
Abstract
Background
DNMT3A gene mutation has been associated with poor prognosis in acute myeloid leukemia, but its clinical implications in myelodysplastic syndrome (MDS) and dynamic changes during disease progression remain controversial.
Results
In this study, DNMT3A mutation was identified in 7.9% of 469 de novo MDS patients. DNMT3A-mutated patients had higher platelet counts at diagnosis, and patients with ring sideroblasts had the highest incidence of DNMT3A mutations, whereas those with multilineage dysplasia had the lowest incidence. Thirty-one (83.8%) of 37 DNMT3A-mutated patients had additional molecular abnormalities at diagnosis, and DNMT3A mutation was highly associated with mutations of IDH2 and SF3B1. Patients with DNMT3A mutations had a higher risk of leukemia transformation and shorter overall survival. Further, DNMT3A mutation was an independent poor prognostic factor irrespective of age, IPSS-R, and genetic alterations. The sequential study demonstrated that the original DNMT3A mutations were retained during follow-ups unless allogeneic hematopoietic stem cell transplantation was performed, while DNMT3A mutation was rarely acquired during disease progression.
Conclusions
DNMT3A mutation predicts unfavorable outcomes in MDS and was stable during disease evolutions. It may thus be a potential biomarker to predict prognosis and monitor the treatment response.
Similar content being viewed by others


Background
Alterations of epigenetic regulation may result in aberrations of gene expression and malignant transformation of cells [1, 2]. DNA methylation, one of the mechanisms for epigenetic control of gene expression, regulates important physiological development, such as gene imprint and X-chromosome inactivation [3, 4]. In mammalians, three DNA methyltransferase (DNMT), namely DNMT1, 3A, and 3B have been identified [5]. Mutation of DNMT3A gene has been reported in patients with myeloid malignancies, including myelodysplastic syndromes (MDS) and acute myeloid leukemia (AML) [6,7,8,9,10,11,12,13].
MDS represent a clinically heterogeneous hematologic neoplasm characterized by variable degrees of cytopenias and risk of leukemia transformation [14]. The incidence (2.6 to 20.2%) of DNMT3A mutation in MDS varied widely, possibly due to different patient population and methods used [12, 13, 15, 16]. Regarding the prognostic relevance, DNMT3A mutation has been reported to predict poor prognosis in AML patients [7,8,9,10,11]. However, the prognostic implications of DNMT3A mutation in MDS are still controversial [12, 13, 15, 17]. Walter et al. [12] and Thol et al. [13] reported that DNMT3A mutation was associated with higher risk of leukemia transformation and shorter survival, but the other studies failed to find these associations [15, 17]. Besides, sequential studies to evaluate the dynamic changes of DNMT3A mutations during disease evolution in MDS are limited. In the present study, we investigated the DNMT3A mutation in 469 patients with de novo MDS and analyzed its associations with the clinical characteristics, outcomes, and other genetic alterations. We also performed sequential analysis of the DNMT3A gene mutation for 431 samples from 148 patients to evaluate the stability of DNMT3A mutation during the clinical course.
Methods
Subjects
This study was approved by the Institutional Review Board/Ethical Committee of the National Taiwan University Hospital (NTUH). Diagnosis and classification of MDS were made according to the French-American-British (FAB) Cooperative Group Criteria and the WHO 2016 classification [18, 19]. From May 1985 to December 2010, a total of 469 adult patients with newly diagnosed MDS at the NTUH who had enough cryopreserved cells for analysis were enrolled. Patients with secondary or therapy-related MDS were excluded. The disease of 362 patients fulfilled the criteria of MDS according to the 2016 WHO classification. Most patients (77.4%) received only palliative treatment, including transfusions, hematopoietic growth factors, immunosuppressive therapy, and low-intensity chemotherapy. Thirty (6.4%) patients received intensive chemotherapy, 7.2% received hypomethylating agents (HMA), and 9.0% received allogeneic hematopoietic stem cell transplantation (HSCT).
Analyses of mutations
Mutational analysis of DNMT3A gene exons 2-23 by PCR and direct sequencing was done as described previously [9]. Analysis of the mutations in other genes involving in activated signaling pathways, such as FLT3-ITD [20], NRAS [21], KRAS [21], JAK2 [21], and PTPN11 [22]; the transcription factor, such as RUNX1 [23]; splicing factors, including SRSF2, U2AF1, and SF3B1 [24]; and epigenetic modifications, including MLL/PTD [25], ASXL1 [26], EZH2 [27], IDH1 [28], IDH2 [29], and TET2 [30], as well as SETBP1 [21], WT1 [31], NPM1 [32], and TP53 [33], were performed as previously described. To detect DNMT3A mutation, we used DNA amplified in vitro from bone marrow (BM) cells with the Illustra GenomiPhi V2 DNA-amplification kit (GE Healthcare, UK). All mutations detected were verified in the original non-amplified samples [34]. Abnormal sequencing results were confirmed by at least two repeated analyses. All nonsense or frameshift mutations were regarded as true mutations. Missense mutations were regarded as true only if they were documented in other studies or could be verified by sequencing of matched normal somatic tissues. Serial analyses of DNMT3A mutations during the clinical course were also performed in 431 samples from 148 patients.
