
Abstract
Genes responsible for the anaerobic catabolism of benzoate in the thermophilic archaeon Ferroglobus placidus were expressed in the thermophilic lignocellulose-degrading bacterium Caldicellulosiruptor bescii, as a first step to engineering this bacterium to degrade this lignin metabolite. The benzoyl-CoA ligase gene was expressed individually, and in combination with benzoyl-CoA reductase and a putative benzoate transporter. This effort also assessed heterologous expression from a synthetically designed operon whereby each coding sequence was proceeded by a unique C. bescii ribosome binding site sequence. The F. placidicus benzoyl-CoA ligase gene was expressed in C. bescii to produce a full-length protein with catalytic activity. A synthetic 6-gene operon encoding three enzymes involved in benzoate degradation was also successfully expressed in C. bescii as determined by RNA analysis, though the protein products of only four of the genes were detected. The discord between the mRNA and protein measurements, especially considering the two genes lacking apparent protein abundance, suggests variable effectiveness of the ribosome binding site sequences utilized in this synthetic operon. The engineered strains did not degrade benzoate. Although the heterologously expressed gene encoding benzoyl-CoA ligase yielded a protein that was catalytically active in vitro, expression in C. bescii of six benzoate catabolism-related genes combined in a synthetic operon yielded mixed results. More effective expression and in vivo activity might be brought about by validating and using different ribosome binding sites and different promoters. Expressing additional pathway components may alleviate any pathway inhibition and enhance benzoyl-CoA reductase activity.
Similar content being viewed by others
Introduction
Consolidated bioprocessing (CBP) is projected to be an inexpensive route to renewably produce bioethanol from lignocellulosic biomass (Lynd et al. 2005, 2008). Caldicellulosiruptor bescii has emerged as a promising candidate microbial biocatalyst to achieve this (Chung et al. 2014), and it is actively being engineered to produce a more effective consolidated bioprocessing host organism. It is well suited for this, as it natively contains the ability to solubilize lignocellulosic biomass, using its array of secreted CAZy enzymes (Blumer-Schuette et al. 2014) and novel biomass digestion strategy, employing large, thermostable cellulolytic enzymes that contain multiple catalytic domains (Brunecky et al. 2013; Blumer-Schuette et al. 2014). C. bescii is an anaerobic thermophile that grows optimally at 78 °C and ferments a wide variety of carbohydrates, including those it solubilizes from lignocellulosic biomass (Hamilton-Brehm et al. 2010). While C. bescii can also solubilize lignin components (Kataeva et al. 2013), it cannot catabolize the resulting soluble constituents and decomposition products. The successful engineering of lignin monomer catabolism in C. bescii (or other such CBP biocatalysts) would substantially increase whole-biomass conversion in a CBP process and improve strain resistance to inhibitors released from biomass.
Organisms that catabolize lignin monomers and other aromatic species initially modify these molecules using specialized pathways with narrow and non-overlapping specificity. These intermediates are transformed, or ‘funneled’ to one of a few common metabolic intermediates, including benzoyl-Coenzyme A in anaerobic microorganisms (Harwood et al. 1998). These common intermediate metabolites then enter subsequent pathways where the aromatic ring portions are dearomatized and cleaved in energy-intensive reactions. Intermediates are further metabolized to acetyl-CoA in a pathway which resembles the β-oxidation pathway for fatty acids (Carmona et al. 2009). For every molecule of benzoyl-CoA processed, two molecules of acetyl-CoA are produced, which can then be utilized in other metabolic reactions. In anaerobic organisms, dearomatization and subsequent degradation reactions are carried out on aromatic molecules activated with CoA moieties through a thioester bond. This CoA-thioester reduces the resonance in aromatic moieties by drawing electrons away from otherwise stable resonance structures. This ultimately reduces the energetic demand of aromatic ring reduction (Fuchs et al. 2011).
One such organism able to catabolize aromatic molecules is the thermophilic archaeon Ferroglobus placidus, which grows optimally at 85 °C. It is able to grow on benzoate using Fe(III) as a terminal electron acceptor (Holmes et al. 2011; Tor and Lovley 2001). F. placidus synthesizes benzoyl-CoA as a metabolic intermediate when catabolizing several different aromatic chemicals (Holmes et al. 2012; Schmid et al. 2016). It utilizes an ATP-dependent benzoyl-CoA ligase (BCL) (Schmid et al. 2016) to add a CoA moiety to benzoate, and a type I benzoyl-CoA reductase (BCR) (Boll et al. 2014; Smith et al. 2015; Schmid et al. 2016) to enable the first, and most energetically demanding, reactions to reduce and break the aromaticity of benzoyl-CoA. Both enzymes are ATP dependent, coupling the hydrolysis of ATP to what are otherwise thermodynamically unfavorable reactions (Boll et al. 2014; Schmid et al. 2015).
