
Abstract
In this study, we propose two techniques for clustering genetic regulatory networks with mixed delays. Laws for identifying coupling parameters are designed, and the Lyapunov theorem and Lipschitz condition are employed to achieve the desired stability and to identify the coupling parameters. Using our proposed methods, the value of a cluster or the sum of clusters can approach the expected values. Numerical examples verify the feasibility and effectiveness of this method.
Similar content being viewed by others


1 Introduction
After the double helix structure of DNA was discovered by Watson and Crick in 1953, we entered a new era of molecular biology, and many subsequent studies have shown that physiological functions are controlled by biological networks instead of several molecules and genes. Biological networks include metabolic networks, protein interaction networks, and genetic regulatory networks (GRNs), where the latter are significant for studies of the interactions between mRNA and proteins at the molecular level. GRNs have been elucidated increasingly in the last 30 years due to great progress in genome sequencing and gene recognition, thereby attracting considerable attention from researchers in the areas of biology, computer science, physics, and mathematics; and thus GRNs is a focus of interdisciplinary research. Various models have been proposed to describe the transcription and translation of DNA, e.g., Boolean networks [1–3] and differential equation models [4–12], where the latter can depict the continuous dynamic behavior of mRNA and protein.
Time delays are ubiquitous in biology [13], physics [14], chemistry [15], optics [16], and complex networks [17, 18]. Thus, time delays need to be considered in GRNs because of the finite speeds of the slow processes of transcription, translation, and translocation [7–10, 12]. Furthermore, inappropriate considerations of time delays can lead to incorrect predictions of the behavior of GRNs.
Stability is essential for the design or control of GRNs because it ensures that an organism can robustly regulate its functions even if the state of the organism moves away from the equilibrium points. Zhang et al. used the improved integral inequality to conduct stability analysis for a GRN with interval time-varying delays [19]. Wang et al. performed exponential convergence analysis for an uncertain GRN with time-varying delays [20]. Zhang et al. established globally asymptotic stability criteria for a GRN with time-varying discrete and unbounded distributed delays [21]. Liu and Wu analyzed the global asymptotical stability of a GRN with time-varying delays via a convex combination method [22].
These previous studies considered the stability of GRNs but not clustering phenomena. In general, cells can communicate with neighboring cells via quorum sensing because the genetic signals transmitted from genes can support different physiological functions [23]. Therefore, clustering is a common phenomenon in interacting populations, and clustering is an important aspect of biological control [24]. Recently, some studies have investigated the clustering of GRNs, such as how the influence of coupling or noise can generate cluster patterns in an ensemble of cellular oscillators [25], how individual GRNs are divided into different clusters by cluster synchronization [23, 24, 26], and how temporal control is introduced to cluster mammalian signaling modules [27]. In this study, we investigate the desired clustering of GRNs where clusters of GRNs could have different features according to two proposed strategies. In particular, we impose no limitations on the number of nodes in each cluster, the division of the clusters of GRNs, and the time delay for feedback regulation. Comparing with the previous research of GRNs, we not only analyze the stability of GRNs, but also make the concentrations of protein and mRNA approach the desired values through our tactics. Linear matrix inequality (LMI) method is often used to obtain the stability criteria for GRNs, but the computation of LMI is complex. In this paper, stability criterion is given by a Lyapunov function.
The real genetic regulatory network is over complex, and the number of nodes is huge. Since “network motifs” [28, 29] were proposed by Alon et al. in 2002, few node genetic regulatory networks have become research hotspots. It is hoped that the real genetic regulatory network can be understood gradually by the research of such network motifs. In 2000, Elowitz et al. used three transcriptional repressor systems to build a synthetic oscillating network in Escherichia coli [30]. Gardner et al. used two transcriptional repressors to construct a synthetic toggle switch genetic regulatory network in Escherichia coli also in 2000 [31]. Therefore, 7 or 20 genes with clustering as numerical examples in Sect. 4 are significant.
