
Abstract
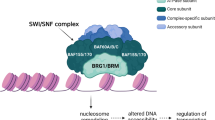
BRM (BRAHMA) is a core, SWI2/SNF2-type ATPase subunit of SWI/SNF chromatin-remodelling complex (CRC) involved in various important regulatory processes including development. Mutations in SMARCA2, a BRM-encoding gene as well as overexpression or epigenetic silencing were found in various human diseases including cancer. Missense mutations in SMARCA2 gene were recently connected with occurrence of Nicolaides–Baraitser genetics syndrome. By contrast, SMARCA2 duplication rather than mutations is characteristic for Coffin–Siris syndrome. It is believed that BRM usually acts as a tumour suppressor or a tumour susceptibility gene. However, other studies provided evidence that BRM function may differ depending on the cancer type and the disease stage, where BRM may play a role in the disease progression. The existence of alternative splicing forms of SMARCA2 gene, leading to appearance of truncated functional, loss of function or gain-of-function forms of BRM protein suggest a far more complicated mode of BRM-containing SWI/SNF CRCs actions. Therefore, the summary of recent knowledge regarding BRM alteration in various types of cancer and highlighting of differences and commonalities between BRM and BRG1, another SWI2/SNF2 type ATPase, will lead to better understanding of SWI/SNF CRCs function in cancer development/progression. BRM has been recently proposed as an attractive target for various anticancer therapies including the use of small molecule inhibitors, synthetic lethality induction or proteolysis-targeting chimera (PROTAC). However, such attempts have some limitations and may lead to severe side effects given the homology of BRM ATPase domain to other ATPases, as well as due to the tissue-specific appearance of BRM- and BRG1-containing SWI/SNF CRC classes. Thus, a better insight into BRM-containing SWI/SNF CRCs function in human tissues and cancers is clearly required to provide a solid basis for establishment of new safe anticancer therapies.
Similar content being viewed by others

Main text
Background
The genomic DNA in eukaryotic cells nuclei is packaged together with histone proteins into a complex called chromatin, enabling the storage of a relatively large amount of DNA in a very compacted form. However, the structure of chromatin restricts the contact between DNA and various protein (e.g. activators, repressors, modifying enzymes) or non-protein (e.g. enhancers, silencers) regulatory elements. Therefore, chromatin is a target for various modifications including chromatin remodelling, which controls the access to DNA sequences. The process is executed by multiprotein chromatin-remodelling complexes (CRCs), which utilise energy from ATP hydrolysis [1].
BRM ATPase, the SWI2/SNF2-type core subunit of SWI/SNF chromatin-remodelling complexes
One of the best-characterised chromatin-remodelling complexes (CRCs) are SWI/SNF CRCs. Originally, the SWI/SNF CRC was described in baker’s yeast, where it affected mating-type switch (SWI) [2] and sucrose fermentation (SNF—sucrose non-fermenting) [3, 4]. The SWI/SNF complexes were thus named for these phenotypic alterations [5]. Homologues of yeast SWI/SNF CRCs’ subunits were subsequently found in other organisms including humans [6], proving that they are highly evolutionary conserved among all Eukaryotes [1]. The SWI/SNF CRCs are involved in the regulation of various crucial cellular processes such as the cell cycle, cell morphology and adhesion, apoptosis, signal transduction, DNA repair and stress response, which are frequently and significantly altered in cancer [7,8,9,10].
SWI/SNF CRCs are multiprotein complexes, built from 10 to 15 subunits. Depending on the subunit composition, several classes of SWI/SNF CRCs may exist in the cell simultaneously [11]. The SWI/SNF subunit composition and activity is cell/tissue-specific [12].
