
Abstract
Introduction
Tanglehead is a grass native to southwestern US rangelands; however, its prevalence as a native invasive on South Texas rangelands has increased rapidly during the last decade. Large areas of monotypic stands have emerged in Jim Hogg, Duval, Brooks, and Kleberg counties. The aim of this research is to understand the spatial and temporal dynamics of these invasions as a model for the assessment of native invasive species. Our specific objectives were to (1) evaluate the feasibility of classifying tanglehead using 1-m resolution imagery data, (2) assess the spatial and temporal distribution of tanglehead in relation to soil type and distance from roads, and (3) quantify the temporal and spatial distribution of tanglehead on our study sites. We combined remote sensing approaches with landscape metrics to quantify the spatial and temporal distribution of tanglehead in five locations across our study area. We calculated the normalized difference vegetation index and combined it with the original aerial imagery to conduct an unsupervised classification with the following land cover classes: woody vegetation, tanglehead, non-tanglehead herbaceous, and bare ground. Soil type and the distance from roads were assessed to determine the relationship between these factors and tanglehead spatial distribution.
Results
We were able to successfully map tanglehead using the 1-m imagery. Our image classification approach resulted in accuracies greater than 85% for all sites. Tanglehead occurred in sandy, loamy sand, and sandy loam soils. Over 70% of tanglehead cover occurred within the first 150 m from the edge of roads. This cover increased from 7.1% (SE = 1.1%) in 2008 to 17.8% (SE = 5.4%) in 2014. Once established, small patches of tanglehead began aggregating or coalescing with existing stands, thereby creating larger patches over larger areas.
Conclusions
Our study has shown the value of analyzing spatiotemporal dynamics of tanglehead with remote sensing techniques and landscape metrics to improve our understanding of establishment and dispersal processes of a native invasive. This study provides useful information to improve rangeland management decisions as well as assessing native invasive dynamics with potential applications for assessing its effects on wildlife habitat, livestock operations, and habitat restoration strategies.
Similar content being viewed by others

Introduction
Invasive plants pose a serious threat to native flora and fauna, often having effects that can be long term and irreversible once established across the landscape (Flanders et al. 2006). Invasives are capable of altering plant-community structure and composition, as well as ecosystem processes such as nutrient cycling and fire regimes (Fulbright et al. 2013). These changes can cause cascading effects on many species of wildlife as they rely on grasses and forbs for food, cover, and other resources essential to their survival. For example, areas in Australia that have been subjected to the invasion of gamba grass (Andropogon gayanus Kunth.), a high biomass producing perennial, have shown changes in fire regimes as well as reductions in soil nitrate availability, resulting in a reduction of native plant communities and wildlife abundance (Grice et al. 2013). Studies have shown that invasive grasses can significantly reduce the growth of long-lived native perennial forbs utilized by wildlife species (Williams and Crone 2006). In the USA, Flanders et al. (2006) observed that overall bird and arthropod abundance, as well as forb and grass species richness, was negatively impacted in areas dominated by invasive grasses.
Invasive species result from a number of factors, including habitat loss, fragmentation, disturbances, and climate change (e.g., Carey et al. 2012; Didham et al. 2007). Changes in climate patterns, in particular, can result in substantial changes to biodiversity, species ranges, and interactions with other species and their life histories (e.g., Hellmann et al. 2008; Mainka and Howard 2010). As a result, climate change, in conjunction with other aforementioned factors, can create shifts in distribution and abundance of invasive species (Kolb et al. 2002; Hoffmeister et al. 2005). These environmental and anthropogenic factors can also cause shifts within native communities and create opportunities for native species to increase in abundance or biomass in a landscape.
Traditionally, invasives are considered non-native or exotic species. However, native species within their natural ranges can have similar effects to those of invasive species. These species are defined as native invaders (Carey et al. 2012; Shackelford et al. 2013). There is growing consensus that native plants can act invasively and pose environmental threats similar to non-native or exotic species (Head and Muir 2004; Valéry et al. 2008). While there has been research conducted native invaders in salt marsh habitats (Minchinton 2012), little research has been conducted on the concept of native plants acting as invasives in rangelands, particularly in South Texas. As a result, the potential impacts that these species may have on communities of vegetation is unknown. Much of the research conducted on native invasives eludes to the possibility that, similar to non-native invasives, they may have negative effects on other native flora and fauna. The ability to quantify the spatial dynamics of native invasives could be key for the proper management of these species and could aid in the maintenance and restoration of native vegetative communities.