2 Model of GRNs
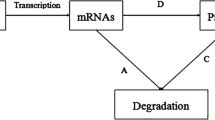
GRNs with mixed delays [12] are described by Eq. (1):
where \(m ( t ) =[m_{1} ( t ) \cdots m_{n} (t)]\in R^{n}\) and \(p ( t ) =[p_{1} ( t ) \cdots p_{n} (t)]\in R^{n}\) are the concentrations of mRNA and protein for node i at the time t, respectively, the parameters \(a_{i}\) and \(\mathrm{c}_{i}\) are the degradation rates of the mRNA and protein, and \(d_{i}\) is the translation rate. The feedback regulation delays \(\tau ( t )\), \(\sigma(t)\) are both positive, and \(f_{i} (x)\) is the Hill form regulatory function, which represents the feedback regulation of the protein on transcription, and it is described by Eq. (2):
where \(H_{i}\) is the Hill coefficient, \(v_{j}\) is a positive constant, and we propose that \(f_{j} ( x )\) is positive. \(l_{i} (t)= \sum_{j\in I_{i}} \alpha_{ij} (t)\) and \(I_{i}\) is the set of all j, which is a repressor of gene i. The coupling matrix \(W = \omega_{ij} \in R^{n\times n}\) is described as follows:
As shown in Fig. 1, we assume that there are M clusters in the GRNs (1) (\(M\geq1\)), which are denoted by \(C_{1}, C_{2}, \ldots, C_{m}\), where \(C_{k} =\{ q_{k-1} +1, q_{k-1} +2,\ldots, q_{k} \}\), with \(k=1,2,\ldots,m, q_{0} =0\), and \(q_{m} =N\).

Cluster of GRNs where \(q_{k} - q_{k-1}\) represents the number of nodes in the kth cluster and \(\{ q_{k-1} +1,\ldots, q_{k-1} + \gamma,\ldots, q_{k} \}\) represents the index set of all the nodes in the kth cluster. We find that \(m_{i} = m_{q_{k-1} +\gamma}\), when the index of the ith gene is the γth gene in the kth cluster
Let \(\overline{m}_{k} ( t ) =( m_{q_{k-1} +1}, m_{q_{k-1} +2},\ldots, m_{q_{k}} ) \), \(\overline{p}_{k} ( t ) =( p_{q_{k-1} +1}, p_{q_{k-1} +2},\ldots, p_{q_{k}})\).
Definition 1
\(X(t) \in R^{n}\) will approach the desired values \(\hat{X} \in R^{n}\) if \(\lim_{t\rightarrow\infty} \vert X(t )- \hat{X} \vert =0\).
3 Desired clustering of GRNs
In this section, the GRNs (1) are clustered using two techniques.
3.1 Method 1
In this method, the concentrations of mRNA and protein within a cluster have the same desired values \(\hat{{m}}_{{k}}\) and \(\hat{{p}}_{{k}}\), and the desired values of the nodes vary in different clusters.
Theorem 1
The desired clustering of GRNs with mixed delays will be achieved when the law for identifying the coupling parameters \(\omega_{ij}\), \(a_{i}\), \(c_{i} \beta_{i}\) (\(i \in \{ q_{k-1} +1, q_{k-1} +2,\ldots, q_{k} \}, j \in \{ 1,\ldots,N \} \)) and \(\hat{{p}}_{{k}}\) is taken as follows, respectively:
where the real numbers δ and \(\varepsilon_{i} \) are positive; the estimated identification of the uncertain adjustment parameters \(\omega_{ij}\) is \(\hat{{\omega}}_{{ij}}\) and α is an adjustment parameter; \({\theta}_{{k}}= d_{k} / c_{k}\) is a scale factor that represents the proportional relationship between \(\hat{{p}}_{{k}}\) and \(\hat{{m}}_{{k}}\), \(c_{k} = c_{q_{k-1} +1} =\cdots= c_{{q}_{{k}}}\), and \(d_{k} = d_{q_{k-1} +1} =\cdots= d_{{q}_{{k}}}\).
Proof
For \(i \in C_{k}\), the error between \(m_{i}\) and \(\hat{{m}}_{{k}}\) and the error between \(p_{i}\) and \(p_{i} ( t-\sigma ( t ) )\) are defined by Eq. (9).