Other organisms not able to natively catabolize aromatic species have been successfully engineered to do so. E. coli, heterologously expressing eight catabolic genes and one transport gene form Pseudomonas putida, gained the ability to grow on protocatechuate and 4-hydroxybenzoate (Clarkson et al. 2017), two common ‘funneling’ intermediates in aerobic lignin monomer catabolism. Similarly, a strain of Azoarcus CI B made deficient in its otherwise native ability to catabolize benzoate, was phenotypically rescued by heterologous expression of a complete benzoate degradation pathway (Zamarro et al. 2017).
Heterologous expression of genes in C. bescii generally comprises the expression of one or two genes simultaneously (e.g. a catabolic enzyme and a selection marker), each expressed from a separate promoter (Yang et al. 2009; Chung et al. 2014; Chung et al. 2015a, b, c, d). In addition to taking initial steps toward enabling lignin monomer and aromatic catabolism, another primary objective of this work was to assess C. bescii heterologous expression of multiple genes from a single transcript, employing ribosome binding sites of the host upstream of each gene to maximize protein expression and confer much more complex biological functions. Efforts herein constitute the first steps toward expression in C. bescii of the full aromatic catabolism pathway(s) in F. placidus linking aromatic catabolism to acetyl-CoA production.
Materials and methods
Construction of C. bescii heterologous expression strains
Heterologous expression strains were constructed in C. bescii JWCB018. C. bescii JWCB018 is a mutant derivative of the C. bescii wildtype strain C. bescii DSMZ 6725 that has been engineered for uracil auxotrophy and deficiency in M. CbeI restriction/methylation system (Chung et al. 2013a, b). C. bescii JWCB018 was a generous gift from, and is also available upon request from the laboratory of Janet Westpheling of the Department of Genetics at the University of Georgia, and the base wildtype strain C. bescii DSMZ 6725 is available through DSMZ. Two expression plasmids were constructed and transformed into C. bescii strain JWCB018 (Additional file 1: Figures S1 and S2, Tables S2 and S3). Plasmids were constructed using plasmid pJGW07 as the vector backbone (Chung et al. 2013a, b). Insert components were generated and cloned from synthesized dsDNA fragments (Integrated DNA Technologies, Inc. Coralville, IA), or amplifications products thereof. The codon-optimized gene sequences have been deposited into GenBank and can be retrieved at the following accession numbers: Ferp_1370_codon_optimized; MK294010, Ferp_1044_codon_optimized; MK294011, Ferp_1184_codon_optimized; MK294012, Ferp_1185_codon_optimized; MK294013, Ferp_1186_codon_optimized; MK294014, Ferp_1187_codon_optimized; MK294015. Plasmid pJOT1 expressed a codon-optimized, his-tagged copy of the Ferp_1044 gene, encoding for a Benzoyl-CoA Ligase (Holmes et al. 2012; Schmid et al. 2015) (Additional file 1: Figure S1), using a previously demonstrated C. bescii promoter natively found upstream of an S-layer protein (Athe_2303). This plasmid (Additional file 1: Table S2) was transformed into strain JWCB018 (Chung et al. 2013a, b) to generate strain JWCB018 pJOT1 (Additional file 1: Table S3). A second expression plasmid, pJOT2, contained a synthetic operon containing six genes, each preceded by a native C. bescii ribosome binding site (Additional file 1: Figure S2). Ribosome binding sites were designated as the 40 bp upstream of highly expressed C. bescii genes identified in a previous study of protein expression in the Caldicellulosiruptor genus (Blumer-Schuette et al. 2012). The individual gene ribosome binding site regions chosen for placement upstream of each gene are indicated in Additional file 1: Figure S2. Ferp_1184-Ferp_1187 encode for the four subunits of Benzoyl-CoA reductase, and Ferp_1370 is annotated as a benzoate transporter in the TIGRFAMs database. These genes, along with the before-mentioned benzoyl-CoA ligase, were codon optimized, arranged in a single transcriptional unit, and expressed from a single transcript also driven by the Athe_2303 promoter (Additional file 1: Figure S2, Table S2). This plasmid was transformed into strain JWCB018 to generate strain JWCB018 pJOT2 (Additional file 1: Table S3). An empty vector containing no expression insert, pJGW07, was also transformed into strain JWCB018, generating strain JWCB018 pJGW07, and subsequently used as an empty vector control in growth and activity experiments. Plasmid constructs and expression cassettes therein were validated by PCR, restriction digest, and Sanger sequencing of insert regions.
Growth conditions and medium used
Caldicellulosiruptor bescii strains were grown in 50 mL LOD medium (Farkas et al. 2013) in serum bottles of total volume of 135 mL. In media containing sodium benzoate, 80 mM MOPS (3-(N-morpholino)propanesulfonic acid) buffer (part# BP308-500, Fisher Scientific, Hampton, NH) was added, and resazurin was omitted as it was found to interfere with UV detection of sodium benzoate. Fermentations were carried out at 75 °C shaking at 200 rpm. Growth was monitored by optical density at 680 nm as done previously (Farkas et al. 2013).
