
Abstract
Background
Conventional targeted leishmanicidal chemotherapy has persistently remained prohibitive for most economically deprived communities due to costs, associated time to accessing health services and duration for successful treatment programme. Alternatives are bound to be incorporated in rational management of leishmaniasis by choice or default due to accessibility and cultural beliefs. Therefore, there is need to rigorously investigate and appraise the activity of medicinal compounds that may have anti-leishmanicidal activity especially in the context of products that are already being utilized by the populations for other ailments but have limited information on their therapeutic value and possible cytoxicity. Hence, the study examined both in vivo and in vitro response of L. major infection to Tephrosia vogelii extracts in BALB/c mice as the mouse model.
Methods
A comparative study design was applied for the in vivo and in vitro assays of the extract with Pentostam (GlaxoSmithKline, UK) and Amphotericin B [Fungizone™, X-Gen Pharmaceuticals (US)] as standard drugs.
Results
In BALB/c mice where the chemotherapeutic extract was administered intraperitoneally, there was significantly (p < 0.05) larger reduction in lesion size and optimal control of parasite burden than those treated orally. However, standard drugs showed better activity. Tephrosia vogelii had 50% inhibitory concentration (IC50) and IC90 of 12 and 68.5 μg/ml respectively, while the standard drugs had IC50 and IC90 of 5.5 and 18 μg/ml for Pentostam and 7.8 and 25.5 μg/ml for Amphotericin B in that order. In the amastigote assay, the infection rates decreased with increase in chemotherapeutic concentration. The multiplication indices for L. major amastigotes in macrophages treated with 200 µg/ml of the standard drugs and extract were significantly different (p < 0.05). 200 µg/ml of T. vogelii extract showed a multiplication index of 20.57, 5.65% for Amphotericin B and 9.56% for Pentostam. There was also significant difference (p < 0.05) in levels of Nitric oxide produced in the macrophages.
Conclusions
The findings demonstrated that T. vogelii extract has anti-leishmanial activity and further assays should be done to ascertain the active compounds responsible for anti-leishmanial activity.
Similar content being viewed by others


Background
The various forms of leishmaniases are considered neglected tropical diseases (NTDs). They affect majorly the economically disenfranchised populations in sub-Saharan Africa where, most of the affected do not seek and/or often synergize conventional medical attention with complementary options to alienate associated chronic suffering and opportunistic infections risks [1,2,3,4]. The observed trend has been linked to rational treatment and control challenges which include but not limited to prolonged period of time to fulfill standard chemotherapy regimen, associated costs, considerable level of side effects and recently, the emergence of antimony-resistant Leishmania strains in some endemic areas [4,5,6,7,8]. Equally, individually held knowledge by the traditional health practitioners (THP) has demonstrated a lot of complementary medicine diversity. This knowledge ranges from medicinal flora that can cure a variety of diseases with valuable clues for potentially anti-parasitic and protozoa compounds as well as current trends in terms of milestones, challenges and opportunities [3, 9,10,11].
Like most other settings globally, Kenya has a diversity of ethnicities and each has its own peculiar customs and beliefs and still rely on herbal remedies and trust the inherent potency value even when they can access modern medicine regardless of being rural or urban [12,13,14]. More than often, choosing to combine both herbal and modern medicine because of the belief they can target more than one complication at ago [13, 14]. Despite this understanding, most of the complementary medicines have not been documented and scientifically evaluated at fundamental assay level to determine their efficacy and dosage vis-à-vis the alleged diverse synergistic benefit of acting on a wide range of possible infections as often advanced by the Traditional health practitioners and communal history that vary by ethnicities [4, 14,15,16].
Various species from the family Fabaceae have demonstrated a diversity of applicability as chemotheraputants. Extracts of the leaf, bark, root, seed and flower (with literature suggesting predominance focus on the leaves) have demonstrated anti-leishmanial, antimicrobial, anthelmintic, antiprotozoal, analgesic, antioxidant and anti-inflammatory activity when tested in vitro and/or in vivo in different animal and murine models [17,18,19,20,21,22] while vector control worth and mitigation of various intermediate hosts (e.g. larvicidal activity against Aedesaegypti, Acaricidal activity on ticks, Biomphalariapfeifferi and Bulinustruncatus) have also been reported widely [20, 23,24,25]. Tephrosia purpurea (family: Fabaceae), commonly used in traditional remedies for the treatment of febrile attacks, enlargement and obstruction of liver, spleen, and kidney, was found to have significant antileishmanial activity, and has been evaluated extensively in various experimental animal models to locate the abode of activity [22] with near nil evidence from Kenya on the same or related family and species. The leguminous Tephrosia vogelii from the same family Fabaceae and class Magnoliopsida (dicotyledons), subclass Rosidae order Fabales, has been and is being used extensively in Kenya to improve soil fertility and as a medicinal product against various antimicrobial, protozoa and parasitic infections [16, 21, 26, 27]. Preferential and selective validation of the medicinal properties of the plant has resulted to progressively limited understanding on anti-protozoa capacity of the plant and as to whether the leguminous extracts would also be having anti-leishmanicidal activity with the understanding of the fact that leishmaniases are caused by obligate intracellular kinetoplastid protozoa of the genus Leishmania. Equally, beyond the scope of this study, the in vitro investigation enhances the availability of information on possible extent of the extract cytoxicity and safety in the Kenyan context where the information is sparse. In consideration of the ascribed available circumstantial animal and murine experimental evidence, gaps and envisaged information benefit in the context of the tropical forested ecological and ethno botanical diversity for alternative medicinal compounds indepth understanding and precisely Tephrosia vogelii [4, 7, 10, 28, 29], the aim of this study was to assess the response of Leishmania major to Tephrosia vogelii therapy in BALB/c mice (in vivo) and also in vitro in a quest to shape and consolidate baseline evidence on cytotoxicity and activity in the home-grown context and pave way for indepth validation.