It was believed that the core complexes of all types of SWI/SNF CRCs consist of four core subunits—one of two ATPase subunits: BRM (encoded by SMARCA2 gene) or BRG1 (encoded by SMARCA4 gene), BAF155 (encoded by SMARCC1 gene), BAF170 (encoded by SMARCC2 gene) and INI1 (SNF5 or BAF47, encoded by SMARCB1 gene) [7, 13]. Together with the core complex, non-core subunits are present in the SWI/SNF CRCs. The number of non-core subunits may differ and their composition influences activity of the whole complex.
Recent studies by several research groups indicated the existence of non-canonical SWI/SNF CRC classes lacking some core (i.e. INI1) and non-core subunits [14, 15]. Furthermore, the presence of non-canonical SWI/SNF complexes was also shown using mouse models [16].
The SWI/SNF CRCs utilise energy from ATP hydrolysis to disrupt contact between the DNA and histones, leading to nucleosome disassembly [17, 18]. They control gene expression by moving or removing nucleosomes covering binding sites for transcription factors [19] or stabilising nucleosome positions. The activity of SWI/SNF CRCs requires recruitment to the DNA by transcription regulators and other factors [20]. The action of SWI/SNF CRCs alters upon interactions with various proteins such as hormone receptors [21], acetylases/deacetylases, etc. and depends on the modification of its subunits by, e.g. acetylation, as has been observed for the BRM ATPase subunit. A comprehensive summary of SWI/SNF CRCs’ action was provided by Sarnowska et al. [19].
BRM and BRG1 ATPase subunits are critical for the SWI/SNF activity. Both of them belong to the SWI2/SNF2 family, share about 75% structural homology and share similar ATPase and helicase activities [6], although their function is not identical. In humans, BRG1 ATPase may be present in both SWI/SNF CRC classes—BAF (BRM or BRG1-associated factors) and PBAF (polybromo BRG1-associated factors), while BRM has been found in BAF class of SWI/SNF complexes only and is the so-called signature subunit of this complex class. BRM has lower ATPase activity than BRG1 [22, 23], therefore, its less important role was postulated.
This hypothesis has been supported by mouse models where Brm-knockout (Brm−/−) mice lived until adulthood and developed tumours while Brg1−/− null mutants caused embryonic lethality [24]. It has also been shown that Brm controls cellular proliferation by regulation of the cell cycle [25]. The mouse model study revealed that both homozygous and heterozygous loss of Brm resulted in an increased risk of tumour development, when exposed to carcinogens [22]. Therefore, it is proposed that Brm rather acts as a cancer susceptibility than a tumour suppressor gene [26]. The importance of Brm in mice has been shown by several additional studies, i.e. using conditional knockout of both genes encoding BRM and BRG1 ATPases in heart. In this case, the concomitant depletion of Brm and Brg1 resulted in severe cardiac dysfunction associated with glycogen accumulation and mitochondrial defects, eventually leading to death [27]. Moreover, functional Brm protein is crucial for the initiation of regeneration phase after liver injury and dominates during the late injury phase on Brg1 function [28].
The double-knockout mice Brm−/−/Brg1−/− exhibited an unexpected ability to overcome loss of both ATPases. In fact, Brm−/−/Brg1−/− mice restored Brm expression via an alternative splicing strategy which resulted in production of truncated but functional Brm protein [29]. This study, together with the existence of alternative splicing variants of SMARCA2 gene [30], indicate that the role of BRM may be more complicated than so far reported. It may be due to the fact that several different forms of BRM protein may exist in the cell.
The loss of human BRM or BRG1 consequently leads to the modified expression of genes that are significant for tumour development, e.g. genes encoding tumour suppressors. Many of them control cellular processes such as metabolism (including drug metabolism), DNA repair, differentiation, adhesion and apoptosis, and are involved in angiogenesis, progression or metastasis of cancer [31]. Recent studies on ovarian cancer revealed that BRG1 and BRM ATPases are mutually exclusive as their parallel inactivation leads to synthetic lethality [32], although other reports indicated the survival of cells with depletion of both ATPases [31]. One possible explanation of this apparent discrepancy may be the existence of shorter, truncated but functional versions of BRM protein which are simply not recognised by the anti-BRM antibody.