HPLC determination of benzoate in medium supernatants
Benzoate present in media supernatants was determined using an Agilent 1290 Infinity HPLC and associated UV detector equipped with an Agilent Zorbax C18 Eclipse Plus reversed phase column (959757-902K, Agilent, Santa Clara, CA). The mobile phase used for elution was 79% monopotassium phosphate (pH = 2.5) and 21% acetonitrile. 20 μL of sample was injected into the column at a mobile phase flowrate of 1.5 mL/min. The column was maintained at 30 °C. Dilutions of sodium benzoate in LOD medium were used as standards to monitor retention time of sodium benzoate, and to generate a standard curve for quantitation. Integrated peak areas of absorbance at 230 nm was used to quantify benzoate, using the absorbance value at 360 nm as the reference.
Western blot and benzoyl-CoA activity assay
50 mL batch cultures of JWCB018 pJOT1 and JWCB018 pJGW07 were grown to mid-log cell density in medium containing sodium benzoate. Mid-log cells were centrifuged at 20,000×g at 4 °C for 4 min in a Piramoon fixed-angle FiberLite rotor (ThermoScientific, Waltham, MA), snap frozen in liquid nitrogen, and stored at − 80 °C. Cell pellets were resuspended in 1 mL of CelLytic B (part # B7435, Sigma Aldrich, St. Louis, MO) and sonicated for 6 cycles at 12 W for 15 s, with 1-min rest between cycles in a Misonix 3000 Sonicator (Misonix Incorporated, Farmingdale, NY). The sonicated mixture was centrifuged for 30 min at 4 °C at maximum speed in a microcentrifuge and the supernatant was retained as clarified cell extract. This extract was used in his-antibody immunodetection (western blot) of Ferp_1044 encoded protein as well as Benzoyl-CoA activity assays.
Toward immunodetection of his-tagged protein, 5 uL of cell lysate was electrophoresed on a Novex 4–20% Tris Glycine SDS Polyacrylamide gel (part # XP04202BOX, ThermoScientific, Waltham, MA) in an XCell SureLock Mini-Cell Electrophoresis Chamber (ThermoScientific, Waltham, MA) according to manufacturer’s instructions. The gel was subsequently placed in contact with a nitrocellulose membrane and protein was transferred to the membrane according to manufacturer’s protocols at 25 V for 90 min in a transfer buffer containing 1X Novex Tris–Glycine Transfer Buffer (Part # LC3675, ThermoScientific, Waltham, MA) and 10% methanol (v/v). Immunodetection was performed on the membrane the WesternBreeze Chromogenic Kit (anti-mouse reagents, ThermoScientific, Waltham, MA) according to manufacturer’s instructions. Monoclonal Anti-Polyhistidine Antibody (mouse IgG) was used as the primary antibody (R&D Systems, Minneapolis, MN).
Benzoyl-CoA Ligase activity was assayed in clarified cell lysates at 65 °C, 70 °C, and 75 °C using a previously described assay (Kawaguchi et al. 2006), though 25 uL of the aforementioned prepared crude cell lysate was used in place of purified protein. The assay was carried out in a Beckman Coulter DU 800 UV spectrophotometer (Beckman Coulter, Brea, CA) using UV transparent cuvettes (Plastibrand, Fisher Scientific, Hampton, NH).
RNA extraction, cDNA synthesis, and qPCR
RNA was extracted from 50 mL of mid-log grown batch culture replicates harvested and centrifuged at 20,000×g at 4 °C for four minutes, snap frozen in liquid nitrogen, and stored at − 80 °C. Total RNA was extracted by first incubating cell pellets in 250 µL of 20 mg/mL Lysozyme (Sigma Aldrich part number L-7651, St. Louis, MO) resuspended in SET buffer (50 mM Tris–HCl pH 8.0 50 mM EDTA, 20% w/v Sucrose) and incubated in a dry stationary bath at 37 °C for 8 min, vortexing briefly every 2 min. RNA was purified with a Qiagen RNEasy Kit according to manufacturer’s protocol (Qiagen, Hilden, Germany). RNA concentration was quantified with a Nanodrop 1000 instrument (ThermoScientific, Waltham, MA) and RNA quality was assessed via RNA Integrity Numbers (RIN) obtained with an Agilent 2100 Bioanalyzer and corresponding RNAchip (Agilent Technologies, Santa Clara, CA). cDNA was generated using a ScriptSeq II Kit (Illumina, San Diego, CA) according to manufacturer’s instructions. cDNA was diluted and used as template in qPCR reactions using Roche FastStart SYBR Green Master Mix (ThermoScientific, Waltham, MA) according to manufacturer’s instructions. Primers were designed to amplify a unique ~ 100 bp region within the CDS of each gene of interest. Primers used to quantify expression of each gene (relative to that of Athe_0001) are listed in Additional file 1: Table S1.