We establish the Lyapunov function as follows.
The derivative form of \(V(t)\) can be described as follows:
where
For the real number \(\varepsilon_{i} > 0\), the following relationship is obtained using the Lipschitz condition:
and Eq. (10) can be simplified as follows:
The following inequation is obtained while Eq. (7) is substituted into Eq. (15):
Clearly, when (5), (6), (8) are satisfied, we find that \(\lim_{t\rightarrow\infty} \dot{V} ( t ) \leq0\). According to stability theory and Definition 1, \(m_{i} ( t ) \in \overline{m}_{k} ( t )\) and \(p_{i} ( t ) \in \overline{p}_{k} ( t )\) will approach the desired values, where \(\hat{{p}}_{{k}}= \lim_{{t\rightarrow\infty}} {\theta}_{{k}} m_{i} ( t-\sigma ( t ) )= \theta_{{k}} \hat{m}_{k}\ ( i \in \{ q_{k-1} +1, q_{k-1} +2,\ldots, q_{k} \} )\). □
3.2 Method 2
In this method, the GRNs is clustered by summing, which means that the sums of \(\overline{m}_{k} (t)\) and \(\overline{p}_{k} (t)\) have the desired values \(\hat{{S}}_{{1k}}\) and \(\hat{{S}}_{{2k}}\), respectively, and the desired values of the sum of the various clusters are different. The sums of the mRNAs and proteins in the kth cluster are defined as follows:
The error between \(S_{1k} ( t )\) and \(\hat{S}_{1k}\) and the error between \(S_{2k} ({t} )\) and \(\hat{{S}}_{{2k}} \) are defined as follows:
Theorem 2
The desired cluster of GRNs with mixed delays will be achieved when the law for identifying the coupling parameters \(\omega_{ij}\), \(a_{i}\), \(c_{i}\), \(\beta_{i}\) \(( i \in \{ q_{k-1} +1, q_{k-1} +2,\ldots, q_{k} \},j \in \{ 1,\ldots,N \} )\), and \(\hat{{S}}_{{2k}}\) is taken as follows, respectively:
where the real numbers ξ and \(\mu_{k} \) are positive; \({\vartheta}_{{k}}= d_{k} / c_{k}\) is a scale factor that represents the proportional relationship between \(\hat{{S}}_{{2k}}\) and \(\hat{S}_{1k}\); \(a_{k} = a_{q_{k-1} +1} =\cdots= a_{{q}_{{k}}}\), \(c_{k} = c_{q_{k-1} +1} =\cdots= c_{{q}_{{k}}}\), and \(d_{k} = d_{q_{k-1} +1} =\cdots= d_{{q}_{{k}}}\).
Proof
The Lyapunov function is established as follows:
The derivative form of \(V(t)\) can be described as follows:
When \(a_{k} = a_{q_{k-1} +1} =\cdots= a_{{q}_{{k}}}\), \(c_{k} = c_{q_{k-1} +1} =\cdots= c_{{q}_{{k}}}\), and \(d_{k} = d_{q_{k-1} +1} =\cdots= d_{{q}_{{k}}}\), Eq. (27) is simplified as follows:
where
For the real number \(\mu_{k} > 0\), the following relationship can be obtained using the Lipschitz condition:
and Eq. (28) can be simplified as follows:
The following inequation is obtained while Eq. (24) is substituted into Eq. (32):
Based on (22), (23), (25), one can deduce \(\lim_{t\rightarrow\infty} \dot{V} ( t ) \leq0\). According to stability theory and Definition 1, \({S}_{{1k}}{(t)}\) and \({S}_{{2k}}{(t)}\) will approach the desired values, where \(\hat{{S}}_{{2k}}= \lim_{{t\rightarrow\infty}} {\vartheta}_{{k}} S_{1k} ( t-\sigma ( t ) )= \vartheta_{{k}} \hat{S}_{1k}\). □
4 Numerical simulation
Example 1
In the numerical simulation of Example 1, the number of the nodes is fixed as \(n =7\); the parameters are fixed as \(a_{i} =2\) (\(i =1,\ldots,7\)), \(c_{k} =1\), and \(d_{k} = c_{k} \hat{p}_{k} / \hat{m}_{k} \ ( k =1,\ldots, M)\); and the adjustment parameter \({\alpha=0.5}\).