Methods
Experimental setup
The in vivo and in vitro studies were carried out using a comparative study design. The efficacy and toxicity of the sample was compared with those of Pentostam (GlaxoSmithKline, UK) and Amphotericin B [Fungizone™, X-Gen Pharmaceuticals (US)]. Rosewell Park Memorial Institute 1640 Medium (RPMI) was used as control in in vitro experimental chemotherapeutic studies. In vivo studies were further subjected to a complete randomized block design. Phosphate Buffered Saline was used as the control. The results were compared to determine the efficacy of the test sample against the known standard drugs for treating leishmaniasis.
Plant collection and extract preparation
Identified and dried T. vogelii leaves were obtained from Kenya Medical Research Institute (KEMRI) herbarium, Nairobi. Extraction was done in KEMRI, Nairobi Kenya at the Centre for Traditional Medicine and Drug Research (CTMDR). The dried leaves were processed (to obtain the extract) as earlier described by Kigondu et al. [28]. The dry leaves were chopped into small pieces and then ground into powder form using a laboratory blender. 1 kg of each powder was soaked in absolute methanol for 3 days to extract compounds. The extract was filtered, dried with Na2SO4 and the solvent removed under vacuum in a rotary evaporator at 30–35 °C and stored at 20 °C until required for use. This methanolic extracts were used for anti-leishmanial tests.
Mice, Leishmania parasites and experimental infections
The BALB/c female mice aged 8 weeks and weighing 20 ± 2 g used for the study were obtained from the KEMRI’s animal house facility in Nairobi. The animals were moved into the experimental room for acclimatization 1 week before the start of the experiments. The mice were housed in 15 cm × 21 cm × 29 cm transparent cages. They were fed with pellets (Mice pellets UNGA® feeds) and water ad libitum.
Leishmania major (strain IDUB/KE/83=NLB-144) which was originally isolated in 1983 from a female Phlebotomus dubosqi collected near marigat, Baringo County in Kenya were used [30]. These parasites were cultivated in schneider’s Insect Medium supplemented with 20% heat inactivated foetal bovine serum, 100 µg/ml penicillin and 50 µg/ml streptomycin and 250 µg/ml 5-fluorocytosine arabinioside [31, 32]. This strain has been maintained by cryopreservation and in vitro culture and periodic passage in BALB/c mice at KEMRI, Nairobi. Promastigotes were incubated at 25 °C grown to stationary phase to generate infective metacyclic forms at 6th day of culture. Promastigotes in the medium were counted with a hemocytometer (Improved Double Neubauer) (Pharmacia-GE Healthcare, Uppsala, Sweden) with a Nikon optiphot optical microscope at 40× magnification.
In vitro studies
Cytotoxicity assays
Vero cells were cultured and maintained in Minimum Essential Medium (MEM) supplemented with 10% Foetal Bovine Serum (FBS). The cells were cultured at 37 °C in 55% CO2 for 24 h, harvested by trypsinization, pooled in a 50 ml vial and 100 µl cell suspension (1 × 106 cells/ml) put into 2 wells of rows A-H in a 96-well micro titer plate, the medium aspirated off and 150 µl of the highest concentration (1000 µg/ml) of the T. vogelii added into the first row and serial dilution carried out. The experimental plates were incubated further at 37 °C for 48 h. The controls used were cells with no extract but medium alone. MTT reagent (10 µl) was added into each well and the cells incubated for 4 h until a purple precipitate clearly visible under a microscope was formed. The medium together with MTT were aspirated off from the wells. Dimethylsulfoxide (DMSO) (100 µl) was added and the plates shaken for 5 min. The absorbance was measured for each well at 562 nm using a micro-titre plate reader [33].
Evaluation of IC50 concentration and anti-promastigote assay
Leishmania major promastigotes (106 parasites/ml) were incubated at 26 °C for 120 h in fresh media (brain heart infusion medium), Supplemented with 10% FBS in the absence or presence of serial concentrations (100, 50, 25, 12.5, 6.25 and 3.125 µg/ml) of the extracts (cell growth was determined daily by assessment of visible turbidity). The IC50 was considered as the concentration that inhibited 50% of L. major growth in vitro. Leishmania major promastigotes were incubated in 24-well plates in the presence of different concentrations of the extracts added. After 5 days of cultivation, aliquots of parasites were transferred to a 96-well micro-titre plate. The parasites were then incubated at 27 °C in 5% CO2 for 24 h; 200 µl of highest concentration of extract was added and diluted. The experimental plates were incubated further at 27 °C for 48 h. The controls used were promastigotes with no extracts and medium alone. In each well, 100 µl of DMSO was added and the plates shaken for 5 min. Absorbance was measured for each well at 562 nm using a micro titre reader [34].
