately forest dynamics (e.g., Alvarez-Clare and Kitajima 2007; Poorter et al. 2008; Aiba and Nakashizuka 2009; Uriarte et al. 2010; Chaturvedi et al. 2011; Fan et al. 2012; Swenson et al. 2012; Iida et al. 2014; Paine et al. 2015; Liu et al. 2016; Poorter et al. 2018; Rüger et al. 2018).
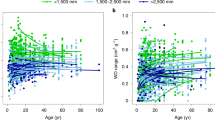
Foundational to nearly all trait-based studies in community ecology is the expectation that the traits measured are related to individual-level demographic performance. Thus, correlations between a single trait of interest and some environmental variable or demographic rate have been performed to explore co-occurrence and try to explain the vast diversity of trees seen in the tropics. However, in many instances, the relationships found between functional traits and these variables are often weak (Baraloto et al. 2012; Paine et al. 2015; Yang et al. 2018) and unable to explain the great diversity of form and function seen (Marks and Lechowicz 2006). For example, Paine et al. (2015) modeled the effects of functional traits (SLA, wood density, and seed mass) and potential evapotranspiration (PET) on size-standardized relative height growth rates (SGR) of 278 species globally and found that functional traits only explained 3.1% of the variation. Poorter et al. (2008) reported Pearson correlation coefficients between functional traits and relative diameter growth rate and mortality rate for five neotropical forests and found between 2 and 74% of the variation explained depending on the trait and location. The range in the amount of variation explained draws attention to the complexity that each forest brings to trait-rate relationships (Fig. 1a). A low percentage of variation explained (r²) is a common theme in functional trait tropical plant community ecology (Fig. 1b). This is alarming as it may indicate that the foundation of functional trait-based tropical tree ecology is far weaker than commonly portrayed. Thus, a major current challenge in tropical tree ecology is to clearly identify the axes of function that most clearly predict tree performance and to identify the reasons why previous work has explained so little variation. It has been stated that weak trait-demography relationships stem from a lack of consideration of the environment (Paine et al. 2015; Garnier et al. 2018; Yang et al. 2018), ignoring individuals in favor of species (Liu et al. 2016; Yang et al. 2018) and/or not considering all relevant dimensions of tree function (e.g., Swenson 2013; Iida et al. 2016; Swenson et al. 2017; Garnier et al. 2018; Yang et al. 2018). In the following, we will discuss each of these possibilities in turn and how tropical tree community ecologists should investigate or consider each of them in the future.

Distribution of coefficient of determination values for tropical plant studies. a Range of coefficient of determination values taken from 33 studies (Additional file 1) that modeled growth rate of tropical plants with a single functional trait, either specific leaf area (SLA), leaf mass per area (LMA), leaf nitrogen content, leaf phosphorus content, wood density, or seed mass. b Frequency of coefficient of determination values for all studies considered
Lack of environmental consideration
At regional or larger scales, studies have shown that climate and soil affect the slopes and intercepts of trait-trait and trait-climate relationships (Wright and Westoby 2002; Wright et al. 2005; Maire et al. 2015), but few tropical studies have tested for environmental effects at smaller scales where most of the literature is focused on herbaceous systems (Freschet et al. 2010; Dwyer and Laughlin 2017). For example, Laughlin et al. (2018) showed the importance of including environment-trait interactions in models of plant survival by comparing the traditional likelihood approach, where the trait is a function of the environment, to the vital rates approach that includes an interaction term between the trait and the environment. This type of analysis must be expanded to tropical forest systems to support known theory that diversity and dynamics are driven by differential demographic rates that result from phenotype-environment interactions (Fonseca et al. 2000; Ackerly 2003; Cavender-Bares et al. 2004; HilleRisLambers et al. 2012; Anderson 2016) (Fig. 2). These future studies have the potential to address a long-standing challenge in community ecology of explaining the vast diversity of forms and functions of trees in the tropics.

Major areas of tropical tree ecology are studied by linking functional traits, demography, and the environment. All three of these contexts and their interactions must be considered in analyses to adequately explain differential performance across environments, identify the axes of function that clearly predict this performance, and explain the vast diversity of form and function of trees seen in the tropics. We argue that the greatest advances in trait-based studies of tropical forests in the future will come through an explicit consideration of how organismal-level function interacts with abiotic and biotic contexts to drive differential tree demography and ultimately forest dynamics. Without an explicit consideration of these contexts and richer assays of tree function via anatomical and physiological traits and functional genes, little additional progress may be made
Ignoring individuals
In most functional trait studies, mean trait values are used for each species with the belief being that intraspecific variation in these traits is relatively small compared to interspecific variation (Albert et al. 2011; Albert et al. 2012; Adler et al. 2013; Funk et al. 2017; Yang et al. 2018). However, species averages undermine the importance of intraspecific variation in processes of community assembly, coexistence, and ecosystem functions (Burton et al. 2017). For example, Messier et al. (2010) was one of the first to show the importance of intraspecific variation by finding approximately equal partitioning of variance in leaf dry matter content (LDMC) and leaf mass per area (LMA) within and among tree species. Furthermore, Umaña et al. (2015) has shown the importance of intraspecific variation in traits and growth rates on species abundance. Of course, we realize the difficulty in obtaining trait data for every individual in a study. However, when working at smaller spatial scales, having intraspecific variation is critically important (Albert et al. 2011). Having this individual level variation in functional traits will allow an understanding of individual performance thus linking demography and evolution (Anderson 2016).
Relevant dimensions of function
Our understanding of the interactions between multiple functional traits and plant performance was and still is a major challenge in community ecology (Ackerly et al. 2000; Dwyer and Laughlin 2017). In the majority of studies, however, traits are examined individually or summarized into single indices to decrease the dimensionality of an analysis. This approach contrasts with a literature calling for the inclusion of multiple functional traits when trying to understand plant demography (Ackerly et al. 2000; Ackerly 2003; Marks and Lechowicz 2006; Adler et al. 2014; Enquist et al. 2015; Dwyer and Laughlin 2017a, Dwyer and Laughlin 2017; D’Andrea et al. 2018). Recent work that incorporates multiple functional traits into analyses of demography has met this call, albeit typically in temperate herbaceous systems (Laughlin and Messier 2015; Dwyer and Laughlin 2017).