We assume that there are two clusters in the GRNs, \(M =2\), where
The temporal evolution of \(m_{i} \ ( i =1,\ldots,7)\) and \(p_{i}\) is shown in Fig. 2. Figure 2 clearly indicates that the concentrations of \(m_{i}\) and \(p_{i} \) at equilibrium approach the desired values. In addition, \(m_{i}, p_{i}\ ( i =1,2,3)\) belong to the first cluster and the concentration of \(m_{i}\), \(p_{i} \ ( i =4,5,6,7)\) belongs to the second cluster.

Time trajectories of \(m_{i} (t)\) and \(p_{i} (t)\), (\(i =1,\ldots,7\)), and identification process for the uncertain parameter \(\omega_{1j} \ ( j =1,\ldots,7)\)
According to Figs. 2 and 3, the curves for identifying \(\omega_{ij} \ (i=1,2,\ldots,7;j=1,2,\ldots,7)\) gradually tend toward the fixed values as follows:
where Ŵ is the identification estimates for the uncertain coupling matrix W;

Identification process for the uncertain parameters \(\omega_{ij} \ ( i =2,\ldots,7; j =1,\ldots,7)\)
Clearly, the value of \(\beta_{i}\), (\(i =1,\ldots,7\)) is positive, and thus inequation (7) is satisfied.
Furthermore, when we expand the number of nodes of GRNs to 20 and assume that there are three clusters in the GRNs, where \(a_{i} =3\ ( i =1,\ldots,20)\), \(c_{k} =1\), \(d_{k} = c_{k} \hat{p}_{k} / \hat{m}_{k} \ ( k =1,\ldots, M ) \); \(M =3\); \(\alpha=0.3\);
The temporal evolution of \(m_{i} \ ( i =1,\ldots,7)\) and \(p_{i}\) is shown in Fig. 4. According to Fig. 4, \(m_{i} (t)\) and \(p_{i} (t)\) reach equilibrium at the desired values and the concentrations of \(m_{i}\) and \(p_{i} \ ( i =1,\ldots,10)\) belong to the first cluster, the concentrations of \(m_{i}\) and \(p_{i} \ ( i =11,\ldots,15)\) belong to the second cluster, and the concentrations of \(m_{i}, p_{i} \ ( i =16,\ldots,20)\) approach the third cluster.

State trajectories of \(m_{i} (t)\) and \(p_{i} (t) \ ( i =1,2,\ldots,20)\)
According to Fig. 5, the curves for identifying \(\omega_{ij} \ (i=1,2,\ldots,20;j=1,2,\ldots,20)\) gradually tend to the fixed values. Based on Fig. 6, inequation (7) is satisfied.

Identification process for adjustment parameter \(\omega_{ij}\), (\(i =1,\ldots,20; j =1,\ldots,20\))

Value of \(\beta_{i}\), (\(i =1,\ldots,20\))
Example 2
In the numerical simulation of Example 2, the number of the nodes is fixed as \(n =7\), and it is assumed that there are two clusters in the GRNs, where \(a_{i} =2\ ( i =1,\ldots,7)\), \(c_{k} =2\), \(d_{k} = c_{k} \hat{S}_{2k} / \hat{S}_{1k} \ ( k =1,\ldots, M )\); \(M =2\); \(\alpha=0.5\).
where \(S_{11} = m_{1} ( t ) + m_{2} ( t ) + m_{3} (t)\) and \(S_{12} = m_{4}(t)+ m_{5} ( t ) + m_{6} ( t ) + m_{7} (t)\); \(S_{21} = p_{1} ( t ) + p_{2} ( t ) + p_{3} (t)\) and \(S_{22} = p_{4}(t)+ p_{5} ( t ) + p_{6} ( t ) + p_{7} (t)\) and the desired values of \(S_{11}\), \(S_{12}\), \(S_{21}\), \(S_{22}\) are fixed at \(\hat{S}_{11} =30\), \(\hat{S}_{12} =10\), \(\hat{S}_{21} =15\), \(\hat{S}_{22} =5\), respectively.