Anti-amastigote assay
Peritoneal macrophages were harvested from 8 week old female BALB/c mice using Phosphate Buffered Saline 4 days after intraperitoneal injection at the Kenya Medical Research Institute (KEMRI) animal breeding facility, Nairobi-Kenya. After harvesting, the macrophages were processed as earlier described [35]. The obtained macrophages were then infected with L. major stationary phase promastigotes at a 6:1 parasite/macrophage ratio. The infected macrophages were incubated at 37 °C in 5% CO2 for 4 h. After incubation the remaining parasites were washed off using PBS and the cultures incubated in RPMI for 24 h. Treatment of infected macrophages with the experimental chemotheraputants was done once. Pentostam and Amphotericin B were used as standard treatment control drugs for comparison of parasite inhibition. The medium and drug were replenished daily for 3 days. On the 4th day, the monolayers were washed with PBS at 37 °C, fixed in methanol and stained with Giemsa. The number of amastigotes was determined by counting at least 100 macrophages in duplicate cultures, and the results expressed as infection rate (IR) and multiplication index (MI) as well as IC90 and IC50 with varied concentration rates of test drugs being 25, 50, 100 and 200 µg/ml [36,37,38,39].
Nitric oxide production assay
Nitric oxide release in macrophage cultures was measured using the Griess reaction for nitrites [40]. Supernatants were collected 48 h after introducing the test drug into the culture medium (in the amastigote assay) and stored at − 70 °C for analysis of nitric oxide (NO) production. Nitrite (NO2) accumulation in the cell culture supernatants was used as an indicator of nitric oxide production and it was determined by a standard Griess reaction [41,42,43].
In vivo studies
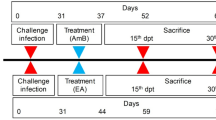
Mice and parasite inoculation, quantifying parasite burden from spleen and liver
BALB/c mice were inoculated with 1 × 106 stationary phase L. major promastigotes in 50 μl phosphate buffered saline into the Left Hind Footpad (LHFP) using a 29 gauge needle and left for 4 weeks incubation period [41, 44]. The inoculated mice were then randomly assigned into 5 groups of 8 mice in each. Group 1 was treated with T. vogelii extracts orally (0.2 mg/ml [200 µg/ml]), group 2 with T. vogelii extracts intraperitoneally (0.2 mg/ml [200 µg/ml]), group 3 with Pentostam intraperitoneally, group 4 with Amphotericin B intraperitoneally and group 5 with sterile PBS intraperitoneally at 100 µg/ml in the order for the latter three while PBS was used as the solvent-treatment commenced at the start of the 5th week post infection (day 29), for 28 days [44,45,46,47]. Lesion sizes (the difference in thickness between the infected footpad and the non-infected contralateral Footpad) were monitored weekly up to week 10; by measurement using a Starret dial caliper (Mitutoyo, Suzano, SP, and Brazil) [44, 48, 49]. The weight of the mice was also monitored on a weekly basis. A total of four randomly selected mice per group were sampled at week 10 post infection and chemotherapy for analysis of L. major parasite loads. The mice were anesthetized intraperitoneally with Pentobarbital sodium 80 mg/kg (Rompun; Bayer Plc., Newbury, UK). The liver and spleen were removed and weighed and changes post-infection and chemotherapy were determined based spleno-somatic and hepato-somatic indices [44]. Splenic and liver L. major burdens were determined from Giemsa-stained impression smears and expressed as Leishman-Donovan units (the number of amastigotes per 1000 host nuclei, multiplied by the weight of the organ) [43, 44].
Data analysis
The data collected on lesion sizes, parasite loads and absorbance were analyzed using the SPSS software. All experiments were performed in triplicate. The mean and standard errors of the lesions of each treatment group were compared using students’ t test and analysis of variance (ANOVA). Students’ t test was used in the analysis of differences in means obtained for the experimental groups. Chi square test was used in the analysis of infection rates and multiplication indices. p values of equal or less than 0.05 (p ≤ 0.05) were considered significant.
Results
Cytotoxicity assay
Results indicating the cell viability of vero cells subjected to the test compounds are shown in Fig. 1. The safety of the test extract was evaluated by treating vero cells with the extracts and viability was determined. Crude extracts of T. vogelii caused no significant adverse effects on the vero cells. Concentration of the standard drugs and test extract required to destroy 50% of the mammalian cell was significantly low in Amphotericin B (200 µg/ml), followed by Pentostam (250 µg/ml) and T. vogelii (1200 µg/ml). RPMI served as the control hence did not have any effect on the vero cells. Minimum inhibitory concentration for all the test drugs did not result in significant vero cell destruction. For Amphotericin B 1% of the cells were destroyed with 10 µg/ml concentration, Pentostam 1% were destroyed with 12 µg/ml treatment while for T. vogelii 1% were destroyed with 25 µg/ml treatment.

Cell viability of the vero cells subjected to the different test drugs
Efficacy of T. vogelii crude extracts on promastigotes of L. major
The promastigote growth inhibition was significantly affected by the various experimental drugs (p < 0.05) after 24 h of exposure. The optimal efficacy of the standard drugs was 97.5 and 98.9% for Amphotericin B and Pentostam respectively. Tephrosia vogelii achieved an optimal efficacy of 91.4% against promastigotes. There was a significant difference in the IC50 and IC90 of the test compounds (p < 0.05). The standard drugs were more effective against promastigote as compared to T. vogelii with Pentostam having an IC50 of 5.5 μg/ml, Amphotericin B with 7.8 μg/ml while that of T. vogelii being 12 μg/ml. There was significant (p < 0.05) difference in the IC90 with the lowest IC90 occurring in Pentostam followed by Amphotericin B then by T. vogelii with 18, 25.5 and 64 μg/ml respectively. Table 1 describes the optimal efficacy, IC90 and IC50 of the test drugs against promastigote forms of the parasites.