Remote sensing techniques have been widely used in vegetation mapping and spatial distribution assessment (Xu et al. 2013; Young and Sanchez-Azofeifa 2004). The use of remote sensing approaches provide the ability to monitor vegetation spanning large areas, analyze spatial dynamics, and offer long-term data analysis. These techniques are useful for classifying, delineating, and quantifying the spatial distribution of vegetation (Homer et al. 1993). Everitt et al. (1995) tested the feasibility of utilizing aerial photography and videography in North Dakota and Montana to distinguish leafy spurge (Euphorbia esula L. var. esula) from other plant species. In Colorado, remote sensing techniques have been used to stratify vegetation, providing the ability to distinguish scree slopes and snow beds (Frank and Thorn 1985). Anderson et al. (1993) used satellite imagery to detect and map false broomweed (Ericameria austrotexana Johnson.) in Texas, with the greatest accuracy occurring during drought conditions. These studies show that remote sensing techniques can be a viable option when monitoring the spatial and temporal distribution of a single species across large areas. The use of remote sensing data, in conjunction with landscape metrics and spatial analysis, can be very useful in understanding the spatial and temporal dynamics of the patterns of native invasives and their related ecological processes. This can be practical in landscapes where livestock and wildlife play an important role in influencing the patterns of vegetation and ecosystem function (Derner et al. 2009; Young et al. 2014) and may aid in better understanding the patterns of establishment by native invasives.
In this study, we analyzed the encroachment of tanglehead (Heteropogon contortus (L.) P. Beauv. ex Roem. & Schult.), a native invasive species, in South Texas. Tanglehead is a perennial bunchgrass native to Texas, northern Mexico, and much of the southwestern USA. It can also be found in Hawaii, Australia, and several African and European countries (Glendening 1941; Tjelmeland and Bryant 2010). Since at least 2000, this species has been expanding across South Texas rangelands, becoming a dominant species in plant communities within this region (Bielfelt and Litt 2016). Possible causes for the expansion of tanglehead may be related to an increase of milder winters and/or changes in rainfall regimes. The encroachment of tanglehead on rangelands is a cause for concern among ranchers and landowners as it may have negative effects on rangeland diversity as well as wildlife habitat (Tjelmeland and Bryant 2010). Unfortunately, little is known about the spatial extent and distribution of tanglehead in South Texas. Field monitoring for this species can be difficult due to the large spatial extent of its distribution. Thus, remote sensing could be useful in analyzing tanglehead and its dispersal.
There have been many studies conducted using remote sensing techniques to monitor vegetation successfully, but none have been applied to tanglehead. The aim of this research is to understand the spatial and temporal dynamics of tanglehead as a model for the assessment of native invasives. The specific objectives of this research are to (1) evaluate the feasibility of classifying tanglehead using the US National Agriculture Imagery Program (NAIP) imagery (hereafter “NAIP aerial imagery”) data, (2) assess the spatial and temporal distribution of tanglehead in relation to soil type and distance from roads, and (3) quantify the temporal and spatial distribution of tanglehead on our study sites. To accomplish these objectives, we combined remote sensing data and techniques with spatial analysis and landscape metrics to describe and quantify the spatial and temporal distribution of tanglehead. We hypothesized that (1) the combination of the normalized difference vegetation index (NDVI) with red, green, blue, and near-infrared bands from aerial imagery would create a spectral signature unique to tanglehead, allowing it to be identified from other vegetation types; (2) soil texture (sandy soils) and roads (distance from roads) influenced the occurrence of tanglehead; and (3) the spatial and temporal dynamics of tanglehead dispersal could be quantified using a multi-temporal analysis.