TA cloning analysis
For the patients with discrepancy of the mutation status of the DNMT3A in sequential samples, TA cloning was performed in the samples without detectable mutation followed by direct sequencing. More than 30 clones were selected for sequencing as previously described [9].
Illumina next generation sequencing (NGS) for serial studies of patients with DNMT3A mutation
Serial analyses of mutations at diagnosis, disease progression, and/or remission were further performed using Illumina next generation sequencing in 32 samples from 13 patients with DNMT3A mutation at diagnosis and one during follow-up. Genomic DNA extracted from BM mononuclear cells was analyzed for mutations in 54 genes involved in myeloid malignancies by TruSight Myeloid Panel (Illumina, San Diego, CA, USA). HiSeq platform (Illumina) was used for sequencing with a median reading depth of 12,000×.
Statistical analysis
The discrete variables of patients with and without gene mutations were compared using the χ2 tests, and the Fisher’s exact test was used if the expected values of contingency tables were smaller than 5. The continuous variables of patients with and without gene mutations were compared using Student’s t test. If the data were not normally distributed, Mann-Whitney U tests were used to compare continuous variables and medians of distributions. Overall survival (OS) was measured from the date of first diagnosis to the date of last follow-up or death from any cause. Time to leukemia transformation was measured from the date of MDS diagnosis to the date confirmed of acute leukemic change. Kaplan-Meier estimation was used to plot survival curves, and log-rank tests were used to calculate the difference of OS and time to leukemia transformation between groups. Multivariate Cox proportional hazard regression analysis was used to investigate independent prognostic factors for OS and time to leukemia transformation. All tests were 2-tailed, and P < 0.05 was considered statistically significant. All statistical analyses were performed with SPSS Version 17 software.
Results
DNMT3A mutations in patients with de novo MDS
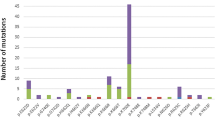
A total of 469 patients with de novo MDS according to the FAB classification were included for mutational analysis. Among them, 171 (36.5%) patients had refractory anemia (RA), 32 (6.8%) had RA with ring sideroblasts (RARS), 159 (33.9%) had RA with excess blasts (RAEB), 53 (11.3%) had RAEB in transformation (RAEB-T), and 54 (11.5%) had chronic myelomonocytic leukemia (CMML) (Table 1). Nineteen different DNMT3A mutations were identified in 37 (7.9%) of the 469 patients, including 7 missense mutations, 2 nonsense mutations, and 10 frameshift mutations (Fig. 1). In addition to the 13 single-nucleotide polymorphisms without amino acid residue alterations, 9 missense mutations with uncertain biologic significance were excluded because they were not reported previously and could not be verified for lack of matched normal somatic tissues or remission BM samples. Thirty-six of the 37 DNMT3A-mutated patients had single heterozygous mutation, and the remaining one patient had double mutations. The most common mutation was R882H (n = 11), followed by R882C (n = 8), G543C (n = 2), and Y735C (n = 2). All other mutations were detected in only one patient each (Fig. 1; Additional file 1: Table S1). According to the 2016 WHO classification, DNMT3A mutations were identified in 28 (7.7%) of the 362 patients (Table 1).

Patterns and locations of DNMT3A mutations
Correlation of DNMT3A mutations with clinical features
DNMT3A-mutated patients had higher platelet counts at diagnosis than DNMT3A-wild patients (Table 1). According to the FAB classification, DNMT3A mutation was highly associated with RARS subtype. Patients with RARS had the highest incidence (18.8%, P = 0.031) of DNMT3A mutations, whereas those with RA had the lowest incidence (2.3%, P = 0.001). By the 2016 WHO classification, patients with MDS with multilineage dysplasia (MDS-MLD) had lower incidence of DNMT3A mutations (2.8%, P = 0.029). No association between age or gender of patients and DNMT3A mutation status was found (Table 1). There was also no difference in the distribution of risk groups according to the international prognostic scoring system (IPSS) or revised IPSS (IPSS-R) between patients with and without DNMT3A mutations (Table 1).
Chromosome data were available in 437 (93.2%) patients at diagnosis, and clonal chromosomal abnormalities were detected in 193 (44.2%) patients. There were no association of the DNMT3A mutations with common chromosomal abnormalities, including loss of Y, − 20/del(20q), − 5/del(5q), + 8, and − 7/del(7q) (Additional file 1: Table S2), or risks of karyotype (Table 1).