Efficacy of T. vogelii extract on amastigotes of L. major
To test the efficacy of the experimental drugs against L. major amastigotes, the infection rates (IR) and multiplication indices (MI) were determined as shown in Fig. 2. There was a general decrease in infection rate with increase in concentration of the treatments applied across the groups with significant difference between T. vogelii and the standard drugs (p = 0.01) whereas, infection of macrophages was not significantly affected when treated with Pentostam and Amphotericin B (p = 0.32). At the least concentration of 25 μg/ml, the macrophages were infected by amastigotes at 80% with T. vogelii treatment. At the same concentration Pentostam and Amphotericin B had 22 and 18% infection rates respectively. The infection rates reduced with increase in concentration with Amphotericin B and Pentostam falling from 18 to 2 and 22 to 3% respectively. The same trend was observed in T. vogelii from 80 to 20% when the concentration was increased to 200 µg/ml.

The infection rates of amastigotes in macrophages following treatments with various test drugs
The Multiplication indices for L. major amastigotes in macrophages treated with 200 µg/ml of the test drugs were significantly different (p < 0.05); T. vogelii extract treatment showed a multiplication index of 20.57%, 5.65% for Amphotericin B and 9.56% for Pentostam. At a lower concentration of 25 µg/ml L. major amastigotes showed multiplication indices of 75.86% in T. vogelii extract, 47.5% in Amphotericin B and 51.07% in Pentostam. The multiplication indices are as shown in Fig. 3.

Multiplication indices (growth rates) of amastigotes in macrophages after treatment with the different test drugs
The optimal efficacy, IC90 and IC50 of the test drugs against amastigote forms of the parasites were significantly inhibited by the various test compounds (Table 2 describes). There were significant differences in the optimal efficacy of the test drugs (p = 0.0001). The optimal efficacy of the standard drugs was 91 and 98% for Amphotericin B and Pentostam respectively. Tephrosia vogelii achieved an optimal efficacy of 88.5% against amastigotes. There was a significant difference in the IC50 and IC90 of the test compounds (p < 0.05). The standard drugs were more effective against amastigote as compared to T. vogelii with Pentostam having an IC50 of 6.8 μg/ml, Amphotericin B with 5.8 μg/ml while that of T. vogelii being 9.2 μg/ml. There was significant (p = 0.001) difference in the IC90 with the lowest IC90 occurring in Pentostam followed by Amphotericin B then by T. vogelii with 16.8, 24.5 and 58.5 μg/ml respectively.
Effect of T. vogelii crude extracts on L. major lesion development in BALB/c mice
The lesion sizes of BALB/c mice at the start of infection and in the course of treatment with the different test drugs are as shown in Fig. 4. There were no significant differences in development of lesion sizes in BALB/c mice during the first 4 weeks post-infection with L. major (p > 0.05). Between week 5 and 10, the differences in lesion sizes progression were subjected to repeated measure ANOVA, which indicated that there were significant differences in lesion sizes among different treatment groups (p = 0.004). The lesion sizes of the untreated control BALB/c mice increased steadily after infection. Smallest lesion sizes occurred in BALB/c mice treated with amphotericin B, which was slightly lower than BALB/c mice treated with Pentostam. BALB/c mice treated with T. vogelii intraperitoneally resulted insignificantly (p < 0.05) larger reduction of lesion sizes than the BALB/c mice treated orally with T. vogelii.

Lesion sizes of BALB/c mice at the start of infection, at the start of treatment and during the treatment with T. vogelii, PBS, Pentostam and Amphotericin B
Body weights, organs weight, organo-somatic indices and parasite loads in spleen and liver of BALB/c mice treated with different test drugs
There were significant differences in the weight of spleen, liver, spleno-somatic and hepato-somatic indices among treatment groups (p < 0.05). The untreated control group had significantly large spleen and liver weights alongside spleno-somatic and hepato-somatic indices accounting for the spleno and hepatomegaly observed. Pentostam and Amphotericin B treated groups had no significant difference between them producing the least spleen and liver weights and respective spleno-somatic and hepato-somatic indices. Tephrosia vogelii treatment produced an intermediate between the untreated control and the standard drugs treated groups. However, intraperitoneal administration of the test extract resulted in lower spleen/liver weights and organo-somatic indices than the oral administration mode (p < 0.05). Body weights, weight of spleen, liver and organo-somatic indices in BALB/c infected with L. major infection under various treatments are as shown in Table 3.
The Leishman Donovan units (LDU) in the spleens of BALB/c mice subjected to oral and intraperitoneal administration of T. vogelii were significantly different (p = 0.0014). Treatment with T. vogelii intraperitoneally resulted in lower LDU (3.21 × 106 than those observed when T. vogelii was administered orally 5.05 × 106. Infected BALB/c mice treated with Amphotericin B and Pentostam resulted in the lowest LDU; 0.24 × 106 and 0.45 × 106 respectively but there was no significant difference between Pentostam and Amphotericin B treated groups. The LDU of L. major parasites in the spleens of BALB/c mice infected with L. major receiving T. vogelii, Amphotericin B, Pentostam and PBS are shown in Fig. 5.

L. major LDU in spleens of BALB/c mice treated with different test drugs
Intraperitoneal and oral administration of T. vogelii produced significantly different LDU trends (p = 0.002) in the liver. Pentostam and Amphotericin B resulted in significantly the lowest LDU; 0.18 × 106 and 0.32 × 106 respectively. However, there was no significant difference between Pentostam and Amphotericin B (p > 0.05) treated groups. The LDU of L. major parasite in the liver of BALB/c mice infected with L. major receiving T. vogelii, Amphotericin B, Pentostam and PBS treatments is shown in Fig. 6.