Quantitative proteomic determination of unique peptides derived from benzoate catabolism enzymes
Cell biomass was collected from 50 mL of mid-log grown batch culture replicates grown in medium containing sodium benzoate (see culture conditions methods). Strains assessed were JWCB018 pJOT2 and JWCB018 pJGW07. Mid-log phase culture replicates were centrifuged at 20,000×g at 4 °C for 4 min and snap frozen in liquid nitrogen. Protein was isolated from cell biomass and proteomic determination was carried out as described previously (Eminoğlu et al. 2017).
Results
Benzoate inhibits growth of C. bescii
The addition of sodium benzoate to the culture medium significantly inhibited the growth of C. bescii at concentrations as low as 1 mM (Fig. 1). Cultures containing benzoate yielded ~ 50% lower maximum cell density than unamended cultures. Cultures containing 10 mM sodium benzoate exhibited an average maximum growth rate which is 27% lower than unamended cultures (0.48 h−1 relative to 0.65 h−1 of unamended cultures). Benzoate was the most abundant monoaromatic compound identified in a previous anaerobic enrichment culture grown on radiolabeled lignin (Colberg and Young 1985), and it is a common food preservative with known antimicrobial activity. Therefore, benzoate accumulation during lignocellulosic decomposition could inhibit growth of this biocatalyst.

Caldicellulosiruptor bescii strain JWCB018 grown in increasing concentrations of sodium benzoate. Growth inhibition is observed at concentrations of 1 mM sodium benzoate and above. Error bars represent the standard deviation from analysis of 3 replicate cultures
Benzoyl-CoA ligase heterologous expression and activity
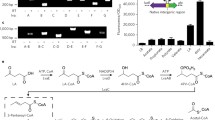
C. bescii strain JWCB018 pJOT1 was constructed to heterologously express a codon-optimized Ferroglobus placidus benzoyl-CoA ligase gene. This strain produced full length benzoyl-CoA ligase with a short affinity tag that enabled identification by Western blot analysis (Fig. 2). Furthermore, we observed benzoyl-CoA ligase activity in clarified cell extracts of this strain that was not present in a control strain that lacked the F. placidus benzoyl-CoA gene. Benzoyl-CoA ligase activity was detected at 65 °C (Table 1), but not when the assay was carried out at 70 °C or 75 °C (data not shown). The C. bescii expression platform used in this study has enabled C. bescii to produce ethanol (Chung et al. 2014; Chung et al. 2015a, b, c, d), detoxify furan aldehydes (Chung et al. 2015a, b, c, d), and increased cellulolytic capability (Chung et al. 2015a, b, c, d). Here we further expanded C. bescii’s repertoire of capabilities to activate a lignin degradation product.

Western blot image of whole-cell lysate from C. bescii expression strain JWCB018 pJOT1 showing a protein product of the size expected for heterologously expressed F. placidus benzoyl-CoA ligase protein as the major detected product. Whole-cell lysate was prepared, electrophoresed, and probed using anti-6× histidine antibody (see methods). Lane 1: MagicMark XP Western Protein Standard (ThermoFischer Scientific, Waltham, MA) protein masses (in kDa) indicated adjacent to each band, Lane 2: Lysate from strain JWCB018 pJOT1, Lane 3: Lysate from strain JWCB018 pJGW07 (hosting an empty expression vector)
Expression of the benzoyl-CoA reductase and putative benzoate transport genes
Caldicellulosiruptor bescii strain JWCB018 pJOT2 was constructed to express six F. placidus genes from a single synthetic operon whose expression was driven by the promoter of a native C. bescii gene encoding a highly abundant protein, the S-layer protein (Athe_2303), of C. bescii. These genes encoded a putative benzoate transporter, benzoyl-CoA ligase, and four subunits of benzoyl-CoA (Holmes et al. 2011, Holmes et al. 2012; Smith et al. 2015). RNAs transcribed from each gene were detected in varying abundance for all six genes by RT-qPCR (Fig. 3). However, unique peptides could only be identified from four of six genes using proteomic analysis (Fig. 3). Peptide evidence was not detected for one of the four subunits of benzoyl-CoA reductase (Ferp_1185), or for the putative benzoate transporter (Ferp_1370) encoded on the heterologous expression vector pJOT2 (Additional file 1: Figure S2). Furthermore, no in vivo catabolism of 10 mM sodium benzoate was observed during fermentation by C. bescii strain JWCB018 pJOT2 (Figs. 4 and 5) after 144 h of fermentation.