In human heterozygous missense, mutations in BRM-encoding SMARCA2 gene were identified in patients with Coffin–Siris (CSS) and Nicolaides–Baraitser (NCBRS) syndromes [33]. Although the last study performed on a large cohort of CSS patients proved that they carry the missense mutation in SMARCA4 (BRG1 encoding gene) gene but not in BRM-encoding SMARCA2 gene. In some CSS patients, duplication of the SMARCA2 gene was detected [34]. On the other hand, missense mutations in SMARCA2 were detected in NCBRS patients [35]. Moreover, one individual with SMARCA2 mutation was previously diagnosed with CSS and after was reclassified for NCBRS [36, 37]. This collectively suggests that missense mutations in SMARCA4 and SMARCA2 may cause different developmental disabilities, although both CSS and NCBRS syndromes share some similar developmental dysfunctions and their distinction is based mostly on foot and hand features [35] (Fig. 1).

The phenotypic differences and commonalities between NCBRS and CSS. The red lines correspond to mutation sites in SMARCA2 and SMARCA4 genes according to [136]
The occurrence of developmental alterations in CSS and NCBRS could be in contradiction with mouse models, where heterozygous loss in the Brm-encoding gene has no developmental effects. Although, in CSS the duplication of SMARCA2 gene may lead to overexpression of BRM protein and as a consequence altered SWI/SNF stoichiometry caused by the pathological competition of BRM with BRG1 ATPase. In NCBRS, the missense mutations in SMARCA2 may result in BRM gain-of-function or loss of function, i.e. by fast protein degradation. The exact role of SMARCA2 missense mutations or duplication during development is still unclear; however, the SMARCA2 polymorphisms may lead to higher cancer risk, suggesting the role of human BRM as a cancer susceptibility gene, similarly to mice [38].
It is also important to note that the classes of human SWI/SNF CRCs containing BRG1 or BRM subunits may regulate different promoters and sometimes they even differentially regulate transcription of the same genes [19]. This could be based on the differences in transcription factor recruitment, subunit composition and the occurrence of differential modifications of SWI/SNF subunits [23]. The picture of BRM/BRG1 interdependence is broadened by the in-depth, high throughput study based on ChIPseq and RNAseq analysis of BRM or BRG1-depleted HepG2 cell line. This study revealed that depletion of one ATPase subunit frequently leads to decreased abundance of the remaining subunit. Additionally, on numerous genes, the remaining subunit is either retained or gained [39]. All the above data suggest that in cancer, both ATPases have similar functions and could be partially redundant.
Mechanism of action
BRM-containing SWI/SNF CRCs regulate expression of a large number of genes involved in carcinogenesis including (i) epithelial–mesenchymal transition genes, e.g. CDH2 (N-cadherin) and SNAI1; (ii) cell cycle genes, e.g. CCND1 (cyclin D1), CCNE2 (cyclin E2), CDK4 and CDK6 (cyclin kinases), (iii) metabolic genes, e.g. GAPDH, ALDOA and LDHA; (iv) cancer suppressor genes and oncogenes, e.g. BRCA1, PTEN, AKT1, HRAS and KRAS. Importantly, BRM also regulates expression of self-encoding SMARCA2 gene and other SWI/SNF subunits [40].
BRM directly interacts with the retinoblastoma protein (Rb) and its family members [7, 41, 42]. Through this interaction, BRM influences cell cycle, causing repression of E2 promoter binding factor (E2F) family transcription factors [43]. Cells lacking BRM cannot enter the G1/S phase resulting in growth arrest [44] (Fig. 2). BRM function in the cell cycle is probably dependent on the phosphorylation of BRM by cyclin E/CDK2 complex causing dissociation of Rb from ATPase [45], and leading to cell cycle progression [46]. However, some data suggest that BRG1 has a more significant role in cell cycle control than BRM and, therefore, the specific role of BRM in Rb-mediated cell cycle inhibition remains elusive [47].