The use of allometries should become more common to circumvent this issue by integrating functional trait measures with whole-plant allocation. Scaling relationships influence many aspects of plant form and function as they are controlled by the physiological and architectural constraints of the plant (Enquist and Niklas 2002; Bonser 2006). For example, plant growth rate is known to scale isometrically with leaf biomass (Niklas and Enquist 2002; Niklas and Enquist 2002a). Yang et al. (2018) have demonstrated that by knowing the allocational context of LMA, the trait-rate relationship is greatly improved. Unfortunately, we currently lack the necessary information for most species-specific allometries, especially for tropical species, and general allometric equations must be used. Remote sensing offers one potential alternative way of solving this issue as it could generate rapid inventories of crown dimensions for entire study sites that could be integrated with leaf level traits.
A related issue regarding dimensions of function asks, “What dimensions or functional traits are actually relevant and are we actually measuring them?”. The most commonly used functional traits are actually indirect measures of physiological rates or “soft functional traits,” which could be a major issue in why the percent of variation explained by functional traits of vital rates is so low (Hodgson et al. 1999; Weiher et al. 1999; Cornelissen et al. 2003; Wright et al. 2010; Swenson 2012; Funk et al. 2017) (Table 1). Incorporation of “hard functional trait” measurements, such as photosynthetic rate, stem and leaf hydraulics, and chemical defenses, that directly measure a rate or process into data collection has seen a slow increase (Hodgson et al. 1999; Weiher et al. 1999; Lavorel and Garnier 2002). This is expected as many of these traits are time-consuming to measure and costlier to obtain (Paine et al. 2015).
Ecological genomics
Along with the rapid increase in the usage of functional traits in ecology has been the increase in DNA and RNA sequencing capacity. This increased capacity has also come at a reduction in cost that makes true ecological genomics a reality in temperate and tropical systems. We envision that the ‘omics revolution can and should be immediately integrated with research into functional diversity, demography, and forest dynamics. This integration should be accomplished on many fronts. This includes forging the integration of data regarding intraspecific trait and genetic variation that is so clearly absent and desperately needed (Crutsinger 2016). Investigating the genetic basis of trait variation has the potential to provide understanding into the physiological processes that allow species to persist globally (Akman et al. 2016). Another clear opportunity for transformative science will come from the integration of transcriptomics and functional ecology where detailed assays of functional genes and their expression can now be obtained (e.g., Swenson 2012; Swenson et al. 2017; Swenson and Jones 2017). In the following, we will describe some of the research that has already been conducted in these realms as well as immediate opportunities for investigation.
Transcriptomes
A major limitation of trait-based studies is the measurement of only a handful of traits. This has two downstream consequences. First, traits or axes of function of interest that are difficult to measure (e.g., defense) are not measured. Second, axes of function that are important but not considered important or totally unknown are not measured. Transcriptomics, RNA sequencing of a tissue or tissues, assays the functional genes expressed in a sample and returns the abundances of transcripts, which offers to transform functional ecology in the next decade (Swenson 2012). This will be largely driven by falling sequencing costs, increases in cyber-infrastructure, and, most importantly, comfort dealing with analytical tools designed to work with non-model system data. Most ecological genomics or transcriptomics studies to date have been severely limited to a few model (or near model) systems and/or in highly controlled experimental settings both of which fail to represent the complexity of ecological systems. However, recent work has been accomplished that compares many species simultaneously and successfully translates the results to ecological questions of general interest. For example, multiple studies have now investigated homologous gene similarities as it relates to drought effects (Kobayashi et al. 2013; Swenson et al. 2017), light responses (Han et al. 2017), and even defense (Zambrano et al. 2017). Specifically, Zambrano et al. (2017) annotated homologous gene trees for plant defense responses across 21 temperate species to test the Janzen-Connell hypothesis on species coexistence. Bowsher et al. (2017) determined differential gene expression in competition experiments for co-occurring Trifolium species to highlight potential niche axes differentiation that allow species to coexist and Kobayashi et al. (2013) investigated the environmental influence on mass flowering of the tropical tree Shorea beccariana [Dipterocarpaceae].
While less common, studies of gene expression (rather than just comparisons of homologous or orthologous gene sequence similarity) are likely to become particularly important in functional ecology and linking function to demographic outcomes. The relationship between genes, their expression, and phenotypes is complex, but can provide insight into understanding how ecological and evolutionary processes affect phenotype (Hersch-Green et al. 2011; Alvarez et al. 2015). This relationship is especially important for individuals or species showing alternative phenotypes where gene expression can potentially help explain the divergence in phenotypes, resource strategies of each type, and demographic outcomes (Alvarez et al. 2015). While identifying the link between genotype and phenotype is still impractical for tropical trees, gene expression patterns can highlight candidate genes that are associated with ecological effects.
The downside of transcriptome analysis is perhaps, at the same time, its upside; many genes that appear related to observed ecological patterns may currently have no known function, but this allows candidate genes or gene families to be identified as being of ecological interest for future research (Swenson 2012; Alvarez et al. 2015; Todd et al. 2016). Studies that analyze gene expression patterns in tropical trees are rare due to the difficulty of collecting samples for RNA isolation in the field and, in general, the high costs of RNA-based research (Akman et al. 2016). Fortunately, costs of gene expression studies are steadily declining making future application to tropical trees promising.