Relative expression of heterologously expressed benzoate catabolism genes in strain JWCB018 pJOT2 measured by RT-qPCR. Relative expression was calculated with respect to dnaA, a constitutively expressed chromosomal replication initiation protein (Athe_0001). Though all genes appeared to be transcribed, unique peptides were not identified for two proteins: Ferp_1370 (a putative benzoate transporter) and Ferp_1185 (one of four subunits of benzoyl-CoA reductase). *Heterologously expressed Ferroglobus placidus proteins were deemed present if normalized protein abundance was significantly (p value < 0.02, n = 3, log2 fold change > 1) greater in strain JWCB018 pJOT2 than in strain JWCB018 pJGW07. Numbers in parenthesis indicate percentages of each gene’s coding sequence represented by unique peptides identified through intracellular proteomics

Growth of C. bescii strains JWCB018 pJOT2 and JWCB018 pJGW07 in defined medium (containing 5 g/L maltose) supplemented with 10 mM sodium benzoate and 80 mM MOPS buffer (initial pH = 6.8). Error bars represent the standard deviation from analysis of 3 replicate cultures

Time course of benzoate concentration in cultures of C. bescii grown in defined medium containing 10 mM added sodium benzoate and 80 mM MOPS buffer. Benzoate concentrations do not decrease through 144 h of fermentation in the presence of C. bescii strain JWCB018 pJOT2. Error bars represent the standard deviation from analysis of 3 replicate cultures
Variation in protein production from heterologous gene expression may have been a result of transcript instability or strengths of the different native C. bescii ribosome binding sites used in this heterologous expression design. We did not see a trend of decreasing mRNA abundance along the length of the transcript, suggesting incomplete transcription and/or mRNA production was not the primary cause of variable expression among the genes. There is also no noticeable correlation between protein abundance and the relative amount of mRNA expression from any of the genes, suggesting either differing strengths of ribosome binding sites employed, varying rates of protein accumulation and turnover, or mRNA degradation may be reasons for the observed variation in protein production.
Discussion
Relatively low activity of heterologous benzoyl-CoA ligase
Benzoyl-CoA ligase activity from this strain was found to be four orders of magnitude smaller than comparable activity measurements in other organisms which natively host and express Benzoyl-CoA ligase (Schühle et al. 2003; Kawaguchi et al. 2006), suggesting expression and/or assay conditions are far from optimal. One reason for this observed discrepancy might be that the in vitro assay conditions do not represent conditions for optimal activity. Native intracellular levels of ATP are relatively low in C. bescii (Bielen et al. 2010), suggesting the primary source of phosphate cleavage derived energy is pyrophosphate and not ATP. Pyrophosphate is a product in the biochemical reaction carried out by benzoyl-CoA ligase, and carryover of this chemical in lysates used in in vitro assays may be inhibiting the activity of the heterologous benzoyl-CoA ligase. Expression of benzoyl-CoA ligase mRNA in strain JWCB018 pJOT2 was found to be 2–3 fold higher than the housekeeping gene dnaA (Fig. 3). Other assessments of benzoyl-CoA ligase activity have been done using lysates that were prepared from cells natively hosting this enzyme (Geissler et al. 1988; Kawaguchi et al. 2006), where elevated concentrations of added sodium benzoate are known to strongly induce expression of benzoyl-CoA ligase (Holmes et al. 2011). In this study, the bcl gene was expressed from a constitutive promoter (Additional file 1: Figure S1), and the sodium benzoate contained in medium used in this study noticeably inhibited C. bescii growth (Fig. 1). Increasing heterologous benzoyl-CoA ligase activity (and possibly that of other heterologous aromatic catabolism enzymes) in C. bescii may be realized by increasing production of the enzyme, increasing intracellular concentrations of ATP, and/or decreasing the intracellular pyrophosphate concentration.
Redox considerations of benzoyl-CoA reduction
Additional genes that may be necessary for benzoate catabolism in F. placidus may be needed to effect benzoate catabolism in C. bescii. Ferp_1180 encodes a ferredoxin that is co-expressed and displays the same benzoate-driven expression patterns as other aromatic catabolism genes in F. placidus (Holmes et al. 2012). The benzoyl-CoA reductase is annotated as being ferredoxin dependent (Holmes et al. 2012), although reduction via this enzyme has been demonstrated using a number of different electron donors (Schmid et al. 2015). This particular ferredoxin, as well as other genes found to be co-regulated in response to benzoate (Holmes et al. 2012), may be necessary for proper functioning of this enzyme.
In other species, oxidoreductase genes are needed to re-reduce the low potential ferredoxin needed for benzoyl-CoA reductase mediated reduction (Carmona et al. 2009). Homologs of the Ferp_1033–Ferp_1034 genes, encoding a 2-oxoglutarate ferredoxin oxidoreductase (Holmes et al. 2012; Boll et al. 2014), were shown to be important to this recycle, and supplying reduced ferredoxin, in Thauera aromatica (Dörner and Boll, 2002). A gene encoding one subunit of 2-oxoglutarate ferredoxin oxidoreductase in F. placidus was found to have increased expression when grown on aromatic substrates (Holmes et al. 2012). The expression of 2-oxoglutarate ferredoxin oxidoreductase from F. placidus (Ferp_1033–Ferp_1034) may also serve to increase benzoyl-CoA reductase activity in vivo in C. bescii.