BRM-involving cellular processes. BRM protein is involved in a variety of cellular processes both in healthy/normal and cancerous cells; for example, gene expression control, alternative splicing, cell cycle control, participating in hormonal response and miRNA transcription and signalling. In pathological situations, in cancer cells, BRM can leave the cell nuclei and migrate to cytoplasm or cell membrane, although specific effects of BRM in these locations are unknown
The link between SWI/SNF CRCs and TP53, a commonly mutated oncogene, is also known [48]; however, the particular role of BRM ATPase in this dependency is still unclear. Xu and colleagues demonstrated that BRM and BRG1 affect TP53-dependent p21 transcription differently. BRG1 knock-down handicaps TP53 binding to p21 promoter although BRM has ability to replace BRG1 in TP53 regulation [49].
Interestingly, mice lacking Brm did not present pathological Tp53 mutations in tumours, although such mutations were accumulated in Brm-positive tumours, suggesting that loss of Brm would restrain selection of Tp53-mutated variant in tumour evolution. Collectively, this observation strongly suggests the existence of various ways of tumour evolution and development [47].
Some data indicate SWI/SNF participation in DNA damage response. BRM is involved in non-homologous end-joining (NHEJ) DNA repair, although its activity in this process depends on SWI/SNF complex composition [50]. BRM recruitment to double-strand breaks depends on, i.e. histone 2B phosphorylation on Ser36 which promotes BRM involvement in this process. Ribeiro-Silva and colleagues [51] observed that BRM is required for correct recruitment of the transcription factor II H (TFIIH) to the DNA damage site and facilitates DNA nucleotide excision repair pathway. Moreover, the SWI/SNF CRC also participates in DNA damage repair by interactions with BRCA1, indicating its important role in homologous recombination [52] (Fig. 2).
BRM also plays an important role in regulation of alternative splicing via interaction with spliceosome components. BRM overexpression favours inclusion of alternative exons [53], which is consistent with the observation that BRM is present not only on promoter regions, but also in the gene body [39] (Fig. 2).
Sakurai et al. [54] and Kobayashi et al. [55] demonstrated that BRM participates in the miRNA containing axis, particularly miR-199 and transcriptionally regulates the miR-302a-3p expression [56]. BRM is also involved in JAK2/STAT3 pathway, causing its activation [57]. The SWI/SNF BRM-containing CRC acts in the control of hormonal signalling pathways and participates in hormonal crosstalk [19]. In particular, BRM is required for proliferation of androgen-dependent prostate cancer [58] and regulates androgen receptor (AR) target genes expression [59]. Additionally, in cooperation with prohibitin, BRG1 or BRM ATPases are crucial for oestrogen antagonist-mediated breast cancer growth suppression [60]. The known BRM interactors and processes involving BRM are summarised in Table 1. The large interaction network of BRM suggests a strong potential impact of any BRM impairment on numerous regulatory processes.
BRM aberrations in human cancer
The importance of an altered level and/or aberrant function of BRM in various cancers is not fully understood, although there is abundant evidence indicating the crucial BRM role in carcinogenesis. About 15% of all cancers display numerous aberrations in BRM abundance or impairment, that may lead to cancer development or progression (Fig. 3).