Methods
Study area
Our study was conducted on five sites with similar vegetation communities in Jim Hogg, and Duval Counties (Fig. 1) thanks to access provided by landowners. Site 1 had an area of 2529.7 ha; site 2, 6587.7 ha; site 3, 2736.7 ha; site 4, 3757.8 ha; and site 5, 6930.5 ha. The total area studied covered 22,542 ha, which corresponds approximately to 2.5% of the Coastal Sand Sheet subregion of South Texas (Texas Parks and Wildlife Department 2016a). The Coastal Sand Sheet subregion encompasses approximately 919,800 ha spread throughout Duval, Jim Hogg, Kenedy, Kleberg, Brooks, Willacy, Hidalgo, Jim Wells, and Starr Counties, and a small portion of Zapata County. Mean annual rainfall for this area of South Texas ranges from 500 to 800 mm (from Zapata to Kenedy Counties) (Texas Parks and Wildlife Department 2016b). Soil composition within the region ranges from deep sandy and coarse sandy soils to clay loam soils which support a mosaic of vegetation types. Live oak (Quercus virginiana Mill.) is scattered throughout the region along with honey mesquite (Prosopis glandulosa Torr.). Shrubs such as granjeno (Celtis pallida Torr.), brasil (Condolia hookeri M.C. Johnst.), black brush acacia (Acacia rigidula Benth.), and catclaw acacia (A. greggii A. Gray var. greggii) are also widespread in the region. Grasslands in this region are dominated by fringed signalgrass (Urochloa ciliatissima (Buckley) R.Webster.), hooded windmillgrass (Chloris cucullata Bisch.), purple threeawn (Aristida purpurea Nutt.), Reverchon’s bristlegrass (Seteria reverchonii (Vasey) Pilg.), and seascoast bluestem (Schizachyrium scoparium Nash E.P. Bicknell), a multitude of forbs including wooly (Croton capitatus Michx.) and northern crotons (C. glandulosus L.) and sand sunflower (Helianthus debilis Nutt.), and a variety of cacti such as Texas prickly pear (Opuntia lindheimeri Salm-Dyck ex Elgelm var. lindheimeri [Engelm.] Parfitt & Pinkava) and tasajillo (O. leptocaulis [D.C.] F.M. Knuth) (Everitt et al. 2002). This subregion has experienced increasing prevalence of tanglehead since the 1990s (Texas Parks and Wildlife Department 2016a; Wester et al. 2017). The prevalence of sandy soils and high temperatures in this area provides excellent conditions conducive for the growth of tanglehead (Tothill 1977).

Location of five sites in Jim Hogg and Duval Counties in the Coastal Sand Sheet of South Texas, USA. The dark gray areas with thick boundaries represent areas covered by NAIP aerial imagery (digital orthophoto quarter quads) for each respective study site. The thin black lines outside the sites represent the county boundaries of Jim Hogg, Duval, Zapata, Brooks, and Starr counties, Texas, USA
Data collection and analyses
We used NAIP aerial imagery from the United States Department of Agriculture Farm Service Agency. This aerial imagery is periodically acquired during the agricultural growing season in the continental USA (United States Department of Agriculture 2015). We downloaded NAIP aerial imagery (1-m resolution, natural color, and color-infrared digital orthophoto quarter quads) of the study sites for the years of 2008, 2010, 2012, and 2014 through the Texas Natural Resources Information System (TNRIS) (http://tnris.org/) because these years have all the required bands (red, green, blue, and near-infrared bands) for our analysis. Available imagery from previous years is in black and white or color infrared (red, green, and near-infrared bands), which did not allow us to perform the methodology developed in this research. We developed a NDVI layer in ERDAS Imagine 2015 (Hexagon Geospatial) and then combined it with the original NAIP aerial imagery bands (red, green, blue, near infrared) using the layer stack function in ERDAS Imagine 2015 (Perotto-Baldivieso et al. 2009). We used NDVI as a measure of vegetation “greenness” to identify the spectral signature of tanglehead. Photosynthetic activity influences the NDVI values, increasing as “greenness” or photosynthetic activity increases in a living plant (Burgan and Hartford 1993). This allows vegetation to be more easily distinguishable from other cover types (Pettorelli et al. 2005). The combined images (NAIP bands + NDVI band) were classified using an unsupervised classification method (Xie et al. 2008; Perotto-Baldivieso et al. 2009). This method is based on an Iterative Self-Organizing Data Analyses Technique Algorithm (ISODATA) which recalculates the statistics of each image after each iteration until the defined number of iterations has been met and/or until a percent convergence threshold (95%) has been reached. The output image was reclassified as woody vegetation, tanglehead, non-tanglehead herbaceous, and bare ground.
Our goal was to achieve an overall image accuracy of 85% accurate or greater (Jensen 1995). This was determined by first conducting an initial desk accuracy assessment. The purpose of this assessment was to review the classification against the original NAIP aerial imagery prior to going to the field. We generated 200 random points per image in ArcMap 10.5 (ESRI; Redlands, CA) and compared visual observation and image classification for each point. We then calculated an overall accuracy assessment using a confusion matrix (Congalton 1991; Foody 2009). If the overall accuracies were < 85%, classified image(s) were reviewed and reclassified. We repeated this process until an accuracy of ≥ 85% was obtained for each individual image. Secondly, field data for each of the classified cover types were collected using 1–2 m accuracy hand-held GPS units (Trimble Juno 5). A total of 1423 field points were collected (Table 1). We collected points for each cover class following the road network and randomly selected sampling areas that were at least 800 m apart. At each sampling area, we randomly collected between 5 and 10 points in a 300 m radius from the sampling area. Once the image classification protocol met an 85% accuracy or more, we repeated the image classification process for 2008, 2010, and 2012. Once the images met accuracy standards, we used the classified imagery to assess the spatial and temporal distribution of tanglehead in relation to soil type, distance from roads, as well as quantifying the amount and spatial distribution of tanglehead in our study sites.