Association of DNMT3A mutation with other genetic mutations
Among the 37 patients with DNMT3A mutations, 33 (89.2%) patients had additional molecular abnormalities at diagnosis, including SF3B1 (n = 11), TET2 (n = 7), IDH2 (n = 7), RUNX1 (n = 7), ASXL1 (n = 5), SRSF2 (n = 5), TP53 (n = 4), N-RAS (n = 2), MLL/PTD (n = 2), U2AF1 (n = 2), NPM1 (n = 2), K-RAS (n = 1), and IDH1 mutations (n = 1) (Additional file 1: Table S1). Fifteen patients had 1 additional mutation, 13 had 2, and 5 had 3 (Additional file 1: Table S1). Patients with DNMT3A mutations had a significantly higher incidence of SF3B1 and IDH2 mutations than those without DNMT3A mutations (P < 0.001 and P < 0.001, respectively; Table 2).
Correlation of DNMT3A mutation with clinical outcome
We could not find the difference in treatment regimens between the patients with DNMT3A mutations and those without. With a median follow-up of 43.9 months (range 0.1–250.7 months), patients with DNMT3A mutations had a higher risk to transform to AML (5-year AML transformation rate, 34.4 versus 22.5%, P = 0.013; Fig. 2). MDS patients, based on either the FAB or the 2016 WHO classification, had a significantly shorter OS if they harbored DNMT3A mutation than those who did not (15.0 versus 32.5 months, P = 0.024, and 16.3 versus 41.6 months, P = 0.011, respectively; Figs. 3 and 4). Further, we could not find the survival difference between the patients with frameshift and non-frameshift mutations. Interestingly, patients with DNMT3A mutations had a better OS if they received allogenic HSCT than those who did not (P = 0.038, Additional file 1: Figure S1).

Kaplan-Meier curves stratified by the status of DNMT3A mutations for time to leukemia transformation among the whole cohort of 469 MDS patients according to the FAB classification

Kaplan-Meier curves stratified by the status of DNMT3A mutations for overall survival among the whole cohort of 469 MDS patients according to the FAB classification

Kaplan-Meier curves stratified by the status of DNMT3A mutations for overall survival among 362 patients based on the 2016 WHO classification
Because DNMT3A mutation was closely associated with SF3B1 mutation, a good prognostic factor in MDS patients [35, 36], we divided the whole cohort to two subgroups, SF3B1-mutated and SF3B1-wild type, to evaluate the prognostic significance of DNMT3A mutation independent of SF3B1 mutation. In the SF3B1-wild patients, DNMT3A mutation predicted worse prognosis (OS, 14.6 ± 4.7 months versus 30.9 ± 3.2 months, P = 0.005). On the other hand, in the 48 SF3B1-mutated patients, DNMT3A mutation had no prognostic implication (OS, 17.7 ± 11.0 months versus 39.7 ± 4.2 months, P = 0.858) (Additional file 1: Figure S2).
Intriguingly, the impact of DNMT3A mutation on OS and time to leukemia transformation remained significant after adjusting the effects of age, gender, IPSS-R [37, 38], and mutations with prognostic significance in multivariate Cox regression analysis (FAB defined patients: OS: hazard ratio, HR 1.733, 95% CI 1.118–2.688, P = 0.014; time to leukemia transformation: HR 3.088, 95% CI 1.574–6.056, P = 0.001; 2016 WHO classification defined patients: OS: HR 1.800, 95% CI 1.080–3.000, P = 0.024; time to leukemia transformation: HR 2.360, 95% CI 1.129–4.933, P = 0.022; Table 3).
Sequential studies of DNMT3A mutations
To investigate the role of DNMT3A mutation in clinical evolution, DNMT3A gene mutation status was sequentially tested during the clinical course in 431 samples from 148 patients, including 13 patients with DNMT3A mutations at diagnosis and 135 patients without the mutation. In the 13 DNMT3A-mutated patients, 8 had disease progression, including 6 [unique patient numbers (UPNs) 1, 5, 7, 13, 24, and 37] with AML transformation. Four patients (UPNs 17, 24, 30, and 36) lost the original DNMT3A and other concurrent mutations/cytogenetic abnormalities when complete remission (CR) was achieved following curative-intent chemotherapy and/or allogeneic HSCT (Table 4). On the other hand, the other 9 patients with DNMT3A mutations at diagnosis retained their mutations during follow-ups. Among the eight with disease progression, one (UPN 37) acquired a novel RUNX1 mutation when the disease transformed to AML.
Among the 135 patients without DNMT3A mutation at diagnosis, 1 (0.7%) patient (UPN 47) acquired a novel DNMT3A mutation during sequential follow-up. This patient had MDS with excess blasts-1 (MDS-EB1) at diagnosis when no DNMT3A mutation was detectable even using more sensitive cloning method and next generation sequencing. He acquired GNAS, ASXL1, and ZRSR2 mutations in addition to DNMT3A mutation in the 19th month and died of progressive cytopenia in the 29th month.