It was found that the heterologous putative benzoate transporter was unable to be expressed from the synthetic operon construct in strain JWCB018 pJOT2. As such, another reason no benzoate degradation was observed in this strain (Fig. 5) was that this strain was unable to import benzoate. The effectiveness of the protein product from the gene Ferp_1370 has not been tested for its ability to facilitate benzoate import into a bacterium.
Abbreviations
- bp:
-
basepairs (of DNA)
- qPCR:
-
quantitative polymerase chain reaction
- PCR:
-
polymerase chain reaction
- CoA:
-
coenzyme A
- EDTA:
-
ethylenediaminetetraacetic acid
- CBP:
-
consolidated bioprocessing
- BCL:
-
benzoyl-coenzyme A ligase
- DOE:
-
U.S. Department of Energy
References
Bielen AAM, Willquist K, Engman J, Van DO, Van Niel EdWJ, Kengen SWM (2010) Pyrophosphate as a central energy carrier in the hydrogen-producing extremely thermophilic Caldicellulosiruptor saccharolyticus. FEMS Microbiol Lett 307:48–54
Blumer-Schuette S, Giannone RJ, Zurawski JV, Ozdemir I, Ma Q, Yin Y, Xu Y, Kataeva I, Poole FL, Adams MWW, Hamilton-Brehm S, Elkins JG, Larimer FW, Land ML, Hauser LJ, Cottingham RW, Hettich RL, Kelly RM (2012) Caldicellulosiruptor core and pangenomes reveal determinants for noncellulosomal thermophilic deconstruction of plant biomass. J Bacteriol 194:4015–4028
Blumer-Schuette S, Brown SD, Sander KB, Bayer EA, Kataeva I, Zurawski JV, Conway JM, Adams MW, Kelly RM (2014) Thermophilic lignocellulose deconstruction. FEMS Microbiol Rev 38:393–448
Boll M, Löffler C, Morris BEL, Kung JW (2014) Anaerobic degradation of homocyclic aromatic compounds via arylcarboxyl-coenzyme A esters: organisms, strategies and key enzymes. Environ Microbiol 16:612–627
Brunecky R, Alahuhta M, Xu Q, Donohoe BS, Crowley MF, Kataeva IA, Yang S, Resch MG, Adams MWW, Lunin VV, Himmel ME, Bomble YJ (2013) Revealing nature’s cellulase diversity: the digestion mechanism of Caldicellulosiruptor bescii CelA. Science 342:1513–1516
Carmona M, Zamarro MT, Blázquez B, Durante-Rodríguez G, Juárez JF, Valderrama JA, Barragán MJL, García JL, Díaz E (2009) Anaerobic catabolism of aromatic compounds: a genetic and genomic view. Microbiol Mol Biol Rev 73:71–133
Chung D, Farkas J, Westpheling J (2013a) Overcoming restriction as a barrier to DNA transformation in Caldicellulosiruptor species results in efficient marker replacement. Biotechnol Biofuel 6:82
Chung D, Cha M, Farkas J, Westpheling J (2013b) Construction of a stable replicating shuttle vector for Caldicellulosiruptor species: use for extending genetic methodologies to other members of this genus. PLoS ONE 8:e62881
Chung D, Cha M, Guss AM, Westpheling J (2014) Direct conversion of plant biomass to ethanol by engineered Caldicellulosiruptor bescii. Proc Natl Acad Sci USA 111:8931–8936
Chung D, Young J, Bomble YJ, Vander Wall TA, Groom J, Himmel ME, Westpheling J (2015a) 4370642; homologous expression of the Caldicellulosiruptor bescii CelA reveals that the extracellular protein is glycosylated. PLoS ONE 10:e0119508
Chung D, Verbeke T, Cross K, Westpheling J, Elkins J (2015b) Expression of a heat-stable NADPH-dependent alcohol dehydrogenase in Caldicellulosiruptor bescii results in furan aldehyde detoxification. Biotechnol Biofuel 8:1–11
Chung D, Cha M, Snyder E, Elkins J, Guss A, Westpheling J (2015c) Cellulosic ethanol production via consolidated bioprocessing at 75 °C by engineered Caldicellulosiruptor bescii. Biotechnol Biofuel 8:163
Chung D, Young J, Cha M, Brunecky R, Bomble Y, Himmel M, Westpheling J (2015d) Expression of the Acidothermus cellulolyticus E1 endoglucanase in Caldicellulosiruptor bescii enhances its ability to deconstruct crystalline cellulose. Biotechnol Boifuel 8:1–9
Clarkson SM, Kridelbaugh DM, Elkins JG, Guss AM, Michener J (2017) Construction and optimization of a heterologous pathway for protocatechuate catabolism in Escherichia coli enables rapid bioconversion of model lignin monomers. Appl Environ Microbiol 83:1313
Colberg PJ, Young LY (1985) Anaerobic degradation of soluble fractions of [14C-lignin] lignocellulose. Appl Environ Microbiol 49:345–349
Dörner E, Boll M (2002) Properties of 2-oxoglutarate: ferredoxin oxidoreductase from Thauera aromatica and its role in enzymatic reduction of the aromatic ring. J Bacteriol 184:3975–3983