We derived soil data for Jim Hogg and Duval Counties from the soil survey geographic database (SSURGO) to analyze the presence of tanglehead in relation to soil textural class (Natural Resources Conservation Service 2017). We grouped soils for each county into one of 12 soil textural classes based on the soil texture triangle (National Agriculture Institute 2015). Textural classes found on the study sites were clay, clay loam, loam, loamy sand, sand, sandy clay loam, and sandy loam. We used zonal statistics in ArcGIS (ESRI; Redlands, CA) to quantify tanglehead cover within each textural class for each study site.
Roads and infrastructure were digitized at a cartographic scale of 1:1500 from the NAIP aerial imagery. We used the Euclidean distance analysis to quantify presence of tanglehead cover in relation to the distance from the roads. This spatial analysis measures the distance from the roads to the center of each pixel in the raster layer. We assessed distance from roads by grouping pixels with tanglehead in 25-m bins and calculated frequency distribution (%) for each bin and each year.
We quantified the spatial structure and temporal changes of tanglehead across the landscape using class level landscape metrics (Fragstats 4.2; McGarigal et al. 2012) for tanglehead. These metrics were percentage cover (PLAND; %), mean patch area (MPA; ha), patch density (PD; number of patches/100 ha), mean shape index (MSI), largest patch index (LPI; %), aggregation index (AI), and edge density (ED; m/ha) (Perotto-Baldivieso et al. 2011; Young et al. 2014; Zemanova et al. 2017). We used PLAND to measure the overall changes in tanglehead cover present in the landscape. PD and MPA describe the number and size of patches of tanglehead. ED quantifies the amount of edge per unit area, and LPI provides a percentage of the area covered by the largest patch of tanglehead in the landscape. We measured the aggregation or lumping of patches using AI and patch complexity with MSI. We compared landscape metrics between years using a Kruskal–Wallis one-way variance analysis with a significance level of 0.05 due to unequal variances and non-normal distributions (Dickins et al. 2013).
Results
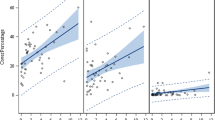
The combination of the NDVI and original bands yielded image classifications that exceeded ≥ 85% overall accuracy across all five sites: 86.1% (site 1), 85.4% (site 2), 89.5% (site 3), 88.2% (site 4), and 85.6% (site 5) (Table 1). The greatest percentages of tanglehead cover in 2014 occurred in sandy soils (mean = 20.1%; SE = 6.2) followed by loamy sand (mean = 16.3%; SE = 5.6) and sandy loam (mean = 15.3%; SE = 5.6) soils. The presence of tanglehead cover increased (Table 2) in loamy sand (X2 = 9.2, P < 0.05, Fig. 2a), sand (X2 = 10.2, P < 0.05, Fig. 2b), and sandy loam (X2 = 7.5, P < 0.10, Fig. 2c) soil textural classes between 2008 and 2014. We did not observe tanglehead cover in other textural classes across the study site. The Euclidean distance analysis values show that the greatest amounts of tanglehead cover occurred within the first 150 m from the roads (Fig. 3). Overall percent tanglehead cover increased (P < 0.01) from 7.1% (SE = 1.1) in 2008 to 17.8% (SE = 5.4) in 2014 (Fig. 4a). Values for MPA increased (P < 0.01) from 5 m2 (SE = 0.7) in 2008 to 14 m2 (SE = 6.8) in 2014 (Fig. 4b). Values of PD decreased (P < 0.01) from 2008 (mean = 15,449.4 patches/100 ha; SE = 1672.5) to 2010 (mean = 6011.3 patches/100 ha; SE = 749.4). This is probably due to small patches coalescing as tanglehead cover increases. However, PD values increased in 2012 (mean = 10,349.9 patches/100 ha; SE = 639.8) and 2014 (mean = 16,444.6 patches/100 ha; SE = 1769.7) (Fig. 4c). Values for MSI were similar across years (X2 = 7.9, P = 0.05, Fig. 4d). Largest patch index values increased from 0.1 ha (SE = 0.03) in 2008 to 0.4 ha (SE = 0.2) in 2014 (Fig. 4e). In 2008, AI values were higher than 2010 and 2012, but similar to 2014 (X2 = 12.3, P < 0.01, Fig. 4f). Values for ED decreased from 2008 (mean = 1608.5 m/ha; SE = 203.3) to 2010 (mean = 1007.6 m/ha; SE = 98.2) and then increased in 2012 (mean = 1628.0 m/ha; SE = 94.2) and 2014 (mean = 2637.7 m/ha; SE = 347.2) (Fig. 4g).