Schematic summary of BRM role in cancer development. ↑—upregulation; ↓—downregulation; TNBC triple-negative breast cancer, HCC hepatocellular carcinoma, UATC upper aerodigestive tract, SCCOHT small cell carcinoma of the ovary, hypercalcaemic type, OCCC ovarian clear cell carcinoma; OCC ovarian cell carcinoma, HNSCC head and neck squamous cell carcinoma, ACC adenoid cystic carcinoma; NSCLC non-small cell lung cancer; AD adenocarcinoma od the lung, LC large cell carcinoma of the lung; PL pleomorphic carcinoma of the lung; ccRCC clear cell renal cell carcinoma, NMSC non-melanoma skin cancer
Rhabdoid tumours
Malignant rhabdoid tumour (MRT) is an extremely aggressive type of cancer that affects mostly children. The mutations in SMARCB1 an INI1/SNF5/BAF47-encoding gene were found in this type of cancer, suggesting the mutation in a gene encoding core SNF5-type subunit of SWI/SNF CRCs as a driving mutation for this cancer type [61, 62]. This INI1 alteration was accompanied by BRM epigenetic silencing in about 70% of MRT cases. BRM was silenced by the HDAC-driven mechanism or by SMARCA2 promoter polymorphisms [53]. Interestingly, BRM expression was induced by INI1 re-expression and synthetic flavonoid treatment. BRM re-expression was necessary for flavonoid or INI1 re-expression induced growth inhibition of rhabdoid cell line [63].
Lung cancer
The downregulation of BRM is most frequently reported in lung cancer (LC) especially in non-small cell lung carcinoma (NSCLC). In case of the adenocarcinoma (AD), the amount of cases with BRM depletion is 6–17% [64, 65], although in the group of poorly differentiated ADs the fraction of cases with BRM depletion reaches 92% [64,65,66]. The highest downregulation of BRM protein was observed in the pleomorphic carcinoma of the lung (PL)—over 40% cases [66], and in the large cell carcinoma of the lung (LCCL)—33–50% cases [64, 66]. BRM staining is positive in lepidic growth components in LC and becomes significantly lower or negative in invasive parts [65, 66].
The BRM level in the primary NSCLC is associated with the survival rate. The overall survival for the group of patients with loss of nuclear BRM was significantly lower comparing to patients with high BRM level. Additionally, a membrane form of BRM was observed in immunohistochemistry in some samples. In the case of AD, 5-year survival of patients with positive staining for the membrane BRM was significantly lower than in case of patients without the membrane BRM, suggesting that the presence of the membrane BRM form may be a good prognostic marker [67]. However, the function of the membrane form of BRM remains unknown and there are no data regarding such localisation of other subunits of SWI/SNF CRCs. Intriguingly, loss of BRM and BRG1 in this type of cancer is correlated with loss of tumour cells’ ability to differentiate [31, 65]. A correlation between loss of BRG1 and BRM and epithelial–mesenchymal transition (EMT) in lung cancer was reported, especially in poorly differentiated ADs. Loss of BRM protein was more frequent in heavy smokers, supporting the hypothesis that BRM depletion enhances susceptibility to cancer induced by the carcinogen exposure [65, 68]. On the other hand, targeting BRM in the BRG1-deficient lung cancer (NSCLC) sensitised cancer cells (cell lines) to radiotherapy [69].
Renal cell carcinoma
SWI/SNF CRCs are aberrant in clear cell renal cell carcinoma (ccRCC), the most common type of renal cancer [70, 71]. The 3% of all ccRCC cases were BRM-negative. All BRM-negative cases were classified as poorly differentiated grade 4 tumours [72]. It is also significant that loss of BRM was observed only in ccRCC, and not in other types of renal cancers like fumarate hydratase-deficient RCC [73].
Interestingly, in the case of RCC, the same pattern as in lung cancer was observed—poorly differentiated tumours or poorly differentiated parts of tumours were lacking a BRM subunit, while more differentiated areas showed BRM expression [72, 74] suggesting that alterations of SMARCA2 gene occur during de-differentiation of ccRCC. Thus, BRM loss may be specifically linked to the tumour aggressiveness. In BRM-deficient ccRCC tumours almost 90% cases displayed genetic alterations in the SMARCA2 gene, such as mutations, promoter methylation or chromosomal aberrations. Interestingly, the alterations were very rare in low-grade part of analysed tumours and in non-neoplastic tissue [74]. These observations strongly suggest that loss of BRM occurred during cancer progression.