The temporal evolution of \(m_{i} \ ( i =1,\ldots,7)\) and \(p_{i}\) are shown in Fig. 7, respectively. Figure 8(c) clearly indicates that the values of \(S_{1k}\) and \(S_{2k}\ ( k =1,2 ) \) approach those of \(\hat{S}_{1k}\) and \(\hat{S}_{2k}\). It is important to note that the GRNs are divided into two clusters, where the concentrations of \(m_{i}\), \(p_{i} \ ( i =1,2,3)\) belong to the first cluster and the concentrations of \(m_{i}\), \(p_{i} \ ( i =4,5,6,7)\) belong to the second cluster.

State trajectory of \(m_{i} (t) \ ( i =1,2,\ldots,7)\), and identification process for the uncertain parameter \(\omega_{1j} \ ( j =1,\ldots,7)\)

(a), (b): Identification process for the adjustment parameter \(\omega_{ij} \ ( i, j =2,\ldots,7)\); (c): state trajectories of \(S_{1k} (t)\) and \(S_{2k} (t) \ ( k =1,2)\)
According to Figs. 7 and 8(a)–(b), the curves for identifying \(\omega_{ij} \ (i=1,2,\ldots,7;j=1,2,\ldots,7)\) gradually tend toward the fixed values as follows:
where Ŵ is the identification estimates for the uncertain coupling matrix W.
Clearly, the value of \(\beta_{k}\) (\(k =1,2\)) is positive, and thus inequation (24) is satisfied.
Furthermore, when we expand the number of nodes of GRNs to 20 and assume that there are three clusters in the GRNs, where \(a_{i} =3\ ( i =1,\ldots,20)\), \(c_{k} =0.5\), \(d_{k} = c_{k} \hat{S}_{2k} / \hat{S}_{1k} \ ( k =1,\ldots, M )\); \(M =3\); \({\alpha=0.2}\),
such that \(S_{11} = \sum_{i=1}^{10} m_{i} ( t ), S_{12} = \sum_{i=11}^{15} m_{i} ( t ), S_{13} = \sum_{i=16}^{20} m_{i} ( t ); S_{21} = \sum_{i=1}^{10} p_{i} ( t ), S_{22} = \sum_{i=11}^{15} p_{i} ( t ), S_{23} = \sum_{i=16}^{20} p_{i} ( t )\).
The desired values of \(S_{11}\), \(S_{12}\), \(S_{13}\), \(S_{21}\), \(S_{22}\), \(S_{23}\) are fixed at \(\hat{S}_{11} =15\), \(\hat{S}_{12} =20\), \(\hat{S}_{13} =30\), \(\hat{S}_{21} =30\), \(\hat{S}_{22} =40\), \(\hat{S}_{23} =60\), respectively.
The temporal evolution of \(m_{i} \) and \(p_{i}\) (\(i =1,\ldots,20\)) is shown in Fig. 9. According to Fig. 10, the values of \(S_{1k}\) and \(S_{2k} \ ( k =1,2,3)\) clearly approach those of \(\hat{S}_{1k}\) and \(\hat{S}_{2k}\). It is important to note that the GRNs are divided into three clusters, where the concentrations of \(m_{i}\), \(p_{i} \ ( i =1,\ldots,10)\) belong to the first cluster, the concentrations of \(m_{i}\), \(p_{i} \ ( i =11,\ldots,15)\) belong to the second cluster, and the concentrations of \(m_{i}\), \(p_{i} \ ( i =16,\ldots,20)\) belong to the third cluster.

State trajectories of \(m_{i} (t)\) and \(p_{i} (t) \ ( i =1,2,\ldots,20)\)

State trajectories of \(S_{1k} (t)\) and \(S_{2k} (t) \ ( k =1,2,3)\)
According to Fig. 11, the curves for identifying \(\omega_{ij} \ (i=1,2,\ldots,20;j=1,2,\ldots,20)\) gradually tend toward the fixed values.