Relationship between tanglehead cover (%) and loamy sand (a), sand (b), and sandy loam (c) soil textural classes in Jim Hogg and Duval Counties in the Coastal Sand Sheet of South Texas, USA. Error bars represent standard error

Percentage of tanglehead in relation to the distance from roads between 2008 and 2014 in Jim Hogg and Duval Counties in the Coastal Sand Sheet of South Texas, USA. Error bars represent standard error

Landscape metrics describing the spatial structure of tanglehead in the five study sites in South Texas: percentage cover (a), mean patch area (b), patch density (c), mean shape index (d), largest patch index (e), aggregation index (f), and edge density (g). Error bars represent standard error
Discussion
One-meter resolution NAIP aerial imagery merged with the NDVI band proved successful in identifying tanglehead from other vegetation types. To our knowledge, this is the first study that has used this approach to monitor tanglehead over large areas. Perotto-Baldivieso et al. (2009) used similar approaches to assess small patches of depression forests in tropical environments. The accuracy (> 85% accuracy) of this methodology in both studies provides justification for expanding the use of this approach to other areas and potentially to assess other species. This technique can prove to be an effective tool for managing and studying native invasives over large areas with potential applications in wildlife and livestock management. However, it is also important to highlight that these methods can have some limitations in terms of spatial, spectral, and temporal resolution. In our study, given the speed of tanglehead expansion, the NAIP aerial imagery used was high in spatial resolution (1 m), but low in spectral (four bands: red, green, blue, and near infrared) and temporal resolution (every 2 years). The use of NDVI merged with the NAIP bands improved the spectral resolution by adding information related to vegetation “greenness” across the study area. Temporal resolution could potentially be improved with LANDSAT imagery (30 m) which is collected every 16 days (Roy et al. 2014). The use of LANDSAT for tanglehead studies still needs to be explored, as cloud cover can limit the image availability for our study area. Spatial resolution can also limit the ability to accurately identify tanglehead within our study area. The MPA values recorded for tanglehead were 0.0041 ha (41 m2), and the pixel resolution for LANDSAT is 0.09 ha (900 m2). Future research should explore the fusion of lower spatial resolution and higher spectral and temporal resolution platforms for the monitoring of tanglehead. Most available high spectral resolution sensors acquire data with a relatively coarse spatial resolution, limiting their applications in areas where vegetation appears in small patches (Zhang 2015). Image fusion combines images from different platforms with minimal loss of the original data, potentially providing a balance between temporal, spectral, and spatial resolution for image classification (Amarsaikhan et al. 2011; Zhang 2010). Additionally, the use of unmanned aerial systems could deliver very high-resolution imagery for monitoring native invasives. Unmanned aerial systems provide rapid acquisition of ultrahigh spatial resolution data at relatively low cost and can be used to sample areas and identify native invasive communities over large areas before image classification begins. This field of remote sensing could be a valuable tool in the mapping and detection of native invasives in the near future (Mafanya et al. 2017).
The use of remote sensing combined with spatial analysis and landscape metrics was the key in quantifying the spatial and temporal dynamics of tanglehead spread in the landscape between 2008 and 2014. We found that the greatest percentage of tanglehead cover occurred near roads (150 m) in soils with sandy texture. As new patches of tanglehead appeared in the landscape (increased PD), existing patches as well as the new patches coalesced, forming larger stands covering larger areas (increased PLAND, LPI, ED, MPA, and AI between 2008 and 2014). Landscape metrics were the key in identifying the spatial patterns that helped us understand the spread of tanglehead at the landscape level. Young et al. (2014) found that the use of spatial data derived from remote sensing, combined with landscape metrics, was valuable for assessing vegetation patterns and their related ecological processes.
Though specific mechanisms of dispersal were not studied, the Euclidean distance analysis results suggest that roads may play an important role in the dispersal of tanglehead. Roads are known to be a dispersal mechanism by humans, livestock, and wildlife (Wichmann et al. 2009). We hypothesize that tanglehead first establishes near roads and that vehicles collect seeds in the tires or on the body of the vehicle and disperse them over great distances (Auffret and Cousins 2013). Roads also act as a corridor utilized by livestock and wildlife, allowing them to freely move through the landscape. Livestock and wildlife can disperse seeds that are partly edible or that may be attached to them (Planchuelo et al. 2016). As a result, livestock and wildlife could be potential vectors by which native invasive species spread. Future research should emphasize on tanglehead response to changes winter temperatures and precipitation as a model to evaluate the impact of climate fluctuation to native invasive species communities.