L. major LDU in liver of BALB/c mice treated with different test drugs
Nitric oxide production in treated L. major-infected macrophages treated with different test drugs
There was significant difference in nitric oxide production in macrophages of BALB/c mice infected with L. major amastigotes and subjected to the different test drugs (p < 0.05). RPMI treatment produced the highest nitric oxide maintaining a consistent trend in all the concentrations. Amphotericin B treatment produced the lowest nitric oxide followed by Pentostam however, there was no significant difference between the two (p > 0.05). Tephrosia vogelii treatment produced higher amounts of nitric oxide compared to the standard drugs treated groups. Nitric oxide production in the macrophages of L. major infected BALB/c mice and treated with different test drugs is as shown in Fig. 7.

Nitric oxide production in the macrophages of L. major infected BALB/c mice and treated with different test drugs
Discussion
Despite existence of substantive chemotheraputants for different forms of leishmaniases there is also increasing evidence of parasite resistance to the conventional anti-parasitic therapies coupled with the prevailing traditional challenges of in accessibility, side effects and costs [50]. Moreover, vaccines against Leishmania species are still under development hence no vaccine exist yet [8]. Therefore, there is a need to search for alternatives and progressive validation of the same from fundamental research level to large scale trials where possible. Studies on medicinal plants and their chemotherapeutic activities against Leishmania species have been on the rise. These plants have demonstrated effectiveness against the parasite and making a contribution as alternative therapies [7]. In varying contexts, the benefit of scaling up chemotherapeutic options including using plant based products has been associated with the capacity to ensure decreased development of resistance by parasites [11, 51].
This study was carried out for the first time in the context to investigate activity of T. vogelii against L. major. The study was set to determine both the in vitro and the in vivo experimental outcomes. In the in vitro component, the cytotoxicity assay was set up to determine the toxicity levels of the test extract on vero cells. The cytotoxicity assay on the crude extracts of T. vogelii caused no significant adverse effects on the vero cells; this is further reflected in BALB/c mice peritoneal macrophages in vitro in the amastigote assay. The non-toxicity of T. vogelii is also reflected in the in vivo treatment using the extract, in which no mice died and none had a marked adverse degeneration. There was no marked change on weight, the appearance of the skin of the mice among other observable factors linked to physiological mechanism hence the plant extract was well tolerated. Compared to Amphotericin B and Pentostam, T. vogelii had less toxicity to vero cells and can lead to novel treatment for L. major infection.
Tephrosia vogelii extracts demonstrated remarkable in vitro anti-leishmanial activity against L. major promastigotes and amastigotes. In both assays, the activity of T. vogelii extracts against L. major showed that the plants contained some pharmacologically active substances that could prevent growth and proliferation of L. major promastigotes and amastigotes in the macrophage [52,53,54,55,56,57].
Leishmania parasite evades the host’s immune system by transforming from the flagellated promastigote to non-flagellated amastigote in macrophages of the mammalian host. This stage is critical for establishment of the infection [58]. This therefore needs a drug that will target the parasite in the amastigote form while inside the macrophages. A reduction in infection rates was evident following treatment with T. vogelii which is an indication that the extract suppressed either the establishment or the development process of L. major amastigotes in BALB/c peritoneal macrophages. The multiplication indices imply that the higher the concentrations the higher the inhibition rate of replication of amastigotes in the macrophages. The presented findings also indicated that T. vogelii extract caused marked reduction in splenomegaly and hepatomegaly (the parasite loads in the spleen and liver) of the mice during treatment period and reduced lesion development; suggesting that the extract indeed inhibited the growth of L. major parasite. T. vogelii given intraperitoneally gave better results than when given orally. This is due to the fact that intraperitoneal treatments usually act faster as there is enhanced uptake for the medicinal components than those given orally. However, the activity of the extract was lower than that of the standard drugs. This would possibly be attributed to the fact that T. vogelii was used in its crude form and isolation of active ingredients may improve its potency [17, 18].
Nitric Oxide is known to mediate macrophage cytotoxicity against microbes and tumor cells. It is usually upregulated in macrophages during L. major infection [57]. Apart from the normal nitric oxide produced by Leishmania infected macrophages naturally, the crude extract stimulated the macrophages in vitro to produce more nitric oxide for activity against the parasite indirectly. T. vogelii caused relatively higher induction of nitric oxide compared to the controls. This suggests that the plant extract employ the mechanism of nitric oxide cytolytic activity to eliminate the amastigotes which could be due to the different compounds present in T. vogelii extract.
There are some limitations associated with the presented finding. First, the in vivo treatment majorly focused on oral and intraperitoneal mode of administration hence other modes of treatment that is intravenous and subcutaneous should also be explored. To confirm on the non-toxic nature of the plant extracts, the effect that various factors such as the growth stage and maturity of the plant, regional variations (where the plant is growing) should be looked into. Leishmaniasis associated immunomodulatory markers were not analyzed in vivo. The evaluation can help give more insight on the validity of the extracts.
Conclusions
Based on the findings of this study, T. vogelii extract exhibited anti-leishmanial activity with inhibitory action in both promastigotes and amastigotes with the activity and toxicity levels being compared to the standard drugs. The plant extract had significant activity when administered orally, hence suggesting can be administered both orally and intraperitoneally unlike the standard drugs that can only be administered through a specified mode. It was interesting to note that, the plant extract seemed to act in synergy with immunomodulation to wade the parasitic infection intracellular as observed through increased production of nitric oxide by the stimulated macrophages. Further studies should be done on isolating and testing individual active compounds which may give better anti-leishmanial activity and limit toxicity.