Eminoğlu A, Murphy SJ, Maloney M, Lanahan A, Giannone RJ, Hettich RL, Tripathi SA, Beldüz AO, Lynd LR, Olson DG (2017) Deletion of the hfsB gene increases ethanol production in Thermoanaerobacterium saccharolyticum and several other thermophilic anaerobic bacteria. Biotechnol Biofuel 10:282
Farkas J, Chung D, Cha M, Copeland J, Grayeski P, Westpheling J (2013) Improved growth media and culture techniques for genetic analysis and assessment of biomass utilization by Caldicellulosiruptor bescii. J Ind Microbiol Biotechnol 40:41–49
Fuchs G, Boll M, Heider J. 2011. Microbial degradation of aromatic compounds—from one strategy to four. Nat Rev Micro 9:803–816
Geissler JF, Harwood CS, Gibson J (1988) Purification and properties of benzoate-coenzyme A ligase, a Rhodopseudomonas palustris enzyme involved in the anaerobic degradation of benzoate. J Bacteriol 170:1709–1714
Hamilton-Brehm S, Mosher JJ, Vishnivetskaya T, Podar M, Carroll S, Allman S, Phelps TJ, Keller M, Elkins JG (2010) Caldicellulosiruptor obsidiansis sp. nov., an anaerobic, extremely thermophilic, cellulolytic bacterium isolated from obsidian pool, Yellowstone National Park. Appl Environ Microbiol 76:1014–1020
Harwood CS, Burchhardt G, Herrmann H, Fuchs G (1998) Anaerobic metabolism of aromatic compounds via the benzoyl-CoA pathway. FEMS Microbiol Rev 22:439–458
Holmes DE, Risso C, Smith JA, Lovley DR (2011) Anaerobic oxidation of benzene by the hyperthermophilic archaeon Ferroglobus placidus. Appl Environ Microbiol 77:5926–5933
Holmes DE, Risso C, Smith JA, Lovley DR (2012) Genome-scale analysis of anaerobic benzoate and phenol metabolism in the hyperthermophilic archaeon Ferroglobus placidus. ISME J 6:146–157
Kataeva I, Foston MB, Yang S, Pattathil S, Biswal AK, Poole Ii FL, Basen M, Rhaesa AM, Thomas TP, Azadi P, Olman V, Saffold TD, Mohler KE, Lewis DL, Doeppke C, Zeng Y, Tschaplinski TJ, York WS, Davis M, Mohnen D, Xu Y, Ragauskas AJ, Ding S, Kelly RM, Hahn MG, Adams MWW (2013) Carbohydrate and lignin are simultaneously solubilized from unpretreated switchgrass by microbial action at high temperature. Energy Environ Sci 6:2186–2195
Kawaguchi K, Shinoda Y, Yurimoto H, Sakai Y, Kato N (2006) Purification and characterization of benzoate-CoA ligase from Magnetospirillum sp. strain TS-6 capable of aerobic and anaerobic degradation of aromatic compounds. FEMS Microbiol Lett 257:208–213
Lynd LR, van Zyl WH, McBride JE, Laser M (2005) Consolidated bioprocessing of cellulosic biomass: an update. Curr Opin Biotechnol 16:577–583
Lynd LR, Laser MS, Bransby D, Dale BE, Davison B, Hamilton R, Himmel M, Keller M, McMillan JD, Sheehan J, Wyman CE (2008) How biotech can transform biofuels. Nat Biotechnol 26:169
Schmid G, René SB, Boll M (2015) Enzymes of the benzoyl-coenzyme A degradation pathway in the hyperthermophilic archaeon Ferroglobus placidus. Environ Microbiol 17:3289–3300
Schmid G, Auerbach H, Pierik AJ, Schünemann V, Boll M (2016) ATP-dependent electron activation module of benzoyl-coenzyme A reductase from the hyperthermophilic archaeon Ferroglobus placidus. Biochemistry 55:5578–5586
Schühle K, Gescher J, Feil U, Paul M, Jahn M, Schägger H, Fuchs G (2003) Benzoate-coenzyme A ligase from Thauera aromatica: an enzyme acting in anaerobic and aerobic pathways. J Bacteriol 185:4920–4929
Smith JA, Aklujkar M, Risso C, Leang C, Giloteaux L, Holmes DE (2015) Mechanisms involved in fe(III) respiration by the hyperthermophilic archaeon Ferroglobus placidus. Appl Environ Microbiol 81:2735–2744
Tor JM, Lovley DR (2001) Anaerobic degradation of aromatic compounds coupled to fe(III) reduction by Ferroglobus placidus. Environ Microbiol 3:281–287
Yang S, Kataeva I, Hamilton-Brehm S, Engle NL, Tschaplinski TJ, Doeppke C, Davis M, Westpheling J, Adams MWW (2009) Efficient degradation of lignocellulosic plant biomass, without pretreatment, by the thermophilic anaerobe “Anaerocellum thermophilum” DSM 6725. Appl Environ Microbiol 75:4762–4769
Zamarro MT, Barragán MJL, Carmona M, García JL, Díaz E (2017) Engineering a bzd cassette for the anaerobic bioconversion of aromatic compounds. Microb Biotechnol 10:1418–1425
Authors’ contributions