Genotyping and genomics
The uses of genotype information in plant ecology are broad including its uses in understanding the abundance, composition and diversity of herbivores and arthropod species (Crutsinger et al. 2006), plant performance and coexistence (Crutsinger et al. 2008; Shao et al. 2018), and ecosystem services (Hersch-Green et al. 2011; Zytynska et al. 2011). One key aspect of genotypic data is potentially having the proportion of genetic variation which can be linked to functional trait variation and responses to the environment (Albert et al. 2011). Of course, untangling genetic (G) and environmental (E) influences on functional traits is a major undertaking causing a dearth in tropical tree G × E experiments (Alberto et al. 2013; Crutsinger 2016). In general, there has been a lack of integration between community genetics studies and functional trait studies despite repeated calls for research that considers both simultaneously (Díaz and Cabido 2001; Hughes et al. 2008; Hersch-Green et al. 2011; Crutsinger 2016). The few studies that have taken on this challenge were able to dissect heritability of trait variation and the distribution of that variation in the community (Johnson and Agrawal 2005; McKown et al. 2014; Barbour et al. 2015). For example, to better understand and predict future species richness and abundance, researchers are analyzing patterns of phenotypic plasticity and local adaptation using functional traits and genotype information with the idea that plastic ability will increase the odds of survival as climate changes (Anderson 2016).
We expect that future genotyping efforts in tropical forest ecology will increasingly take advantage of assembled genomes. This likely will start with a single genome being sequenced for a given major lineage of tropical trees that can be used to design sequence capture probes across a genus and to annotate functional and non-functional genes whose variation is being quantified in population genomic studies. Thus, while the production of assembled genomes may occur for a myriad of reasons that are not immediately associated with ecological questions, these genomes likely can be immediately leveraged for detailed studies of tree demography and the relatedness and functional gene similarity of the individuals being studied. Such work will accelerate a synthesis of functional ecology, population and community ecology, and evolutionary ecology allowing the next generation of tropical field ecologist to address some of the most important questions facing biology.
Conclusion
In this review, we have shown how functional traits have and will continue to have large impacts in major areas of tropical plant ecology. Throughout the establishment of plant functional trait-based ecology, simplification of the field has been urged (Díaz et al. 1998). Despite the ease that this simplification has brought, we have now reached a point of expansion and integration of trait-based ecology which adds complexity to data acquisition and analyses (Clark 2010). As advances in functional trait-based ecology increase, however, it is important to remember the investment it takes to amass adequate data to carry out research in each of the areas mentioned above. At the core, tropical inventories of species and even individuals and their functional diversity are needed.
In order to reach a main goal of trait-based ecology, scaling from individuals to ecosystems, we must integrate and synthesize across multiple fields including physiology, population biology, community ecology, ecosystem ecology, and global ecology (Enquist et al. 2015). In particular, stronger incorporation of evolutionary thinking into all areas of tropical tree ecology is needed, especially if we hope to find the elusive link between functional traits, demography, and dynamics across scales (Garnier et al. 2018; Yang et al. 2018).
Abbreviations
- DGVM:
-
Dynamic global vegetation model
- E:
-
Environment
- G:
-
Genetic
- LDMC:
-
Leaf dry matter content
- LES:
-
Leaf economics spectrum
- LMA:
-
Leaf mass per area
- PET:
-
Potential evapotranspiration
- SGR:
-
Size-standardized relative height growth rates
- SLA:
-
Specific leaf area
References
Ackerly DD (2003) Community assembly, niche conservatism, and adaptive evolution in changing environments. Int J Plant Sci 164:S165–S184
Ackerly DD, Dudley SA, Sultan SE, Schmitt J, Coleman JS, Linder CR, Sandquist DR, Geber MA, Evans AS, Dawson TE, Lechowicz MJ (2000) The evolution of plant ecophysiological traits: recent advances and future directions. BioScience 50:979–995
Adler PB, Fajardo A, Kleinhesselink AR, Kraft NJB (2013) Trait-based tests of coexistence mechanisms. Ecol Lett 16:1294–1306
Adler PB, Salguero-Gómez R, Compagnoni A, Hsu JS, Ray-Mukherjee J, Mbeau-Che C, Franco M (2014) Functional traits explain variation in plant life history strategies. Proc Natl Acad Sci U S A 111:740–745
Aerts R (1990) Nutrient use efficiency in evergreen and deciduous species from heathlands. Oecologia 84:391–397
Aiba M, Nakashizuka T (2009) Growth properties of 16 non-pioneer rain forest tree species differing in sapling architecture. J Ecol 97:992–999
Aitken SN, Yeaman S, Holliday JA, Wang T, Curtis-McLane S (2008) Adaptation, migration or extirpation: climate change outcomes for tree populations. Evol Appl 1:95–111
Akman M, Carlson JE, Holsinger KE, Latimer AM (2016) Transcriptome sequencing reveals population differentiation in gene expression linked to functional traits and environmental gradients in the South African shrub Protea repens. New Phytol 210:295–309
Albert CH, de Bello F, Boulangeat I, Pellet G, Lavorel S, Thuiller W (2012) On the importance of intraspecific variability for the quantification of functional diversity. Oikos 121:116–126