Cancers of gastrointestinal tract
Loss of SWI/SNF subunits correlates with undifferentiated tumour phenotypes in gastrointestinal tract (GI) cancers. Interestingly, in rare GI cancers, with rhabdoid features no concurrent loss of BRM and BRG1 ATPases was observed. On the other hand, concomitant inactivation of BRM and INI1/SNF5/BAF47 frequently lost in rhabdoid carcinomas was reported [75].
Inactivation of the BRM-encoding SMARCA2 gene by the presence of promoter indel polymorphisms correlates with higher risk of colorectal cancer [76]. These promoter indels increase risk of upper aerodigestive tract cancers (UATC) more than twofold suggesting that BRM downregulation may be significant for development and progression of GI cancers [77].
The reduction of BRM expression was observed in gastric cancers, while in such cases, BRG1 level was unaffected [78]. Importantly, this effect was characteristic only for cancer cells, but was not seen in premalignant lesions suggesting the importance of BRM loss at later stages of the stomach cancer development [79]. In about 10% of gastric cancers, methylation of SMARCA2 promoter region was identified [79].
Altered levels of BRM expression are also observed during pancreatic cancer development, although the mechanism is not yet fully understood. High levels of BRM are associated with patients’ poor survival, linked to larger tumour size, metastasis to other organs, lymphatic invasion and stage IV disease [80]. On the other hand, the downregulation of BRM may be a significant marker in the pancreatic cancer [57], indicating that BRM levels undergo dynamic changes in different stages of the disease. BRM silencing in pancreatic cancer cell line correlates with lower cell viability, proliferation rate and growth both in vitro and in vivo. This fact may be contrasted with the observation that downregulation of BRM is vital for pancreatic cancer progression [57] indicating that BRM is actually essential for this process. Intriguingly, in data presented by Shain et al., no pancreatic cancer cell lines with decreased BRM expression were observed [81]. A recent study indicated that BRM promotes pancreatic cancer growth and chemoresistance via activation JAK2/STAT3 pathway [57]. It also transcriptionally regulates the miR-302a-3p and promotes pancreatic cancer metastasis by epigenetic modulation of SOCS5/STAT3 signalling axis [56]. However, to the large extent, the role of BRM in pancreatic cancer remains elusive.
Hepatocellular cancer
According to current knowledge, normal human hepatocytes display negative staining for BRG1 and positive for BRM protein [82]. The lack of BRG1 protein in normal human hepatocytes is, however, counterintuitive to expectations, especially given that in mouse models Brg1 is more important than Brm for liver regeneration after injury [28]. By contrast, a majority of hepatocellular carcinoma (HCC) is characterised by the positive BRG1 staining. In 22.5% of HCC cases, the loss of BRM protein was found while in 15% of analysed cases both BRM and BRG1 were lost. The depletion of BRM in HCC significantly corresponded to poor overall survival [82]. Pasic et al. correlated this phenomenon with the occurrence of BRM promoter polymorphisms that were also found in other cancers, leading to poorer patient survival [83].
Interestingly, in HCC tumour cells, additional cytoplasmic localisation of BRM was found, while no such cases were observed in healthy hepatocytes. This indicates that changes in BRM localisation may also contribute to carcinogenesis, although the mechanism of this phenomenon remains unknown [82].
Head and neck cancers
The head and neck squamous cell carcinoma (HNSCC) is one of the most common head and neck cancers, accounting for up to 90% of cases. In 16% of patient samples, the total loss of BRM was demonstrated, 11% featured weak staining and 16% were mosaic. Occurrence of SMARCA2 promoter region polymorphisms correlates with HNSCC risk, especially in HPV-positive oropharyngeal cancer [84].