Conclusions
Our study has shown the value of analyzing spatial and temporal dynamics of tanglehead with remote sensing techniques, spatial analysis, and landscape metrics to improve our understanding of establishment and dispersal processes of a native invasive. Our research specifically sought to validate the feasibility of classifying tanglehead and quantifying its spatial and temporal distribution in the western South Texas Sand Sheet as an example to monitor the invasion of individual species. Field monitoring of a vegetative species can be extremely expensive and time intensive and is dependent upon landowner cooperation to be successful. The ability to remotely analyze vegetation can provide opportunities to evaluate large areas at minimal cost and aid in understanding dispersal processes of native invasives. The approach taken in this study generates information about the distribution and dispersal of a native invasive species and could potentially be used to help make management decisions and develop monitoring programs to control and reduce the expansion of native invasives. The approaches from this study also have the potential to be utilized to assess and quantify the effects of native invasive species on wildlife habitat, livestock operations, and restoration strategies.
Abbreviations
- AI:
-
Aggregation index
- CIR:
-
Color infrared
- ED:
-
Edge density
- ISODATA:
-
Iterative Self-Organizing Data Analysis Technique Algorithm
- LPI:
-
Largest patch index
- MPA:
-
Mean patch area
- MSI:
-
Mean shape index
- NAIP:
-
National Agriculture Imagery Program
- NDVI:
-
Normalized difference vegetation index
- PD:
-
Patch density
- PLAND:
-
Percentage of vegetation cover
- SSURGO:
-
Soil survey geographic database
- TNRIS:
-
Texas Natural Resources Information System
References
Amarsaikhan D, Saandar M, Ganzorig M, Blotevogel HH, Egshiglen E, Gantuyal R, Nergui B, Enkhjargal D (2011) Comparison of multisource image fusion methods and land cover classification. Int J Remote Sens 33:2532–2550
Anderson GL, Everitt JH, Richardson AJ, Escobar DE (1993) Using satellite data to map false broomweed (Ericameria austrotexana) infestations on South Texas rangelands. Weed Technol 7:865–871
Auffret AG, Cousins SAO (2013) Grassland connectivity by motor vehicles and grazing livestock. Ecography 36:1150–1157
Bielfelt BJ, Litt AR (2016) Effects of Heteropogon contortus (tanglehead) on rangelands: the tangled issue of native invasive species. Rangeland Ecol Manag 69:508–512
Burgan RE, Hartford RA (1993) Monitoring vegetation greenness with satellite data. U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Ogden Ut. Gen Tech Rep INT-297. 13 P
Carey JP, Sanderson BL, Barnas KA, Olden JD (2012) Native invaders––challenges for science, management, policy, and society. Front In Ecol And The Environ 10:373–381
Congalton GR (1991) A review of assessing the accuracy of classifications of remotely sensed data. Remote Sens Environ 37:35–46
Derner JD, Lauenroth WK, Stapp P, Augustine DJ (2009) Livestock as ecosystem engineers for grassland bird habitat in the western great plains of North America. Rangeland Ecol Manag 62:111–118
Dickins EL, Yallop AR, Perotto-Baldivieso HL (2013) A multiple-scale analysis of host plant selection in Lepidoptera. J Insect Conserv 17:933–939
Didham RK, Tylianakis JM, Gemmell NJ, Tand TA, Ewers RM (2007) Interactive effects of habitat modification and species invasion on native species decline. Trends Ecol Evol 22:489–496
Everitt JH, Anderson GL, Escobar DE, Davis MR, Spencer NR, Andrascik RJ (1995) Use of remote sensing for detecting and mapping leafy spurge (Euphorbia esula). Weed Technol 9:599–609
Everitt JH, Drawe LD, Lonard IR (2002) Trees, shrubs, & cacti of South Texas. revised ed. Texas Tech University Press, Lubbock
Flanders AA, Kuvlesky WP, Ruthven DC, Zaiglin RE, Bingham RL, Fulbright TE, Hernandez F, Brennan LA (2006) Effects of invasive exotic grasses on South Texas rangeland breeding birds. Auk 123:171–182
Foody GM (2009) Sample size determination for image classification accuracy assessment and comparison. Int J Remote Sens 30:5273–5291
Frank TD, Thorn CE (1985) Stratifying alpine tundra for geomorphic studies using digitized aerial imagery. Artic and Alpine Research 17:179–187