Albert CH, Grassein F, Schurr FM, Vieilledent G, Violle C (2011) When and how should intraspecific variability be considered in trait-based plant ecology? Persp Plant Ecol Evol Sys 13:2017–2225
Alberto FJ, Aitken SN, Alía R, González-Martínez SC, Hänninen H, Kremer A, Lefévre F, Lenormand T, Yeaman S, Whetten R, Savolainen O (2013) Potential for evolutionary responses to climate change – evidence from tree populations. Glob Change Biol 19:1645–1661
Alvarez M, Schrey AW, Richards CL (2015) Ten years of transcriptomics in wild populations: what have we learned about their ecology and evolution? Mol Ecol 24:710–725
Alvarez-Clare S, Kitajima K (2007) Physical defence traits enhance seedling survival of neotropical tree species. Funct Ecol 21:1044–1054
Amahowe IO, Gaoue OG, Natta AK, Piponiot C, Zobi IC, Hérault B (2018) Functional traits partially mediate the effects of chronic anthropogenic disturbance on the growth of a tropical tree. AoB PLANTS 10:ply036. https://doi.org/10.1093/aobpla/ply036
Anderson JT (2016) Plant fitness in a rapidly changing world. New Phytol 210:81–87
Angert AL, Huxman TE, Barron-Gafford GA, Gerst KL, Venable DL (2007) Linking growth strategies to long-term population dynamics in a guild of desert annuals. J Ecol 95:321–331
Baraloto C, Bonal D, ,Goldberg DE (2006) Differential seedling growth response to soil resource availability among nine neotropical tree species. J Trop Ecol 22:487–497
Baraloto C, Hardy OJ, Paine CET, Dexter KG, Cruaud C, Dunning LT, Gonzalez M, Molino J, Sabatier D, Savolainen V, Chave J (2012) Using functional traits and phylogenetic trees to examine the assembly of tropical tree communities. J Ecol 100:690–701
Baraloto C, Paine CET, Poorter L, Beauchene J, Bonal D, Domenach A, Hérault B, Patiño S, Roggy J, Chave J (2010) Decoupled leaf and stem economics in rain forest trees. Ecol Lett 13:1338–1347
Barbour MA, Rodriguez-Cabal MA, Wu ET, Julkunen-Tiitto R, Ritland CE, Miscampbell AE, Jules ES, Crutsinger GM (2015) Multiple plant traits shape the genetic basis of herbivore community assembly. Funct Ecol 29:995–1006
Bigler C, Gavin D, Gunning C, Veblen TT (2007) Drought induces lagged tree mortality in a subalpine forest in the Rocky Mountains. Oikos 116:1983–1994
Blackman GE, Wilson GE (1951) Physiological and ecological studies in the analysis of plant environment: VII. An analysis of the differential effects of light intensity on the net assimilation rate, leaf-area ratio, and relative growth rate of different species. Ann Bot 15:373–408
Bongers F, Popma J (1990) Leaf dynamics of seedlings of rain forest species in relation to canopy gaps. Oecologia 82:122–127
Bonser SP (2006) Form defining function: interpreting leaf functional variability in integrated plant phenotypes. Oikos 114:187–190
Bowsher A, Shetty P, Anacker BL, Siefert A, Strauss S, Friesen ML (2017) Transcriptomic responses to conspecific and congeneric competition in co-occurring Trifolium. J Ecol 105:602–615
Burton JI, Perakis SS, McKenzie SC, Lawrence CE, Puettmann KJ (2017) Intraspecific variability and reaction norms of forest understorey plant species traits. Funct Ecol 31:1881–1893
Butler EE, Datta A, Flores-Moreno H, Chen M, Wythers KR, Fazayeli F, Banerjee A, Atkin OK, Kattge J, Amiaud B, Blonder B, Boenisch G, Bond-Lamberty B, Brown KA, Byun C, Campetella G, Cerabolini BEL, Cornelissen JHC, Craine JM, Craven D, de Vries FT, Díaz S, Domingues TF, Forey E, González-Melo A, Gross N, Han W, Hattingh WN, Hickler T, Jansen S, Kramer K, Kraft NJB, Kurokawa H, Laughlin DC, Meir P, Minden V, Niinemets U, Onoda Y, Peñuelas J, Read Q, Sack L, Schamp B, Soudzilovskaia NA, Spasojevic MJ, Sosinski E, Thornton PE, Valladares F, van Bodegom PM, Williams M, Wirth C, Reich PB (2017) Mapping local and global variability in plant trait distributions. Proc Natl Acad Sci U S A 114:E10937–E10946
Calow P (1987) Towards a definition of functional ecology. Funct Ecol 1:57–61
Carlquist SJ (1975) Ecological strategies of xylem evolution. University of California Press, Berkeley
Cavender-Bares J, Kitajima K, Bazzaz FA (2004) Multiple trait associations in relation to habitat differentiation among 17 Floridian oak species. Ecol Monogr 74:635–662
Chaturvedi RK, Raghubanshi AS, Singh JS (2011) Leaf attributes and tree growth in a tropical dry forest. J Veg Sci 22:917–931
Chave J, Coomes D, Jansen S, Lewis SL, Swenson NG, Zanne AE (2009) Towards a worldwide wood economics spectrum. Ecol Lett 12:351–366
Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S, Bhaskar R, Bucci SJ, Field TS, Gleason SM, Hacke UG, Jacobsen AL, Lens F, Maherali H, Martínez-Vilalta J, Myr S, Mencuccini M, Mitchell PJ, Nardini A, Pittermann J, Pratt RB, Sperry JS, Westoby M, Wright IJ, Zaane AE (2012) Global convergence in the vulnerability of forests to drought. Nature 491:752–756
Clark JS (2010) Individuals and the variation needed for high species diversity in forest trees. Science 327:1129–1132
Cornelissen JHC, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich DE, Reich PB, ter Steege H, Morgan HD, van der Heijden MGA, Pausas JG, Poorter H (2003) A handbook of protocols for standardized and easy measurement of plant functional traits worldwide. Aust J Bot 51:335–380
Crutsinger GM (2016) A community genetics perspective: opportunities for the coming decade. New Phytol 210:65–70
Crutsinger GM, Souza L, Sanders NJ (2008) Intraspecific diversity and dominant genotypes resist plant invasions. Ecol Lett 11:16–23
Crutsinger GM, Collins MD, Fordyce JA, Gompert Z, Nice CC, Sanders NJ (2006) Plant genotypic diversity predicts community structure and governs an ecosystem process. Science 313:966–968.