We further analyzed the variant allele frequencies of the mutations in the 48 DNMT3A-mutated patients by NGS (Table 4). The mutant burden of DNMT3A mutations at diagnosis ranged from 8.4 to 45.24% with a median of 31.1%. Among the 13 patients with serial studies during the clinical courses, the mutation burden at subsequent follow-ups, compared to that at diagnosis, was increased in 3 patient (UPNs 5, 13, and 24), decreased in 6 patients (UPNs 1, 7, 10, 17, 30, and 36, Table 4) and stationary in 4 patients (UPNs 21, 23, 27, and 37). All of the three patients with increased DNMT3A mutation burden had leukemia transformation. Their variant allele frequencies of DNMT3A and other co-occurring mutations were increased at least 10% (10.0–347.1%) at leukemia transformation compared with those at baseline. The patient (UPN 37) who had least increase in variant allele frequency of DNMT3A mutation during disease progression acquired RUNX1 mutation at leukemia transformation. In contrast, the variant allele frequencies of DNMT3A and other concurrent mutations were relative stationary or even decreased during follow-up in the patients without leukemia transformation.
Discussion
In the present study, we identified 19 different DNMT3A mutations in 37 (7.9%) of the 469 FAB-defined and 7.7% of the 2016 WHO-defined MDS patients. Similar to previous studies on AML or MDS cohorts [7,8,9,10, 12, 13, 17], most mutations are located in the MTase domain, especially at amino acid R882 locus. Of these 19 mutations, 10 are frameshift and 2 are nonsense mutations. They generate truncated peptides with complete or partial deletion of the MTase and are expected to abolish the normal function of DNMT3A gene. The R882 mutations result in impaired gene function [7, 39], but the influence of the remaining missense mutations on the enzyme activity are unclear. In this study, the prevalence of DNMT3A mutation is 7.9 and 7.7% in MDS according to the FAB and 2016 WHO classification, respectively (Table 1), similar to most of the previous reports (7.8 to 10%) [12, 40,41,42] but higher than that of Thol et al. (2.6%) [13].
The reports with detailed demographics of MDS patients with DNMT3A mutation in literature are limited. In the report of Walter et al., but not in the current study and other studies [40, 42], DNMT3A mutations were associated with older age; in contrast, DNMT3A mutations were associated with higher platelet count in our study but not in other studies [12, 40, 42]. The association of DNMT3A mutations with higher platelet count was also shown in AML in previous studies [8, 9]. No comparison of age and hemogram between patients with and without DNMT3A was done in the study of Thol et al. [13] in which only five patients were found to have DNMT3A mutation. The causes of differences in the incidence of DNMT3A mutation and the clinical characteristics of DNMT3A-mutated patients might result from the differences in patient population recruited, detection platform used, sample size, and DNMT3A regions screened. In the study of Thol et al. [13], exons 15-23 instead of exons 2-23 of DNMT3A gene were analyzed in most patients (173 of 193 patients). Therefore, some patients harboring DNMT3A mutations might not be detected, and this might partially explain the lower incidence of DNMT3A mutation in their cohort (2.6%).
In this study, DNMT3A mutations were positively associated with IDH2 and SF3B1 mutations (Table 2). The close association of DNMT3A and IDH2 mutations was also shown in AML [9]. Mutations of DNMT3A and SF3B1, a component of spliceosome complex frequently mutated in RARS, have been reported to occur concurrently more often than expected by chance in lower-risk MDS patients [17]. In our cohort, the positive association of these two genetic alterations could also be found in lower-risk MDS patients (P < 0.001; Table 2). In addition, we could find a trend of positive correlation between these two mutations in higher-risk MDS patients (P = 0.098; Table 2). The close associations between DNMT3A mutation and RARS and between DNMT3A and SF3B1 mutations in this study (Table 1) might be related with each other. To investigate the associations among the RARS subtype, DNMT3A mutation, and SF3B1 mutation, we divided the whole cohort to RARS and non-RARS patients. The close association of DNMT3A and SF3B1 mutations retained in both subgroups. In contrast, no association between DNMT3A mutation and RARS subtype was found when we divided the whole population to SF3B1-mutated and SF3B1 wild-type patients. In the studies of more than 100 genes by high-throughput DNA sequencing, Haferlach et al. [43] and Papaemmanuil et al. [44] also found a positive correlation between DNMT3A and SF3B1 mutations, indicating that interaction between these two gene mutations may play a role in the pathogenesis of MDS, but further investigations are needed to elucidate its mechanism, especially in RARS subtype. No data regarding the association between DNMT3A mutation and RARS were shown in these two studies.