In 5% of cases of salivary gland adenoid cystic carcinoma (ACC), the SMARCA2 mutation was found [85]. Additionally, both mRNA and protein level of BRM were significantly elevated in ACC cells, comparing to the healthy tissue. Interestingly, the BRM overexpression was observed in every sample in all tumour areas, although ACC is the most heterogeneous cancer type. This strongly suggests that salivary gland ACC is characterised by BRM overexpression [86] although the specific mechanism of BRM action in the ACC cells needs further elucidation.
Breast cancer
Cohet et al. described that the presence of both SWI/SNF ATPases is crucial for optimal cell cycle progression in non-malignant mammary epithelial cells and knock-down of either BRM or BRG1 affects cell cycle, while the double knock-down of BRM and BRG1 results in cell death [87].
The analyses of a set of different breast cancers with various subtypes and stages revealed the increase of BRM levels in a significant majority of analysed cases [88]. However, the authors did not mention whether nuclear, cytoplasmic or other BRM staining was taken into consideration. Additionally, it was observed that both BRG1 and BRM are required for the triple-negative breast cancer (TNBC) proliferation and that double SMARCA2 and SMARCA4 knock-down results in slowed tumour growth in xenografts [88]. By contrast, a statistically significant downregulation of SMARCA2 transcript was observed in all breast cancer types comparing to healthy tissue. Based on this observation, it was postulated that expression of SMARCA2 and SMARCA4 has prognostic value [89], although another study shows that the BRM protein level varies among various breast cancer types. Namely, decreased level of BRM was observed in MDA-MB-231 (TNBC cancer cell line) comparing to the less malignant MCF-7 (ER positive) cells. Furthermore, BRM regulates tight junction protein expression via targeting their promoters, thus takes part in the breast cancer metastasis [90].
Gynaecological cancers
Although in gynaecological cancers many alterations of SWI/SNF subunits were observed [91,92,93,94], differences in BRM expression were found only in small cell carcinoma of the ovary, hypercalcaemic type (SCCOHT), ovarian cell carcinoma (OCC) and in the endometrial cancer.
SCCOHT is a rare subtype of the ovarian cancer that affects mainly young women. At first, loss of BRG1 protein caused by somatic and germline mutations in SMARCA4 gene (coding for BRG1 protein) in SCCOHT was identified by a few groups [95,96,97,98,99]. Recently, loss of BRM protein was found in this type of cancer [100, 101]. Importantly, no mutations in SMARCA2 gene were found, suggesting epigenetic control of SMARCA2 gene expression.
Noteworthy, for SCCOHT analysis, the ovarian clear cell carcinoma (OCCC), a different type of ovarian cancer, was used as a basis for comparison. Interestingly, in OCCC tumours, the lack of BRM or BRG1 was found but never loss of both ATPases. The mutations in the SMARCA2 gene, in about 2% of OCCC samples, were described [102]. Although, in epithelial ovarian cancer (the most common type of ovarian cancer) BRM overexpression strongly correlates with resistance to cisplatin, probably due to the reduction of apoptosis and influence of metabolism and cancer-associated signalling pathways [103]. These findings confirmed earlier discoveries that the downregulation of BRM increases cisplatin sensitivity [104].
The lack of both ATPases also was found in endometrial cancer [105], although no information about clinical significance of such loss has been so far described.
Prostate cancer
Analysis of prostate cancer samples revealed that level of BRM was significantly downregulated in primary prostate tumours and metastases, although BRG1 level increased with the disease progression [106, 107]. The slightly lower signal for BRM was found in malignant sites comparing to the non-invasive parts of cancer [107]. BRM has been found to be required for the proliferation of AR-dependent prostatic adenocarcinoma cells [58].