Fulbright TE, Hickman KR, Hewitt DG (2013) Exotic grass invasion and wildlife abundance and diversity, South-Central United States. Wildl Soc B 37:503–509
Glendening GE (1941) Development of seedlings of Heteropogon contortus as related to soil moisture and competition. Botanical Gaz 102:684–698
Grice AC, Vanderduys EP, Perry JJ, Cook GD (2013) Patterns and processes of invasive grass impacts on wildlife in Australia. Wildl Society Bull 37:478–485
Head L, Muir P (2004) Nativeness, invasiveness, and nation in Australian plants. American Geogr Soc 94:199–217
Hellmann JJ, Byers JE, Bierwagen BG, Dukes JS (2008) Five potential consequences of climate change for invasive species. Conserv Biol 22:534–543
Hoffmeister TS, Vet LEM, Biere A, Holsinger K, Filser J (2005) Ecological and evolutionary consequences of biological invasion and habitat fragmentation. Ecosystems 8:657–667
Homer CG, Edwards TC, Ramsey RD, Price KP (1993) Use of remote sensing methods in modelling sage grouse winter habitat. J Wildlife Manage 57:78–84
Jensen JR (1995) Introductory digital image processing. Prentice Hall, New Jersey
Kolb A, Alpert P, Enters D, Holzapfel C (2002) Patterns of invasion within a grassland community. J Ecol 90:871–881
Mafanya M, Tsele P, Botai J, Manyama P, Swart B, Monate T (2017) Evaluating pixel and object based image classification techniques for mapping plant invasions from UAV derived aerial imagery: Harrisia pomanensis as a case study. ISPRS J Photogramm 129:1–11
Mainka SA, Howard GW (2010) Climate change and invasive species: double jeopardy. Integr Zoology 5:102–111
McGarigal K, Cushman SA, Ene E (2012) FRAGSTATS v4: spatial pattern analysis program for categorical and continuous maps. University of Massachusetts, Amherst Available at http://www.umass.edu/landeco/research/fragstats/fragstats.html
Minchinton TE (2012) Mangrove invasion of salt marsh in temperate Australia: predicting patterns of seedling recruitment under fluctuating environmental conditions. In: Dahdouh-Guebas F, Satyanarayana B (eds) proceedings of the international conference: meeting on mangrove ecology, functioning and management (MMM3), Galle, July 2012. VLIZ special publication 57, VLIZ, Brussels, p 115
National Agriculture Institute (2015) Just the facts: introduction to plant science. National Agriculture Institute, Inc., Idaho
Natural Resources Conservation Service (2017) Web soil survey. https://websoilsurvey.nrcs.usda.gov/. Accessed 11 Aug 2017
Perotto-Baldivieso HL, Meléndez-Ackerman E, García MA, Leimgruber P, Cooper SM, Martinez A, Calle P, Ramos Gonzales OM, Quiñones M, Christen C, Pons G (2009) Spatial distribution, connectivity, and the influence of scale: habitat availability for the endangered Mona Island rock iguana. Biodivers Conserv 18:905–917
Perotto-Baldivieso HL, Wu XB, Peterson MJ, Smeins FE, Silvy NJ, Schwertner TW (2011) Flooding-induced landscape changes along dendritic stream networks and implications for wildlife habitat. Landscape Urban Plan 99:115–122
Pettorelli N, Vik JO, Mysterud A, Gaillard JM, Tucker CJ, Stenseth NC (2005) Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends in Ecol and Evol 20:503–510
Planchuelo G, Catalán P, Delgado JA (2016) Gone with the wind and stream: dispersal in the invasive species Ailanthus altissima. Acta Oecol 73:31–37
Roy DP, Wulder MA, Loveland TR, Woodcock CE, Allen RG, Anderson MC, Helder D, Irons JR, Johnson DM, Kennedy R, Scambos TA, Schaaf CB, Schott JR, Sheng Y, Vermote EF, Belward AS, Bindschadler R, Cohen WB, Gao F, Hipple JD, Hostert P, Huntington J, Justice CO, Kilic A, Kovalskyy V, Lee ZP, Lymburner L, Masek JG, McCorkel J, Shuai Y, Trezza R, Vogelmann J, Wynne RH, Zhu Z (2014) Landsat-8: science and product vision for terrestrial global change research. Remote Sens Environ 145:154–172
Shackelford N, Renton M, Perring MP, Hobbs RJ (2013) Modeling disturbance-based native invasive species control and its implications for management. Ecol Appl 23:1331–1344
Texas Parks and Wildlife Department (2016a). Texas natural subregions. https://tpwd.texas.gov/gis/data/downloads. Accessed 30 Aug 2016