DNMT3A mutation has been identified as a poor prognostic factor in AML patients [7,8,9,10,11]. However, its prognostic impact on MDS patients remains uncertain. Walter et al. demonstrated DNMT3A mutations were associated with shorter survival and higher risk of leukemia transformation in univariate analysis [12], and Thol et al. also reported a higher rate of transformation to AML in patients with this mutation [13]. However, three other studies did not reveal significant impact of DNMT3A mutations on survival [15, 17, 44]. In this study, we showed that DNMT3A mutation was associated with poor outcomes, including higher risk of AML transformation and shorter OS. Bejar et al. [17] had speculated that the negative prognostic effect of DNMT3A mutation might be mitigated by the co-existence of SF3B1 mutation. In their cohort, 22% patients had SF3B1 mutation and they did not find the prognostic significance of DNMT3A mutation. The same was also true in another study, in which 24% of patients had SF3B1 mutation [44]. Both cohorts had significantly higher incidence of SF3B1 mutation than ours (10.2%). It may be possible that DNMT3A mutation would have prognostic effect only in MDS cohorts with low prevalence of SF3B1 mutation. Nevertheless, we distinctly showed that DNMT3A mutation was an independent poor prognostic factor for OS irrespective of the status of SF3B1 mutation and other prognostic factors.
Based on the finding of higher risk of AML transformation and shorter survival in DNMT3A-mutated patients, as shown in current study, it would be interesting to investigate the effect of allogenic HSCT in these patients. We found that patients with DNMT3A mutations had a better OS if they received allogenic HSCT than those who did not. It implied that HSCT might ameliorate the poor survival impact of the adverse-risk genotype. Further prospective studies with more patients recruited are needed to verify this point. In a study of 46 decitabine-treated AML patients, Metzeler proposed that DNMT3A-mutated patients might have better treatment response and longer OS [45]. Subsequently, Traina et al. reported DNMT3A mutation as an independent predictor of better response and improved progression-free survival in MDS patients treated with DNMT inhibitors [41]. In our study, only 2 of 36 patients treated with HMA had DNMT3A mutation. These two patients had treatment response and OS similar to others. The influence of DNMT3A mutation on the treatment response to DNMT inhibitors was not evaluated because of the small number of DNMT3A-mutated patients.
DNMT3A mutation was found quite stable during disease evolution in AML patients [9, 46], but to the best of our knowledge, the dynamic change of this mutation in MDS patients has not been reported yet in literature. Here we showed that DNMT3A mutation was also quite stable in the clinical course of MDS patients; all DNMT3A-mutated patients retained the original mutations during sequential follow-ups unless CR was achieved after allogeneic HSCT or intensive chemotherapy. On the other hand, DNMT3A mutation was rarely acquired during disease evolution; only one (0.7%) of the 145 DNMT3A-wild patients acquired the mutation subsequently (Table 4).
It is well known that age-related clonal hematopoiesis is associated with increase in the risk of hematologic cancer and the majority of the variants occurred in three genes: DNMT3A, TET2, and ASXL1 [47,48,49]. Hematologic cancers were more common in persons with a variant allele fraction of 0.10 or greater. Therefore, it was proposed that DNMT3A mutation is relevant for initiating hematopoietic stem cell clonal expansion and an early initiation event for hematological malignancies. Our finding that DNMT3A mutation was retained unless CR was achieved was consistent with this hypothesis. In patients who failed to achieve remission, the clone harboring DNMT3A mutation survived and may contribute to subsequent relapse. Persistence of DNMT3A mutation in some AML patients in CR was described by us and other researchers [9, 50,51,52,53,54]. In a recent study of Gaidzik et al., DNMT3A mutant transcript levels in CR did not predict outcome in AML patients [54]. In contrast, Thol et al. showed that patients with DNMT3A-mutated lympho-myeloid clonal hematopoiesis (LM-CH) in CR had a higher cumulative incidence of relapse at 10 years compared with those without DNMT3A-mutated LM-CH (75 versus 27%) [55]. In the present study, we aimed to delineate the dynamic pattern of DNMT3A mutation in MDS development and progression. By NGS, the only patient (UPN 13) who retained his original DNMT3A mutation after high intensity chemotherapy finally relapsed. On the other hand, none of the patients in CR who lost their original DNMT3A mutation after allogeneic HSCT experienced disease relapse. Our data suggested that DNMT3A mutation might be used to assess the treatment response and the risk of relapse after curative-intent treatments in MDS patients. Together, whether retaining of DNMT3A mutations after curative-intent treatment is informative for the assessment of the relapse risk in MDS patients remains unclear. It should be cautious to interpret in clinical decision-making and more large-scale studies in MDS patients are warranted to clarify this point.
Conclusions
We identified associations of DNMT3A mutations with distinct clinical features and mutations of SF3B1 and IDH2 genes. In addition, we demonstrated that DNMT3A mutations independently predicted poor outcomes and were stable in the clinical course. It may be used as a biomarker to monitor the response after curative-intent treatment. Additional file 1, is available at Clinical Epigenetics’ website.