D’Andrea R, Ostling A, O’Dwyer JP (2018) Translucent windows: how uncertainty in competitive interactions impacts detection of community pattern. Ecol Lett 21:826–835
Díaz S, Cabido M (1997) Plant functional types and ecosystem function in relation to global change. J Veg Sci 8:463–474
Díaz S, Cabido M (2001) Vive la difference: plant functional diversity matters to ecosystem processes. Trends Ecol Evol 16:646–655
Díaz S, Cabido M, Casanoves F (1998) Plant functional traits and environmental filters at a regional scale. J Veg Sci 9:113–122
Diemer M (1998) Life span and dynamics of leaves of herbaceous perennials in high-elevation environments: ‘news from the elephant’s leg’. Fun Ecol 12:413–425
Dobzhansky T (1950) Evolution in the tropics. Am Sci 38:208–221
Donovan LA, Maherali H, Caruso CM, Huber H, de Kroon H (2011) The evolution of the worldwide leaf economics spectrum. Trends Ecol Evol 26:88–95
Dwyer JM, Laughlin DC (2017) Constraints on trait combinations explain climatic drivers of biodiversity: the importance of trait covariance in community assembly. Ecol Lett 20:872–882
Dwyer JM, Laughlin DC (2017a) Selection of trait combinations along environmental gradients. J Veg Sci 28:672–673
Elger A, Willby NJ (2003) Leaf dry matter content as an integrative expression of plant palatability: the case of freshwater macrophytes. Fun Ecol 17:58–65
Ellsworth DS, Reich PB (1992) Leaf mass per area, nitrogen content and photosynthetic carbon gain in Acer saccharum seedlings in contrasting forest light environments. Fun Ecol 6:423–435
Enquist BJ, Niklas KJ (2002) Global allocation rules for patterns of biomass partitioning in seed plants. Science 295:1517–1520
Enquist BJ, Norberg J, Bonser SP, Violle C, Webb CT, Henderson A, Sloat LL, Savage VM (2015) Scaling from traits to ecosystems: developing a general trait driver theory via integrating trait-based and metabolic scaling theories. Adv Ecol Res 52:249–318
Enquist BJ, West GB, Charnov EL, Brown JH (1999) Allometric scaling of production and life-history variation in vascular plants. Nature 401:907–911
Esquivel-Muelbert A, Galbraith D, Dexter KG, Baker TR, Lewis SL, Meir P, Rowland L, da Costa ACL, Nepstad D, Phillips OL (2017) Biogeographic distributions of neotropical trees reflect their directly measured drought tolerances. Sci Rep 7:8334
Fan Z, Zhang S, Hao G, Slik JWF, Cao K (2012) Hydraulic conductivity traits predict growth rates and adult stature of 40 Asian tropical tree species better than wood density. J Ecol 100:732–741
Fonseca CR, McC Overton J, Collins B, Westoby M (2000) Shifts in trait-combinations along rainfall and phosphorus gradients. J Ecol 88:964–977
Freschet GT, Cornelissen JHC, van Logtestijn RSP, Aerts R (2010) Evidence of the ‘plant economics spectrum’ in a subarctic flora. J Ecol 98:362–373
Funk JL, Larson JE, Ames GM, Butterfield BJ, Cavender-Bares J, Firn J, Laughlin DC, Sutton-Grier AE, Williams L, Wright J (2017) Revisiting the holy grail: using plant functional traits to understand ecological processes. Biol Rev 92:1156–1173
Fyllas NM, Gloor E, Mercado LM, Sitch S, Quesada CA, Dominques TF, Galbraith DR, Torre-Lezama A, Vilanova E, Ramírez-Angulo H, Higuchi N, Neill DA, Silveira M, Ferreira L, Aymard GA, Malhi Y, Phillips OL, Lloyd J (2014) Analysing Amazonian forest productivity using a new individual and trait-based model (TFS v.1). Geosci Model Dev 7:1251–1269
Fyllas NM, Quesada CA, Lloyd J (2012) Deriving plant functional types for Amazonian forests for use in vegetation dynamics models. Persp Plant Ecol Evol Sys 14:97–110
Garnier E, Fayolle A, Navas M, Damgaard C, Cruz P, Hubert D, Richarte J, Autran P, Leurent C, Violle C (2018) Plant demographic and functional responses to management intensification: a long-term study in a Mediterranean rangeland. J Ecol 106:1363–1376
Gentry AH (1988) Changes in plant community diversity and floristic composition on environmental and geographical gradients. Ann Mo Bot Gard 75:1–34
Grubb PJ (1977) The maintenance of species richness in plant communities: the importance of the regeneration niche. Biol Rev 52:107–145
Han B, Umaña MN, Mi X, Liu X, Chen L, Wang Y, Liang Y, Wei W, Ma K (2017) The role of transcriptomics linked with responses to light environment on seedling mortality in a subtropical forest. China J Ecol 105:592–601
Hersch-Green EI, Turley NE, Johnson MTJ (2011) Community genetics: what have we accomplished and where should we be going? Phil Trans R Soc B 366:1453–1460
HilleRisLambers J, Adler PB, Harpole WS, Levine JM, Mayfield MM (2012) Rethinking community assembly through the lens of coexistence theory. Annu Rev Ecol Evol Syst 43:227–248
Hodgson JG, Wilson PJ, Hunt R, Grime JP, Thompson K (1999) Allocating C-S-R plant functional types: a soft approach to a hard problem. Oikos 85:282–294
Hughes AR, Inouye BD, Johnson MTJ, Underwood N, Vellend M (2008) Ecological consequences of genetic diversity. Ecol Lett 11:609–623
Iida Y, Kohyama TS, Swenson NG, Su S, Chen C, Chiang J, Sun I (2014a) Linking functional traits and demographic rates in a subtropical tree community: the importance of size dependency. J Ecol 102:641–650
Iida Y, Poorter L, Sterck F, Kassim AR, Potts MD, Kubo T, Kohyama TS (2014) Linking size-dependent growth and mortality with architectural traits across 145 co-occurring tropical tree species. Ecology 95:353–363