Texas Parks and Wildlife Department (2016b) The state of water in the South Texas Brush Country. http://tpwd.texas.gov/landwater/water/environconcerns/regions/southtexas.phtml. Accessed 11 May 2016
Tjelmeland A, Bryant FC (2010) From the field: tanglehead: understanding an emerging threat to quail habitat in South Texas. The Bobwhite Post 13:1–2
Tothill JC (1977) Seed germination studies with Heteropogon contortus. Australian J Ecol 2:477–484 https://doi.org/10.1111/j.1442-9993.1977.tb01163.x
United States Department of Agriculture (2015) NAIP imagery. https://www.fsa.usda.gov/programs-and-services/aerial-photography/imagery-programs/naip-imagery. Accessed on 7 October 2017
Valéry L, Fritz H, Lefeuvre JC, Simberloff D (2008) In search of a real definition of the biological invasion phenomenon itself. Biol Invasions 10:1345–1351
Wester DB, Grace JL, Smith F (2017) Tanglehead in South Texas. Texas Wildlife April 2017:24–25
Wichmann MC, Alexander MJ, Soons MB, Galsworthy S, Dunne L, Gould R, Fairfax C, Niggemann M, Hails RS, Bullock JM (2009) Human mediated dispersal of seeds over long distances. Proc R Soc B 276:523–532
Williams JL, Crone EE (2006) The impact of invasive grasses on the population growth of Anemone patens, a long-lived native forb. Ecol 87:3200–3208
Xie Y, Sha Z, Yu M (2008) Remote sensing imagery in vegetation mapping: a review. J Plant Ecol 1:9–23
Xu D, Guo X, Li Z, Yang X, Yin H (2013) Measuring the dead component of mixed grassland with Landsat imagery. Remote Sens Environ 142:33–43
Young D, Perotto-Baldivieso HL, Brewer T, Homer R, Santos SA (2014) Monitoring British upland ecosystems with the use of landscape structure as an indicator for state-and-transition models. Rangeland Ecol Manag 67:380–388
Young JE, Sanchez-Azofeifa GA (2004) The role of geographical information systems and optical remote sensing in monitoring boreal ecosystems. Ecol Bull 51:367–378
Zemanova MA, Perotto-Baldivieso HL, Dickins EL, Gill AB, Leonard JP, Wester DB (2017) Impact of deforestation on habitat connectivity thresholds for large carnivores in tropical forests. Ecol Proc 6:21
Zhang C (2015) Applying data fusion techniques for benthic habitat mapping and monitoring in a coral reef ecosystem. ISPRS J Photogramm 104:213–223
Zhang J (2010) Multi-source remote sensing data fusion: status and trends. Int J Image and Data Fusion 1:5–24
Acknowledgements
The authors would like to thank The Houston Livestock Show and Rodeo and The Rotary Club of Corpus Christi Harvey Weil Sportsman Conservationist Award for their financial support. We also thank the landowners who made this research possible. We would also like to thank the Rene Barrientos tuition assistance program for providing JM with financial support. Finally, we would like to thank the Caesar Kleberg Wildlife Research Institute internal reviewers A.A.T. Conkey and A.M. Foley and two anonymous reviewers who helped improve this manuscript. This manuscript is Caesar Kleberg Wildlife Research Institute publication number 18-112.
Funding
The funding for this project was provided by the Houston Livestock Show and Rodeo and The Rotary Club of Corpus Christi Harvey Weil Sportsman Conservationist Award.
Availability of data and materials
The aerial photography used for this study are available through the Texas Natural Resources Information System (TNRIS) (https://tnris.org/).
Author information
Authors and Affiliations
Contributions
JMM and HLP-B made substantial contributions to conception, design, and acquisition of data. All authors provided significant contributions to the design, analysis, and interpretation of the data; and they have been involved in drafting the manuscript and revising it critically for important intellectual content. JMM, HLP-B, FH, EDG, SR-H, JTE, MTP, and TMS have given final approval of the version to be published; they take public responsibility for appropriate portions of the content; and they have agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Mata, J.M., Perotto-Baldivieso, H.L., Hernández, F. et al. Quantifying the spatial and temporal distribution of tanglehead (Heteropogon contortus) on South Texas rangelands. Ecol Process 7, 2 (2018). https://doi.org/10.1186/s13717-018-0113-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13717-018-0113-0