Abbreviations
- AML:
-
Acute myeloid leukemia
- BM:
-
Bone marrow
- CMML:
-
Chronic myelomonocytic leukemia
- CR:
-
Complete remission
- FAB:
-
French-American-British
- HMA:
-
Hypomethylating agents
- HSCT:
-
Hematopoietic stem cell transplantation
- IPSS:
-
International prognostic scoring system
- IPSS-R:
-
Revised international prognostic scoring system
- MDS:
-
Myelodysplastic syndrome
- MDS-EB1:
-
Myelodysplastic syndrome with excess blasts-1
- MDS-MLD:
-
Myelodysplastic syndrome with multilineage dysplasia
- NGS:
-
Next generation sequencing
- NTUH:
-
National Taiwan University Hospital
- OS:
-
Overall survival
- RA:
-
Refractory anemia
- RAEB:
-
Refractory anemia with excess blasts
- RAEB-T:
-
Refractory anemia with excess blasts in transformation
- RARS:
-
Refractory anemia with ring sideroblasts
- UPN:
-
Unique patient number
References
Jones PA, Laird PW. Cancer epigenetics comes of age. Nat Genet. 1999;21:163–7.
Seligson DB, Horvath S, Shi T, et al. Global histone modification patterns predict risk of prostate cancer recurrence. Nature. 2005;435:1262–6.
Razin A, Cedar H. DNA methylation and genomic imprinting. Cell. 1994;77:473–6.
Lee JT, Jaenisch R. The (epi)genetic control of mammalian X-chromosome inactivation. Curr Opin Genet Dev. 1997;7:274–80.
Okano M, Xie S, Li E. Cloning and characterization of a family of novel mammalian DNA (cytosine-5) methyltransferases. Nat Genet. 1998;19:219–20.
Yamashita Y, Yuan J, Suetake I, et al. Array-based genomic resequencing of human leukemia. Oncogene. 2010;29:3723–31.
Ley TJ, Ding L, Walter MJ, et al. DNMT3A mutations in acute myeloid leukemia. N Engl J Med. 2010;363:2424–33.
Thol F, Damm F, Ludeking A, et al. Incidence and prognostic influence of DNMT3A mutations in acute myeloid leukemia. Journal of clinical oncology : official journal of the American Society of Clinical Oncology. 2011;29:2889–96.
Hou HA, Kuo YY, Liu CY, et al. DNMT3A mutations in acute myeloid leukemia: stability during disease evolution and clinical implications. Blood. 2012;119:559–68.
Ribeiro AF, Pratcorona M, Erpelinck-Verschueren C, et al. Mutant DNMT3A: a marker of poor prognosis in acute myeloid leukemia. Blood. 2012;119:5824–31.
Shivarov V, Gueorguieva R, Stoimenov A, et al. DNMT3A mutation is a poor prognosis biomarker in AML: results of a meta-analysis of 4500 AML patients. Leuk Res. 2013;37:1445–50.
Walter MJ, Ding L, Shen D, et al. Recurrent DNMT3A mutations in patients with myelodysplastic syndromes. Leukemia. 2011;25:1153–8.
Thol F, Winschel C, Ludeking A, et al. Rare occurrence of DNMT3A mutations in myelodysplastic syndromes. Haematologica. 2011;96:1870–3.
Tefferi A, Vardiman JW. Myelodysplastic syndromes. N Engl J Med. 2009;361:1872–85.
Roller A, Grossmann V, Bacher U, et al. Landmark analysis of DNMT3A mutations in hematological malignancies. Leukemia. 2013;27:1573–8.
Chesnais V, Renneville A, Toma A, et al. Effect of lenalidomide treatment on clonal architecture of myelodysplastic syndromes without 5q deletion. Blood. 2016;127:749–60.
Bejar R, Stevenson KE, Caughey BA, et al. Validation of a prognostic model and the impact of mutations in patients with lower-risk myelodysplastic syndromes. Journal of clinical oncology: official journal of the American Society of Clinical Oncology. 2012;30:3376–82.
Bennett JM, Catovsky D, Daniel MT, et al. Proposals for the classification of the myelodysplastic syndromes. Br J Haematol. 1982;51:189–99.
Arber DA, Orazi A, Hasserjian R, et al. The 2016 revision to the World Health Organization classification of myeloid neoplasms and acute leukemia. Blood. 2016;127:2391–405.
Hou HA, Lin CC, Chou WC, et al. Integration of cytogenetic and molecular alterations in risk stratification of 318 patients with de novo non-M3 acute myeloid leukemia. Leukemia. 2014;28:50–8.
Hou HA, Kuo YY, Tang JL, et al. Clinical implications of the SETBP1 mutation in patients with primary myelodysplastic syndrome and its stability during disease progression. Am J Hematol. 2014;89:181–6.
Hou HA, Chou WC, Lin LI, et al. Characterization of acute myeloid leukemia with PTPN11 mutation: the mutation is closely associated with NPM1 mutation but inversely related to FLT3/ITD. Leukemia. 2008;22:1075–8.