KS helped conceive of the study, design experiments, generate expression constructs and bacterial strains used, collect and analyze data, interpret results, draft and edit the manuscript. MY helped generate expression constructs and bacterial strains, collect and analyze data, and edit the manuscript. KM helped collect and analyze sodium benzoate HPLC data. JW helped collect and analyze sodium benzoate HPLC data, and generate and analyze western blot analysis. RJG performed and helped interpret proteomic analysis. SDB helped conceive of study, interpret results, and edit the manuscript. MR helped collect and analyze sodium benzoate HPLC data. DEG provided expertise and guidance in enzymatic assay for Benzoyl-CoA activity, helped interpret results, and draft the manuscript. BH helped conceive of the study, generate expression constructs, and edit the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The authors would like to acknowledge Dale Pelletier, Michael W.W. Adams, and Farris Poole for valuable discussions, assistance with analysis, and critical review of the manuscript. The authors would like to acknowledge Dawn Klingeman for valuable insight and assistance with RNA extraction, cDNA generation and qPCR quantification. The authors would like to also acknowledge Dr. Janet Westpheling for the kind gift of the C. bescii strain JWCB018 and the plasmid pJGW07 used in this study.
This manuscript has been authored by UT-Battelle, LLC, under contract DE-AC05-00OR22725 with the US Department of Energy (DOE). The US government retains and the publisher, by accepting the article for publication, acknowledges that the US government retains a nonexclusive, paid-up, irrevocable, worldwide license to publish or reproduce the published form of this manuscript, or allow others to do so, for US government purposes. DOE will provide public access to these results of federally sponsored research in accordance with the DOE Public Access Plan (http://energy.gov/downloads/doe-public-access-plan).
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
All data supporting the claims of this manuscript are presented and made available in this manuscript and supporting information. The wildtype C. bescii strain is available through DSMZ (as C. bescii strain DSMZ 6725), and the base strain used for experiments herein; JWCB018 (Chung et al. 2013a, b) was a kind gift of, and available upon request from, Dr. Janet Westpheling, Professor in the Department of Genetics at the University of Georgia. The codon-optimized gene sequences have been deposited into GenBank and can be retrieved at the following accession numbers: Ferp_1370_codon_optimized; MK294010, Ferp_1044_codon_optimized; MK294011, Ferp_1184_codon_optimized; MK294012, Ferp_1185_codon_optimized; MK294013, Ferp_1186_codon_optimized; MK294014, Ferp_1187_codon_optimized; MK294015.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Funding
This work was supported by the BioEnergy Science Center (BESC), which is a U.S. Department of Energy (DOE) Bioenergy Research Center supported by the Office of Biological and Environmental Research in the DOE Office of Science. This research was supported in part by appointments to the Higher Education Research Experiences Program and the ASTRO Programs at Oak Ridge National Laboratory, administered by ORAU through the DOE Oak Ridge Institute for Science and Education. This work was supported in part by the U.S. Department of Energy, Office of Science, and Office of Workforce Development for Teachers and Scientists (WDTS) under the Visiting Faculty Program (VFP) administered by the Oak Ridge Institute for Science and Education (ORISE). Oak Ridge National Laboratory is managed by UT-Battelle LLC for DOE under contract DE-AC05-00OR22725.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Additional file
Additional file 1: Figure S1.
Diagram of plasmid pJOT1 used to heterologously express a codon-optimized Ferp_1044 gene from Ferroglobus placidus, encoding a benzoyl-CoA ligase, in C. bescii. Figure S2. Diagram of plasmid pJOT2 used to heterologously express codon-optimized genes from Ferroglobus placidus encoding a benzoyl-CoA ligase (Ferp_1044), a benzoyl-CoA reductase (Ferp_1184–Ferp_1187), and a putative benzoate transporter (Ferp_1370). Unique ribosome binding site regions (~ 40 upstream basepairs) were identified from highly transcribed C. bescii genes (Blumer-Schuette et al. 2012) and placed upstream of each gene. Table S1. Primers used in this study. Table S2. Plasmids used in this study.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Sander, K., Yeary, M., Mahan, K. et al. Expression of benzoyl-CoA metabolism genes in the lignocellulolytic host Caldicellulosiruptor bescii. AMB Expr 9, 59 (2019). https://doi.org/10.1186/s13568-019-0783-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13568-019-0783-8