Iida Y, Sun I, Price CA, Chen C, Chen Z, Chiang J, Huang C, Swenson NG (2016) Linking leaf veins to growth and mortality rates: an example from a subtropical tree community. Ecol Evol 6:6085–6096
Jentsch A, Kreyling J, Beierkuhnlein C (2007) A new generation of climate-change experiments: events, not trends. Front Ecol Environ 5:365–374
Johnson MTJ, Agrawal AA (2005) Plant genotype and environment interact to shape a diverse arthropod community on evening primrose (Oenothera biennis). Ecology 86:874–885
Katabuchi M, Kurokawa H, Davies SJ, Tan S, Nakashizuka T (2012) Soil resource availability shapes community trait structure in a species-rich dipterocarp forest. J Ecol 100:643–651
Keddy PA (1992) A pragmatic approach to functional ecology. Funct Ecol 6:621–626
Kitajima K (1992) The importance of cotyledon functional morphology and patterns of seed reserve utilization for the physiological ecology of neotropical tree seedlings. Dissertation, University of Illinois, Urbana
Kitajima K (1994) Relative importance of photosynthetic traits and allocation patterns as correlates of seedling shade tolerance of 13 tropical trees. Oecologia 98:419–428
Kitajima K (1996) Ecophysiology of tropical tree seedlings. In: Mulkey SS, Chazdon RL, Smith AP (eds) Tropical forest plant ecophysiology. Chapman and Hall, New York
Kobayashi MJ, Takeuchi Y, Kenta T, Kume T, Diway B, Shimizu KK (2013) Mass flowering of the tropical tree Shorea beccariana was preceded by expression changes in flowering and drought-responsive genes. Mol Ecol 22:4767–4782
Kraft NJB, Ackerly DD (2010) Functional trait and phylogenetic tests of community assembly across spatial scales in an Amazonia forest. Ecol Monogr 80:401–422
Kraft NJB, Metz MR, Condit RS, Chave J (2010) The relationship between wood density and mortality in a global tropical forest data set. New Phytol 188:1124–1136
Kraft NJB, Valencia R, Ackerly DD (2008) Functional traits and niche-based tree community assembly in an Amazonian forest. Science 322:580–582
Kunstler G, Lavergne S, Courbaud B, Thuiller W, Vieilledent G, Zimmermann NE, Kattge J, Coomes DA (2012) Competitive interactions between forest trees are driven by species’ trait hierarchy, not phylogenetic or functional similarity: implications for forest community assembly. Ecol Lett 15:831–840
Lambers H, Poorter H (1992) Inherent variation in growth rate between higher plants: a search for physiological causes and ecological consequences. Adv Ecol Res 23:187–261
Lasky JR, Bachelot B, Muscarella R, Schwartz N, Forero-Montaña J, Hytch CJ, Swenson NG, Thompson J, Zimmerman JK, Uriarte M (2015) Ontogenetic shifts in trait-mediated mechanisms of plant community assembly. Ecology 96:2157–2169
Laughlin DC, Laughlin DE (2013) Advances in modeling trait-based plant community assembly. Trends Plant Sci 18:584–593
Laughlin DC, Messier J (2015) Fitness and multidimensional phenotypes in dynamic adaptive landscapes. Trends Ecol Evol 30:487–496
Laughlin DC, Strahan RT, Adler PB, Moore MM (2018) Survival rates indicate that correlations between community-weighted mean traits and environments can be unreliable estimates of the adaptive value of traits. Ecol Lett 21:411–421
Lavorel S, Garnier E (2002) Predicting changes in community composition and ecosystem functional from plant traits: revisiting the holy grail. Funct Ecol 16:545–556
Liu X, Swenson NG, Lin D, Mi X, Umaña MN, Schmid B, Ma K (2016) Linking individual-level functional traits to tree growth in a subtropical forest. Ecology 97:2396–2405
Maire V, Wright IJ, Prentice IC, Batjes NH, Bhaskar R, van Bodegom PM, Cornwell WK, Ellsworth D, Niinemets Ü, Ordonez A, Reich PB, Santiago LS (2015) Global effects on soil and climate on leaf photosynthetic traits and rates. Glob Ecol Biogeogr 24:706–717
Maréchaux I, Chave J (2017) An individual-based forest model to jointly simulate carbon and tree diversity in Amazonia: description and applications. Ecol Monogr 87:632–664
Marks CO, Lechowicz MJ (2006) Alternative designs and the evolution of functional diversity. Am Nat 167:55–66
McDowell N, Allen CD, Anderson-Teixeira K, Brando P, Brienen R, Chambers J, Christoffersen B, Davies S, Doughty C, Duque A, Espirito-Santo F, Fisher R, Fontes CG, Galbraith D, Goodsman D, Grossiord C, Hartmann H, Holm J, Johnson DJ, Kassim AR, Keller M, Koven C, Kueppers L, Kumagai T, Malhi Y, McMahon SM, Mencuccini M, Meir P, Moorcroft P, Muller-Landau HC, Phillips OL, Powell T, Sierra CA, Sperry J, Warren J, Xu C, Xu X (2018) Drivers and mechanisms of tree mortality in moist tropical forests. New Phytol 219:851–869
McGill BJ, Enquist BJ, Weiher E, Westoby M (2006) Rebuilding community ecology from functional traits. Trends Ecol Evol 21:178–185
McKown AD, Klápště J, Guy RD, Geraldes A, Porth I, Hannemann J, Friedmann M, Muchero W, Tuskan GA, Ehlting J, Cronk QCB, El-Kassaby YA, Mansfield SD, Douglas CJ (2014) Genome-wide association implicates numerous genes underlying ecological trait variation in natural populations of Populus trichocarpa. New Phytol 203:535–553
Messier J, McGill BJ, Lechowicz MJ (2010) How do traits vary across ecological scales? A case for trait-based ecology. Ecol Lett 13:838–848
Metz MR, Sousa WP, Valencia R (2010) Widespread density-dependent seedling mortality promotes species coexistence in a highly diverse Amazonian rain forest. Ecology 91:3675–3685