Abbreviations
- IC:
-
inhibitory concentration
- NTDs:
-
neglected tropical diseases
- THP:
-
Traditional Health Practitioners
- RPMI:
-
Rosewell Park Memorial Institute 1640 Medium
- KEMRI:
-
Kenya Medical Research Institute
- CTMRD:
-
Centre for Traditional Medicine and Drug Research
- FBS:
-
Foetal Bovine Serum
- MEM:
-
Minimum Essential Medium
- MTT:
-
3-4,5-dimethgylthiaol-2-yl-2, 5-diphenyltetrazolium bromide
- DMSO:
-
dimethylsulfoxide
- IR:
-
infection rate
- MI:
-
multiplication index
- NO:
-
nitric oxide
- NO2 :
-
nitrite
- LHFP:
-
left hind footpad
- PBS:
-
phosphate buffered saline
- LDU:
-
Leishman-Donovan units
- ANOVA:
-
analysis of variance
References
Organization WH. Global tuberculosis control: WHO report 2010. World Health Organization; 2010.
Alvar J, Velez ID, Bern C, Herrero M, Desjeux P, Cano J, Jannin J, den Boer M, Team WLC. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE. 2012;7(5):e35671.
Monzote L. Current treatment of leishmaniasis: a review. Open Antimicrob Agents J. 2009;1:1.
Organization WH. WHO traditional medicine strategy 2002–2005; 2002.
Bryceson A. A policy for leishmaniasis with respect to the prevention and control of drug resistance. Trop Med Int Health. 2001;6(11):928–34.
Tinui W. Situational analysis of leishmaniases research in Kenya. Afr J Health Sci. 2008;13(1):7–21.
Croft SL, Coombs GH. Leishmaniasis–current chemotherapy and recent advances in the search for novel drugs. Trends Parasitol. 2003;19(11):502–8.
Handman E. Leishmaniasis: current status of vaccine development. Clin Microbiol Rev. 2001;14(2):229–43.
Fournet A, Barrios AA, Munoz V, Hocquemiller R, Cave A. Effect of natural naphthoquinones in BALB/c mice infected with Leishmania amazonensis and L. venezuelensis. Trop Med Parasitol. 1992;43(4):219–22.
Laban LT, Anjili CO, Mutiso JM, Ingonga J, Kiige SG, Ngedzo MM, Gicheru MM. Experimental therapeutic studies of Solanum aculeastrum Dunal. on Leishmania major infection in BALB/c mice. Iran J Basic Med Sci. 2015;18(1):64.
Ridley RG, Fletcher ER. Making a difference: 30 years of TDR. Nat Rev Microbiol. 2008;6(5):401–7.
Matheka DM, Demaio AR. Complementary and alternative medicine use among diabetic patients in Africa: a Kenyan perspective. Pan Afr Med J. 2014;15(1):110.
Mothupi MC. Use of herbal medicine during pregnancy among women with access to public healthcare in Nairobi, Kenya: a cross-sectional survey. BMC Complement Altern Med. 2014;14(1):1.
Kigen GK, Ronoh HK, Kipkore WK, Rotich JK. Current trends of traditional herbal medicine practice in Kenya: a review. Afr J Pharmacol Ther. 2013;2(1):32–7.
Maina N, Kagira J, Achila O, Karanja S, Ngotho M. Herbal medicines in Kenya: a review of the toxicity and quality control issues. Afr J Health Sci. 2013;24:29–47. http://www.ajhsjournal.or.ke/?p=801.
Lambert J, Leonard K, Mungai G, Omindi-Ogaja E, Gatheru G, Mirangi T, Owara J, Herbst CH, Ramana GNV, Lemiere C. The contribution of traditional herbal medicine practitioners to Kenyan health care delivery: results from community health-seeking behavior vignettes and a traditional herbal medicine practitioner survey. Health, Nutrition and Population (HNP) discussion paper. Washington, DC: World Bank; 2011. http://documents.worldbank.org/curated/en/620571468285024188/The-contribution-of-traditional-herbal-medicine-practitioners-to-Kenyan-health-care-delivery-results-from-community-health-seeking-behavior-vignettes-and-a-traditional-herbal-medicine-practitioner-survey.
Touqeer S, Saeed MA, Ajaib M. A review on the phytochemistry and pharmacology of genus Tephrosia. Phytopharmacology. 2013;4(3):598–637.
Wanga B, Akenga T, Imbuga M, Gitonga L, Olubayo F, Namungu P. Antimicrobial acitivity of extracts from Tephrosia vogelii Hook F. J Agric Sci Technol. 2007;8(1):1–14.
Adaudi A, Aluwong T, Salawu O, Anuka J. Blood pressure, analgesic and anti-inflammatory properties of methanolic extracts of Tephrosia vogelii in experimental animals. Niger Vet J. 2009;30(3):37–43.
Ganapaty S, Pannakal ST, Srilakshmi GV, Lakshmi P, Waterman PG, Brun R. Pumilanol, an antiprotozoal isoflavanol from Tephrosia pumila. Phytochem Lett. 2008;1(4):175–8.
Ganapaty S, Srilakshmi G, Thomas PS, Rajarajeshwari N, Ramakrishna S. Cytotoxicity and antiprotozoal activity of flavonoids from three Tephrosia species. J Nat Rem. 2009;9(2):202–8.