Tang JL, Hou HA, Chen CY, et al. AML1/RUNX1 mutations in 470 adult patients with de novo acute myeloid leukemia: prognostic implication and interaction with other gene alterations. Blood. 2009;114:5352–61.
Hou HA, Liu CY, Kuo YY, et al. Splicing factor mutations predict poor prognosis in patients with de novo acute myeloid leukemia. Oncotarget. 2016;7:9084–101.
Shiah HS, Kuo YY, Tang JL, et al. Clinical and biological implications of partial tandem duplication of the MLL gene in acute myeloid leukemia without chromosomal abnormalities at 11q23. Leukemia. 2002;16:196–202.
Chen TC, Hou HA, Chou WC, et al. Dynamics of ASXL1 mutation and other associated genetic alterations during disease progression in patients with primary myelodysplastic syndrome. Blood cancer journal. 2014;e177:4.
Ernst T, Chase AJ, Score J, et al. Inactivating mutations of the histone methyltransferase gene EZH2 in myeloid disorders. Nat Genet. 2010;42:722–6.
Lin CC, Hou HA, Chou WC, et al. IDH mutations are closely associated with mutations of DNMT3A, ASXL1 and SRSF2 in patients with myelodysplastic syndromes and are stable during disease evolution. Am J Hematol. 2014;89:137–44.
Chou WC, Lei WC, Ko BS, et al. The prognostic impact and stability of Isocitrate dehydrogenase 2 mutation in adult patients with acute myeloid leukemia. Leukemia. 2011;25:246–53.
Chou WC, Chou SC, Liu CY, et al. TET2 mutation is an unfavorable prognostic factor in acute myeloid leukemia patients with intermediate-risk cytogenetics. Blood. 2011;118:3803–10.
Hou HA, Huang TC, Lin LI, et al. WT1 mutation in 470 adult patients with acute myeloid leukemia: stability during disease evolution and implication of its incorporation into a survival scoring system. Blood. 2010;115:5222–31.
Falini B, Mecucci C, Tiacci E, et al. Cytoplasmic nucleophosmin in acute myelogenous leukemia with a normal karyotype. N Engl J Med. 2005;352:254–66.
Hou HA, Chou WC, Kuo YY, et al. TP53 mutations in de novo acute myeloid leukemia patients: longitudinal follow-ups show the mutation is stable during disease evolution. Blood cancer journal. 2015;e331:5.
Hou HA, Tsai CH, Lin CC, Chou WC, Kuo YY, Liu CY, Tseng MH, Peng YL, Liu MC, Liu CW, Liao XW, Lin LI, Yao M, Tang JL, Tien HF. Incorporation of mutations in five genes in the revised international prognostic scoring system can improve risk stratification in the patients with myelodysplastic syndrome. Blood Cancer J. 2018.
Papaemmanuil E, Cazzola M, Boultwood J, et al. Somatic SF3B1 mutation in myelodysplasia with ring sideroblasts. N Engl J Med. 2011;365:1384–95.
Malcovati L, Karimi M, Papaemmanuil E, et al. SF3B1 mutation identifies a distinct subset of myelodysplastic syndrome with ring sideroblasts. Blood. 2015;126:233–41.
Greenberg PL, Tuechler H, Schanz J, et al. Revised international prognostic scoring system for myelodysplastic syndromes. Blood. 2012;120:2454–65.
Yang YT, Hou HA, Liu CY, et al. IPSS-R in 555 Taiwanese patients with primary MDS: integration of monosomal karyotype can better risk-stratify the patients. Am J Hematol. 2014;89:E142–9.
Kim SJ, Zhao H, Hardikar S, et al. A DNMT3A mutation common in AML exhibits dominant-negative effects in murine ES cells. Blood. 2013;122:4086–9.
Lin J, Yao DM, Qian J, et al. Recurrent DNMT3A R882 mutations in Chinese patients with acute myeloid leukemia and myelodysplastic syndrome. PLoS One. 2011;6:e26906.
Traina F, Visconte V, Elson P, et al. Impact of molecular mutations on treatment response to DNMT inhibitors in myelodysplasia and related neoplasms. Leukemia. 2014;28:78–87.
Tefferi A, Lasho TL, Patnaik MM, et al. Targeted next-generation sequencing in myelodysplastic syndromes and prognostic interaction between mutations and IPSS-R. Am J Hematol. 2017;92(12):1311–7.
Haferlach T, Nagata Y, Grossmann V, Okuno Y, Bacher U, Nagae G, Schnittger S, Sanada M, Kon A, Alpermann T, Yoshida K, Roller A, Nadarajah N, Shiraishi Y, Shiozawa Y, Chiba K, Tanaka H, Koeffler HP, Klein H-U, Dugas M, Aburatani H, Kohlmann A, Miyano S, Haferlach C, Kern W, Ogawa S. Landscape of genetic lesions in 944 patients with myelodysplastic syndromes. Leukemia. 2014;28(2):241–247.