Identification process for the uncertain parameter \(\omega_{ij}\), (\(i =1,\ldots,20; j =1,\ldots,20\))
Clearly, the value of \(\beta_{k}\) (\(k =1,2,3\)) is positive, and thus inequation (24) is satisfied.
5 Conclusions
In this study, we investigated desired clusters of GRNs with fixed time lags based on the Lyapunov theorem and Lipschitz condition. GRNs can be clustered using the two proposed strategies. In the first method, the concentrations of mRNAs in the same cluster approach the unique desired value. In the second method, the sums of clusters tend toward the desired value. Numerical examples were provided to demonstrate the effectiveness of the proposed clustering techniques. The results showed that the number of possible clusters in the GRNs does not affect the targeted stability of the cluster. In addition, not limiting the number of nodes in each cluster enhances the practicality and flexibility of these methods.
References
Chen, H.W., Sun, L.J., Liu, Y.: Partial stability and stabilization of Boolean networks. Int. J. Syst. Sci. 47, 2119–2127 (2016)
Paroni, A., Graudenzi, A., Caravagna, G., Damiani, C., Mauri, G., Antoniotti, M.: Cabernet: a Cytoscape app for augmented Boolean models of gene regulatory networks. BMC Bioinform. 17, 64 (2016)
Zheng, Q.B., Shen, L.Z., Shang, X.Q., Liu, W.B.: Detecting small attractors of large Boolean networks by function-reduction-based strategy. IET Syst. Biol. 10, 49–56 (2016)
Liu, R.J., Lu, J.Q., Lou, J.G., Aladi, A., Aladi, F.E.: Set stabilization of Boolean networks under pinning control strategy. Neurocomputing 260, 142–148 (2017)
Wang, J.N., Guo, B.H., Wei, W., Mi, Z.L., Yin, Z.Q., Zheng, Z.M.: The stability of Boolean network with transmission sensitivity. Phys. A, Stat. Mech. Appl. 481, 70–78 (2017)
Baroness, A., Jimenez, E., Lacerta-Garcia, M.A., Almagro, F.J., Pena, B.: Parameter inference of general nonlinear dynamical models of gene regulatory networks from small and noisy time series. Neurocomputing 175, 555–563 (2016)
Koo, J.H., Ji, D.H., Won, S.C., Park, J.H.: An improved robust delay-dependent stability criterion for genetic regulatory networks with interval time delays. Commun. Nonlinear Sci. Numer. Simul. 17, 3399–3405 (2012)
Li, L., Yang, Y.Q.: On sampled-data control for stabilization of genetic regulatory networks with leakage delays. Neurocomputing 149, 1225–1231 (2015)
Zou, C.Y., Wei, X.P., Zhang, Q., Zhou, C.: Passivity of reaction–diffusion genetic regulatory networks with time-varying delays. Neural Process. Lett. 46, 1–18 (2017)
Yu, T.T., Liu, J.X., Zeng, Y., Zhang, X., Zeng, Q.S., Wu, L.G.: Stability analysis of genetic regulatory networks with switching parameters and time delays. IEEE Trans. Neural Netw. Learn. Syst. (2017) https://doi.org/10.1109/TNNLS.2016.2636185
Lu, L., He, B., Man, C.T., Wang, S.: Robust state estimation for Markov jump genetic regulatory networks based on passivity theory. Complexity 21, 214–223 (2016)
Wang, Y., Wang, Z.D., Liang, J.L.: On robust stability of stochastic genetic regulatory networks with time delays: a delay fractioning approach. IEEE Trans. Syst. Man Cybern., Part B, Cybern. 40, 729–740 (2010)
Li, Y., Ma, W.B., Xiao, L.M., Yang, W.T.: Global stability analysis of the equilibrium of an improved time-delayed dynamic model to describe the development of T cells in the thymus. Filomat 31, 347–361 (2017)
Xia, W., Kang, J.S.: Stability of LCL-filtered grid-connected inverters with capacitor current feedback active damping considering controller time delays. J. Mod. Pow. Syst. Clean Energy 5, 584–598 (2017)