Sharma P, Rastogi S, Bhatnagar S, Srivastava JK, Dube A, Guru PY, Kulshrestha DK, Mehrotra BN, Dhawan BN. Antileishmanial action of Tephrosia purpurea linn, extract and its fractions against experimental visceral leishmaniasis. Drug Dev Res. 2003;60(4):285–93.
Matovu H, Olila D. Larvicidal activity of Tephrosia vogelii crude extracts on mosquito larval stages. Res J Biol Sci. 2007;2(6):612–6.
Kalume M, Losson B, Angenot L, Tits M, Wauters J, Frédérich M, Saegerman C. Rotenoid content and in vitro acaricidal activity of Tephrosia vogelii leaf extract on the tick Rhipicephalus appendiculatus. Vet Parasitol. 2012;190(1):204–9.
Jacques DT, Safiou A, Jédirfort H, Souaïbou F. In vitro effect of the ethanolic extract of Tephrosia vogelii on Rhipicephalus sanguineus in Abomey-Calavi. Avicenna J Phytomed. 2015;5(3):247.
Odhong C, Wahome R, Vaarst M, Nalubwama S, Halberg N, Githigia S. In vitro anthelmintic effects of crude aqueous extracts of Tephrosia vogelii, Tephrosia villosa and Carica papaya leaves and seeds. Afr J Biotechnol. 2015;13(52):4667–72.
Rutunga V, Karanja NK, Gachene CK, Palm C. Biomass production and nutrient accumulation by Tephrosia vogelii (Hemsley) A. Gray and Tithonia diversifolia Hook F. fallows during the six-month growth period at Maseno, Western Kenya. Biotechnol Agronomie, Société et Environnement. 1999;3(4):237–2460.
Kigondu EV, Rukunga GM, Keriko JM, Tonui WK, Gathirwa JW, Kirira PG, Irungu B, Ingonga JM, Ndiege IO. Anti-parasitic activity and cytotoxicity of selected medicinal plants from Kenya. J Ethnopharmacol. 2009;123(3):504–9.
Bent S, Ko R. Commonly used herbal medicines in the United States: a review. Am J Med. 2004;116(7):478–85.
Beach R, Kiilu G, Hendricks L, Oster C, Leeuwenburg J. Cutaneous leishmaniasis in Kenya: transmission of Leishmania major to man by the bite of a naturally infected Phlebotomus duboscqi. Trans R Soc Trop Med Hyg. 1984;78(6):747–51.
Hendricks L, Wright N. Diagnosis of cutaneous leishmaniasis by in vitro cultivation of saline aspirates in Schneider’s Drosophila Medium. Am J Trop Med Hyg. 1979;28(6):962–4.
Kimber C, Evans D, Robinson B, Peters W. Control of yeast contamination with 5-fluorocytosine in the in vitro cultivation of Leishmania spp. Ann Trop Med Parasitol. 1981;75(4):453.
Wang Y, Yuan B, Deng X, He L, Wang S, Zhang Y, Han Q. Comparison of alpha1-adrenergic receptor cell-membrane stationary phases prepared from expressed cell line and from rabbit hepatocytes. Anal Bioanal Chem. 2006;386(7–8):2003–11.
Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods. 1983;65(1–2):55–63.
Delorenzi JC, Attias M, Gattass CR, Andrade M, Rezende C, da Cunha Pinto Â, Henriques AT, Bou-Habib DC, Saraiva EM. Antileishmanial activity of an indole alkaloid from Peschiera australis. Antimicrob Agents Chemother. 2001;45(5):1349–54.
Berman JD, Lee LS. Activity of antileishmanial agents against amastigotes in human monocyte-derived macrophages and in mouse peritoneal macrophages. J Parasitol. 1984;70:220–5.
Makwali JA, Wanjala FM, Kaburi JC, Ingonga J, Byrum WW, Anjili CO. Combination and monotherapy of Leishmania major infection in BALB/c mice using plant extracts and herbicides. J Vector Borne Dis. 2012;49(3):123.
Nigussie D, Tasew G, Makonnen E, Debella A, Hurrisa B. In-vitro investigation of fractionated extracts of Albizia gummifera seed against Leishmania donovani amastigote stage. J Clin Cell Immunol. 2015;6(373):2.
Ogeto T, Odhiambo R, Shivairo R, Muleke C, Osero B, Anjili C, Ingonga J, Osuga I. Antileishmanial activity of Aloe secundiflora plant extracts against Leishmania major. Adv Life Sci Tech. 2013;13:9–18.
Holzmuller P, Sereno D, Cavaleyra M, Mangot I, Daulouede S, Vincendeau P, Lemesre J-L. Nitric oxide-mediated proteasome-dependent oligonucleosomal DNA fragmentation in Leishmania amazonensis amastigotes. Infect Immun. 2002;70(7):3727–35.
Vodovotz Y, Bogdan C, Paik J, Xie Q, Nathan C. Mechanisms of suppression of macrophage nitric oxide release by transforming growth factor beta. J Exp Med. 1993;178(2):605–13.
Bogdan C, Vodovotz Y, Paik J, Xie Q, Nathan C. Mechanism of suppression of nitric oxide synthase expression by interleukin-4 in primary mouse macrophages. J Leukoc Biol. 1994;55(2):227–33.
Bradley D, Kirkley J. Regulation of Leishmania populations within the host. I. the variable course of Leishmania donovani infections in mice. Clin Exp Immunol. 1977;30(1):119.