Papaemmanuil E, Gerstung M, Malcovati L, et al. Clinical and biological implications of driver mutations in myelodysplastic syndromes. Blood. 2013;122:3616–27. quiz 3699
Metzeler KH, Walker A, Geyer S, et al. DNMT3A mutations and response to the hypomethylating agent decitabine in acute myeloid leukemia. Leukemia. 2012;26:1106–7.
Gaidzik VI, Weber D, Paschka P, et al. DNMT3A mutant transcript levels persist in remission and do not predict outcome in patients with acute myeloid leukemia. Leukemia. 2017;32:30–7.
Genovese G, Kahler AK, Handsaker RE, et al. Clonal hematopoiesis and blood-cancer risk inferred from blood DNA sequence. N Engl J Med. 2014;371:2477–87.
Xie M, Lu C, Wang J, et al. Age-related mutations associated with clonal hematopoietic expansion and malignancies. Nat Med. 2014;20:1472–8.
Jaiswal S, Fontanillas P, Flannick J, et al. Age-related clonal hematopoiesis associated with adverse outcomes. N Engl J Med. 2014;371:2488–98.
Ploen GG, Nederby L, Guldberg P, et al. Persistence of DNMT3A mutations at long-term remission in adult patients with AML. Br J Haematol. 2014;167:478–86.
Ivey A, Hills RK, Simpson MA, et al. Assessment of minimal residual disease in standard-risk AML. N Engl J Med. 2016;374:422–33.
Jeziskova I, Musilova M, Culen M, et al. Distribution of mutations in DNMT3A gene and the suitability of mutations in R882 codon for MRD monitoring in patients with AML. Int J Hematol. 2015;102:553–7.
Bhatnagar B, Eisfeld AK, Nicolet D, et al. Persistence of DNMT3A R882 mutations during remission does not adversely affect outcomes of patients with acute myeloid leukaemia. Br J Haematol. 2016;175:226–36.
Gaidzik VI, Weber D, Paschka P, et al. DNMT3A mutant transcript levels persist in remission and do not predict outcome in patients with acute myeloid leukemia. Leukemia. 2018;32:30–7.
Thol F, Klesse S, Kohler L, et al. Acute myeloid leukemia derived from lympho-myeloid clonal hematopoiesis. Leukemia. 2017;31:1286–95.
Acknowledgements
We would like to acknowledge the service provided by the DNA Sequencing Core of the First Core Laboratory, National Taiwan University College of Medicine.
Funding
This work was partially sponsored by grants MOST 103-2628-B-002-008-MY3, 103-2923-B-002-001, MOST 103-2314-B-002- 130-MY3, 103-2314-B-002-131 MY3, 104-2314-B-002-128-MY4, and 106- 2314-B-002-226-MY3 from the Ministry of Science and Technology (Taiwan), National Taiwan University Hospital−National Taiwan University joint research grant (UN103-051), and MOHW 105-TDU-B-211-134004 from the Ministry of Health and Welfare (Taiwan), NTUH 102P06, from the Department of Medical Research, National Taiwan University Hospital, and Taiwan Health Foundation.
Availability of data and materials
The datasets generated and/or analyzed during the current study are not publicly available due to individual privacy but are available from the corresponding author on reasonable request.
Author information
Authors and Affiliations
Contributions
M-EL was responsible for the data management and interpretation, mutation analysis, statistical analysis, and manuscript writing; H-AH was responsible for the study design, study plan and coordination, data management and interpretation, mutation analysis, statistical analysis, and manuscript writing; S-JW contributed patient samples and clinical data and was responsible for the data interpretation; C-HT and Y-YK were responsible for the mutation analysis and interpretation; J-LT, MY, C-CL, W-CC, S-YH, B-SK, S-CH, C-TL, and C-YC contributed patient samples and clinical data; M-HT, C-WL, and M-CL performed the gene mutation and chromosomal studies; H-FT designed and coordinated the study over the entire period and wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study was approved by the Institutional Review Board/Ethical Committee of the National Taiwan University Hospital (NTUH20150709RINA).
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional file
Additional file 1:
Table S1. The mutation patterns in 37 MDS patients with DNMT3A mutations at diagnosis. Table S2. Cytogenetics between MDS patients with and without DNMT3A mutation. Figure S1. Kaplan–Meier survival curves for overall survival among patients with DNMT3A mutations stratified by whether receiving allogeneic HSCT or not. Figure S2. Kaplan-Meier curves stratified by the status of DNMT3A mutations for overall survival among the 421 SF3B1-wild type MDS patients (A) and among the 48 SF3B1-mutated MDS patients (B). (DOC 242 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Lin, ME., Hou, HA., Tsai, CH. et al. Dynamics of DNMT3A mutation and prognostic relevance in patients with primary myelodysplastic syndrome. Clin Epigenet 10, 42 (2018). https://doi.org/10.1186/s13148-018-0476-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13148-018-0476-1