Moles AT, Falster DS, Leishman MR, Westoby M (2004) Small-seeded species produce more seeds per square metre of canopy per year, but not per individual per lifetime. J Ecol 92:384–396
Mulkey SS, Chazdon RL, Smith AP (1996) Tropical forest plant ecophysiology. Chapman and Hall, New York
Niklas KJ (1997) The evolutionary biology of plants. University of Chicago Press, Chicago
Niklas KJ, Enquist BJ (2002) On the vegetative biomass partitioning of seed plant leaves, stems, and roots. Am Nat 159:482–497
Niklas KJ, Enquist BJ (2002a) Canonical rules for plant organ biomass partitioning and annual allocation. Am J Bot 89:812–819
Osunkoya OO, Ash JE, Hopkins MS, Graham AW (1994) Influence of seed size and seedling ecological attributes on shade-tolerance of rain-forest tree species in Northern Queensland. J Ecol 82:149–163
Paine CET, Amissah L, Auge H, Baraloto C, Baruffol M, Bourland N, Bruelheide H, Daïnou K, de Gouvenain RC, Doucet J, Doust S, Fine PVA, Fortunel C, Haase J, Holl KD, Jactel H, Li X, Kitajima K, Koricheva J, Martínez-Garza C, Messier C, Paquette A, Philipson C, Piotto D, Poorter L, Posada JM, Potvin C, Rainio K, Russo SE, Ruiz-Jaen M, Scherer-Lorenzen M, Webb CO, Wright SJ, Zahawi RA, Hector A (2015) Globally, functional traits are weak predictors of juvenile tree growth, and we do not know why. J Ecol 103:978–989
Paine CET, Baraloto C, Chave J, Hérault B (2011) Functional traits of individual trees reveal ecological constraints on community assembly in tropical rain forests. Oikos 120:720–727
Paine CET, Stenflo M, Philipson D, Saner P, Bagchi R, Ong RC, Hector A (2012) Differential growth responses in seedlings of ten species of Dipterocarpaceae to experimental shading and defoliation. J Trop Ecol 28:377–384
Poorter H, Remkes C (1990) Leaf area ratio and net assimilation rate of 24 wild species differing in relative growth rate. Oecologia 83:553–559
Poorter L, Castilho CV, Schietti J, Oliveira RS, Costa FRC (2018) Can traits predict individual growth performance? A test in a hyperdiverse tropical forest. New Phytol 219:109–121
Poorter L, Wright SJ, Paz H, Ackerly DD, Condit R, Ibarra-Manríquez G, Harms KE, Licona JC, Martínez-Ramos M, Mazer SJ, Muller-Landau HC, Peña-Claros M, Webb CO, Wright IJ (2008) Are functional traits good predictors of demographic rates? Evidence from five neotropical forests. Ecology 89:1908–1920
Putz FE, Coley PD, Lu K, Montalvo A, Aiello A (1983) Uprooting and snapping of trees: structural determinants and ecological consequences. Can J For Res 13:1011–1020.
Reich PB, Walters MB, Ellsworth DS (1997) From tropics to tundra: global convergence in plant functioning. Proc Natl Acad Sci U S A 94:13730–13734
Reich PB, Walters MB, Ellsworth DS, Vose JM, Volin JC, Gresham C, Bowman WD (1998) Relationships of leaf dark respiration to leaf nitrogen, specific leaf area and leaf life-span: a test across biomes and functional groups. Oecologia 114:471–482
Reich PB, Walters MD, Ellsworth DS (1992) Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems. Ecol Monogr 62:365–392
Rüger N, Comita LS, Condit R, Purves D, Rosenbaum B, Visser MD, Wright SJ, Wirth C (2018) Beyond the fast-slow continuum: demographic dimensions structuring a tropical tree community. Ecol Lett 21:1075–1084
Santiago LS, Goldstein G, Meinzer FC, Fisher JB, Machado K, Woodruff D, Jones T (2004) Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees. Oecologia 140:543–550
Shao X, Brown C, Worthy SJ, Liu L, Cao M, Li Q, Lin L, Swenson NG (2018) Intra-specific relatedness, spatial clustering and reduced demographic performance in tropical rainforest trees. Ecol Lett 21:1174–1181
Shipley B (2010) From plant traits to vegetation structure: chance and selection in the assembly of ecological communities. Cambridge University Press, Cambridge
Swenson NG (2012) The functional ecology and diversity of tropical tree assemblages through space and time: from local to regional and from traits to transcriptomes. ISRN Forestry 2012:743617
Swenson NG (2013) The assembly of tropical tree communities – the advances and shortcomings of phylogenetic and functional trait analyses. Ecography 36:264–276
Swenson NG, Enquist BJ (2007) Ecological and evolutionary determinants of a key plant functional trait: wood density and its community-wide variation across latitude and elevation. Am J Bot 94:451–459
Swenson NG, Enquist BJ (2009) Opposing assembly mechanisms in a Neotropical dry forest: implications for phylogenetic and functional community ecology. Ecology 90:2161–2170
Swenson NG, Erickson DL, Mi X, Bourg NA, Forero-Montaña J, Ge X, Howe R, Lake JK, Liu X, Ma K, Pei N, Thompson J, Uriarte M, Wolf A, Wright SJ, Ye W, Zhang J, Zimmerman JK, Kress WJ (2012) Phylogenetic and functional alpha and beta diversity in temperate and tropical tree communities. Ecology 93:S112–S125
Swenson NG, Iida Y, Howe R, Wolf A, Umaña MN, Petprakob K, Turner BL, Ma K (2017) Tree co-occurrence and transcriptomic response to drought. Nat Commun 8:1996
Swenson NG, Jones FA (2017) Community transcriptomics, genomics and the problem of species co-occurrence. J Ecol 105:563–568
Todd EV, Black MA, Gemmell NJ (2016) The power and promise of RNA-seq in ecology and evolution. Mol Ecol 25:1224–1241