Khayeka-Wandabwa C, Kutima H, Nyambati V, Ingonga J, Oyoo-Okoth E, Karani L, Jumba B, Githuku K, Anjili CO. Combination therapy using Pentostam and Praziquantel improves lesion healing and parasite resolution in BALB/c mice co-infected with Leishmania major and Schistosoma mansoni. Parasit Vectors. 2013;6:244.
Okech B, Irungu LW, Anjili C, Munyua J, Njagi E, Rukungu G. In vitro activity of total aqueous ethanol leaf extracts of Ricinus communis on Leishmania major promastigotes. Kenya J Sci Technol. 2006;13(1):1–4.
Wabwoba BW, Anjili CO, Ngeiywa MM, Ngure PK, Kigondu EM, Ingonga J, Makwali J. Experimental chemotherapy with Allium sativum (Liliaceae) methanolic extract in rodents infected with Leishmania major and Leishmania donovani. J Vector Borne Dis. 2010;47(3):160.
Mohamed-Ahmed AH, Seifert K, Yardley V, Burrell-Saward H, Brocchini S, Croft SL. Antileishmanial activity, uptake, and biodistribution of an Amphotericin B and poly (α-Glutamic Acid) complex. Antimicrob Agents Chemother. 2013;57(10):4608–14.
La Flamme AC, Scott P, Pearce EJ. Schistosomiasis delays lesion resolution during Leishmania major infection by impairing parasite killing by macrophages. Parasite Immunol. 2002;24(7):339–45.
Nolan TJ, Farrell JP. Experimental infections of the multimammate rat (Mastomys natalensis) with Leishmania donovani and Leishmania major. Am J Trop Med Hyg. 1987;36(2):264–9.
Chen Y, Yan T, Gao C, Cao W, Huang R. Natural products from the genus Tephrosia. Molecules. 2014;19(2):1432–58.
Dharmananda S. Gallnuts and the uses of tannins in Chinese medicine. Chennai: ITM; 2003.
Ferguson LR. Role of plant polyphenols in genomic stability. Mutat Res. 2001;475(1):89–111.
Tasdemir D, Kaiser M, Brun R, Yardley V, Schmidt TJ, Tosun F, Rüedi P. Antitrypanosomal and antileishmanial activities of flavonoids and their analogues: in vitro, in vivo, structure–activity relationship, and quantitative structure–activity relationship studies. Antimicrob Agents Chemother. 2006;50(4):1352–64.
Murillo G, Salti G, Kosmeder Ii J, Pezzuto J, Mehta R. Deguelin inhibits the growth of colon cancer cells through the induction of apoptosis and cell cycle arrest. Eur J Cancer. 2002;38(18):2446–54.
Wenzel UA, Bank E, Florian C, Förster S, Zimara N, Steinacker J, Klinger M, Reiling N, Ritter U, van Zandbergen G. Leishmania major parasite stage-dependent host cell invasion and immune evasion. FASEB J. 2012;26(1):29–39.
Carneiro SM, Carvalho FA, Santana LC, Sousa AP, Neto JM, Chaves MH. The cytotoxic and antileishmanial activity of extracts and fractions of leaves and fruits of Azadirachta indica (A Juss.). Biol Res. 2012;45(2):111–6.
Shio MT, Hassani K, Isnard A, Ralph B, Contreras I, Gomez MA, Abu-Dayyeh I, Olivier M. Host cell signalling and Leishmania mechanisms of evasion. J Trop Med. 2011;2012:819512.
Olivier M, Gregory DJ, Forget G. Subversion mechanisms by which Leishmania parasites can escape the host immune response: a signaling point of view. Clin Microbiol Rev. 2005;18(2):293–305.
Authors’ contributions
This work was carried out in collaboration between all authors. SMN, CKW, COA and JAM conceived and designed the study. SMN, BNJ and CKW performed the experiments. SMN, CKW, JAM, BNJ, JKC, EOA and COA gave substantial inputs in the draft manuscript. SMN and CKW gave substantial inputs to the plan for analysis and analyzed the data. All the authors participated in drafting and revising the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The authors thank Mr. Lucas Ogutu of Kenya Medical Research Institute (KEMRI) for rearing the animals used in this study. We thank Dr. Oyoo-Okoth Elijah, for the technical assistance during data analysis support in the different facets of this study.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The data and study tools/materials that support the findings of this study are available from the Department of Biological Sciences, School of Science, University of Eldoret but restrictions apply to the availability of these data, which were used under license for the current study, and so are not publicly available. Data are however available from the authors upon reasonable request and with permission of Department of Biological Sciences repository.
Consent to publish
Not applicable.
Ethics approval and consent to participate
All procedures were per the Kenya Medical Research Institute (KEMRI) operational and approved protocols under the bioprospecting for plant based anti-leishmanial compounds programme/theme as regulated by the Scientific Steering Committee (SSC), Ethical Review Committee (ERC) and Animal Care and Use committee (ACUC). Approval to conduct the study was further obtained from the University of Eldoret, Faculty of Science board of postgraduate research clearance committee. The guidelines were strictly adhered to during the research.
Funding
Not applicable.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Marango, S.N., Khayeka-Wandabwa, C., Makwali, J.A. et al. Experimental therapeutic assays of Tephrosia vogelii against Leishmania major infection in murine model: in vitro and in vivo. BMC Res Notes 10, 698 (2017). https://doi.org/10.1186/s13104-017-3022-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13104-017-3022-x