Jia, X.L., Chen, X.K., Xu, S.Y., Zhang, B.Y., Zhang, Z.Q.: Adaptive output feedback control of nonlinear time-delay systems with application to chemical reactor systems. IEEE Trans. Ind. Electron. 64, 4792–4799 (2017)
Ji, S., Hong, Y.H.: Effect of bias current on complexity and time delay signature of chaos in semiconductor laser with time-delayed optical feedback. IEEE J. Sel. Top. Quantum Electron. 23, 1800706 (2017)
Tu, Z.W., Ding, N., Li, L.L., Feng, Y.M., Zou, L.M., Zhang, W.: Adaptive synchronization of memristive neural networks with time-varying delays and reaction-diffusion term. Appl. Math. Comput. 311, 118–128 (2017)
Geng, J., Liu, M., Zhang, Y.Q.: Stability of a stochastic one-predator-two-prey population model with time delays. Commun. Nonlinear Sci. Numer. Simul. 53, 65–82 (2017)
Zhang, X., Wu, L., Cui, S.: An improved integral inequality to stability analysis of genetic regulatory networks with interval time-varying delays. IEEE/ACM Trans. Comput. Biol. Bioinform. 12, 398–409 (2015)
Wang, W., Nguang, S.K., Zhong, S., Liu, F.: Exponential convergence analysis of uncertain genetic regulatory networks with time-varying delays. ISA Trans. 53, 1544–1553 (2014)
Zhang, X., Han, Y., Wu, L., Zou, J.: M-matrix-based globally asymptotic stability criteria for genetic regulatory networks with time-varying discrete and unbounded distributed delays. Neurocomputing 174, 1060–1069 (2016)
Liu, Y., Wu, H.: Analysis on global asymptotical stability of genetic regulatory networks with time-varying delays via convex combination method. Math. Probl. Eng. 2015, Article ID 303918 (2015)
Guan, Z.H., Yue, D.D., Hu, B., Li, T., Liu, F.: Cluster synchronization of coupled genetic regulatory networks with delays via aperiodically adaptive intermittent control. IEEE Trans. Nanobiosci. 16, 585–599 (2017)
Yue, D.D., Guan, Z.H., Li, T., Liao, R.Q., Liu, F.: Event-based cluster synchronization of coupled genetic regulatory networks. Phys. A, Stat. Mech. Appl. 482, 649–665 (2017)
Zhou, T.S., Zhang, J.J., Yuan, Z.J., Chen, L.N.: Synchronization of genetic oscillators. Chaos 18, 037126 (2008)
Zhang, J.J., Yuan, Z.J., Zhou, T.S.: Synchronization and clustering of synthetic genetic networks: a role for cis-regulatory modules. Phys. Rev. E 79, 041903 (2009)
Werner, S.L., Barken, D., Hoffmann, A.: Stimulus specificity of gene expression programs determined by temporal control of IKK activity. Science 309, 1857–1861 (2005)
Milo, R., Shen-Orr, S., Itzkovitz, S., Kashtan, N., Chklovskii, D., Alon, U.: Network motifs: simple building blocks of complex networks. Science 298, 824–827 (2002)
Shen-Orr, S., Milo, R., Mangan, S., Alon, U.: Network motifs in the transcriptional regulation network of Escherichia coli. Nat. Genet. 31, 64–68 (2002)
Elowitz, M.B., Leibler, S.: A synthetic oscillatory network of transcriptional regulators. Nature 403, 335–338 (2000)
Gardner, T.S., Cantor, C.R., Collins, J.J.: Construction of a genetic toggle switch in Escherichia coli. Nature 403, 339–342 (2000)
Acknowledgements
This study was supported by the National Natural Science Foundation of China (Nos. 61425002, 61751203, 61772100 and 61672124) and by the Program for Changjiang Scholars and Innovative Research Team in University (No. IRT_15R07).
Author information
Authors and Affiliations
Contributions
All authors contributed equally to the writing of this paper. All of the authors read and approved the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Zou, C., Wei, X., Zhang, Q. et al. Desired clustering of genetic regulatory networks with mixed delays. Adv Differ Equ 2018, 150 (2018). https://doi.org/10.1186/s13662-018-1534-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-018-1534-x