Umaña MN, Forero-Montaña J, Muscarella R, Nytch CJ, Thompson J, Uriarte M, Zimmerman J, Swenson NG (2016) Interspecific functional convergence and divergence and intraspecific negative density dependence underlie the seed-to-seedling transition in tropical trees. Am Nat 187:99–109
Umaña MN, Mi X, Cao M, Enquist BJ, Hao Z, Howe R, Iida Y, Johnson D, Lin L, Liu X, Ma K, Sun I, Thompson J, Uriarte M, Wang X, Wolf A, Yang J, Zimmerman JK, Swenson NG (2017) The role of functional uniqueness and spatial aggregation in explaining rarity in trees. Glob Ecol Biogeogr 26:777–786
Umaña MN, Zhang C, Cao M, Lin L, Swenson NG (2015) Commonness, rarity, and intraspecific variation in traits and performance in tropical tree seedlings. Ecol Lett 18:1329–1337
Umaña MN, Zhang C, Cao M, Lin L, Swenson NG (2017a) A core-transient framework for trait-based community ecology: an example from a tropical tree seedling community. Ecol Lett 20:619–628
Umaña MN, Zipkin EF, Zhang C, Cao M, Lin L, Swenson NG (2018) Individual-level trait variation and negative density dependence affect growth in tropical tree seedlings. J Ecol 106:2446–2455
Uriarte M, Swenson NG, Chazdon RL, Comita LS, Kress WJ, Erickson D, Forero-Montaña J, Zimmerman JK, Thompson J (2010) Trait similarity, shared ancestry and the structure of neighbourhood interactions in a subtropical wet forest: implications for community assembly. Ecol Lett 13:1503–1514
van Gelder HA, Poorter L, Sterck FJ (2006) Wood mechanics, allometry, and life-history variation in a tropical rain forest tree community. New Phytol 171:367–378
Venable L (1996) Packaging and provisioning in plant reproduction. Phil Trans R Soc Lond B 351:1319–1329
von Humboldt A, Bonpland A (1807) Essai sur la géographie des plantes. Schoell, Paris
Wallace AR (1878) Tropical nature and other essays. Macmillan and Company, London
Weiher E, van der Werf A, Thompson K, Roderick M, Garnier E, Eriksson O (1999) Challenging Theophrastus: a common core list of plant traits for functional ecology. J Veg Sci 10:609–620
Westoby M (1998) A leaf-height-seed (LHS) plant ecology strategy scheme. Plant Soil 199:213–227
Westoby M, Falster DS, Moles AT, Vesk PA, Wright IJ (2002) Plant ecological strategies: some leading dimensions of variations between species. Annu Rev Ecol Syst 33:125–159
Westoby M, Wright IJ (2006) Land-plant ecology on the basis of functional traits. Trends Ecol Evol 21:261–268
Wright IJ, Reich PB, Cornelissen JHC, Falster DS, Garnier E, Hikosaka K, Lamont BB, Lee W, Oleksyn J, Osada N, Poorter H, Villar R, Warton DI, Westoby M (2005) Assessing the generality of global leaf trait relationships. New Phytol 166:485–496
Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen JHC, Diemer M, Flexas J, Garnier E, Groom PK, Gulias J, Hikosaka K, Lamont BB, Lee T, Lee W, Luck C, Midgley JJ, Navas M, Niinemets Ü, Oleksyn J, Osada N, Poorter N, Poot P, Prior L, Pyankov VI, Roumet C, Thomas SC, Tjoelker MG, Veneklaas EJ, Villar R (2004) The worldwide leaf economics spectrum. Nature 428:821–827
Wright IJ, Westoby M (2002) Leaves at low versus high rainfall: coordination of structure, lifespan and physiology. New Phytol 155:403–416
Wright SJ, Kitajima K, Kraft NJB, Reich PB, Wright IJ, Bunker DE, Condit R, Dalling JW, Davies SJ, Díaz S, Engelbrecht BMJ, Harms KE, Hubbell SP, Marks CO, Ruiz-Jaen MC, Salvador CM, Zanne AE (2010) Functional traits and the growth-mortality trade-off in tropical trees. Ecology 91:3664–3674
Xu X, Medvigy D, Powers SJ, Becknell JM, Guan K (2016) Diversity in plant hydraulic traits explains seasonal and inter-annual variations of vegetation dynamics in seasonally dry tropical forests. New Phytol 212:80–95
Yang J, Cao M, Swenson NG (2018) Why functional traits do not predict tree demographic rates. Trends Ecol Evol 33:326–336
Zambrano J, Iida Y, Howe R, Lin L, Umana MN, Wolf A, Worthy SJ, Swenson NG (2017) Neighbourhood defense gene similarity effect on tree performance: a community transcriptomic approach. J Ecol 105:616–626
Zimmerman JK, Everham IIIEM, Waide RB, Lodge JD, Taylor CM, Brokaw NVL (1994) Responses of tree species to hurricane winds in subtropical wet forest in Puerto Rico: implications for tropical tree life histories. J Ecol 82:911–922
Zytynska SE, Fay MF, Penney D, Preziosi RF (2011) Genetic variation in a tropical tree species influences the associated epiphytic plant and invertebrate communities in a complex forest ecosystem. Phil Trans R Soc B 366:1329–1336
Acknowledgements
We thank two anonymous reviewers of the manuscript for constructive comments that improved the quality of our writing. We thank the University of Maryland for ongoing research support.
Funding
SJW and NGS were funded by a US National Science Foundation grant (EF-1638488) awarded to NGS.
Availability of data and materials
Not applicable
Author information
Authors and Affiliations
Contributions
SJW wrote the initial drafts of the manuscript with comments from NGS. SJW and NGS worked together to form the ideas and framework for the manuscript. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional file
Additional file 1:
Reference list of the 33 studies used in the production of Fig. 1. (DOCX 22 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Worthy, S.J., Swenson, N.G. Functional perspectives on tropical tree demography and forest dynamics. Ecol Process 8, 1 (2019). https://doi.org/10.1186/s13717-018-0154-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13717-018-0154-4