Skin cancers
Alterations in BRM expression seem to be important for the non-melanoma skin cancer (NMSC) development, and in this case, the role of SMARCA2 as a susceptibility gene is strongly pronouncing. Analyses of NMSC patient samples, specifically squamous cell carcinoma (SCC) and basal cell carcinoma (BCC) with comparison to benign precancerous lesions—actinic keratosis (AK), which is claimed to be a progenitor for SCC and BCC development, and normal skin showed that the transcript level of SMARCA2 gene decreased in SCC cells. By contrast, no differences were observed for SMARCA4 gene [108]. Surprisingly, at the protein level, both BRM and BRG1 were downregulated about tenfold in both SCC and BCC comparing to AK and normal skin, indicating altered regulation of their expression through some other mechanism. The authors suggested that loss of ATPases happens after the development of benign skin lesions, where no downregulation of BRM was observed [108]. Additionally, a mutation in SMARCA2 gene was identified in 17% of NMSC. Since this mutation was observed only in SCCs and BCCs and not in precancerous lesions or normal skin, this observation suggested that the mutation was preferentially selected in the process of cancer development [109]. Subsequently, a possible mechanism underlying BRM loss in NMSC was discovered based on a mouse model. It was found that mouse keratinocytes with deleted Brm (Brm−/−) grew faster than normal (Brm+/+) after UV-irradiation. This suggests the ability of Brm null mutant keratinocytes to escape UV-induced cell cycle arrest faster than in normal cells. Moreover, although Brm−/− keratinocytes accumulate more DNA damage, they do not compensate for this with more intense DNA repair [110]. All these effects result in the ability of Brm null mutant keratinocytes to undergo selective pressure that can cause overgrowth of cells with accumulated mutations over normal cells and hence lead to cancer development [110].
SWI/SNF ATPases are also claimed to be important components in regulation of microphthalmia-associated transcription factor (MITF) expression. MITF is a key factor linked to development of melanoma. BRG1 was described as a main epigenetic regulator of MITF expression. However, in case of cancers with inactivated BRG1, BRM replaces siblings ATPase and performs their function. In such cases, pharmacological exclusion of BRM could lead to the reduction of melanoma growth or even cancer cell death [111].
Other neoplasms
In case of leukaemia, evidence of the importance of BRM protein is rather weak. In this cancer type, SWI/SNF complexes are mainly built around the BRG1 ATPase that is essential for survival and growth of this neoplasm [112]. BRM is the main ATPase expressed in quiescent hematopoietic stem cells suggesting that loss of BRM significance takes place at the time of leukaemia development. This hypothesis is supported by discovery of Doménech et al. [113] who identified mutations in SMARCA2 gene in leukemic cells. In acute myeloid leukaemia with monosomy, 7 (about 13% of cases) novel mutations of SMARCA2 gene were identified, although these are not thought to be driving mutations because the samples were collected at more advanced stages of carcinogenesis [114]. In proximal-type epithelioid sarcoma with pure rhabdoid tumour features, additional loss of BRM was found [115]. Additionally, the occurrence of SNPs in the SMARCA2 gene seems to correlate with risk for oligodendroglioma development [116].
Collectively, in various types of cancer, loss or strong decrease of BRG1 or BRM and sometimes both ATPases, was observed (Table 2). BRG1 and BRM seem to have similar/redundant function in cancer cells, although, they play different roles during human development. This hypothesis supports the observation that mutation in BRG1 encoding gene is frequently associated with CSS while the mutation in BRM-encoding gene is more typical for NCBRS. These two different genetic disorders carry some similar phenotypic aberration but also differ from each other (see Fig. 1). The exact role of BRM and BRG1 in somatic cells and how the imbalance in their abundance trigger to disorders or cancer development still remains unknown. Therefore, investigation of BRM and BRG1 differential function in various cancer types and in somatic cells seems to be one of the most exciting and important directions for further research.
Mechanism of BRM alteration in cancer
Alterations of BRM were reported in various cancer types, but only in some of them, SMARCA2 mutations were found. Therefore, it is highly probable that various mechanisms of BRM control exist including mutation-independent regulation of SMARCA2 gene expression.
Mutations
In the majority of cancers lacking BRM, no mutations of the SMARCA2 gene were found suggesting that epigenetic regulation plays more crucial role in the BRM inactivation [82, 117, 118]. However, SMARCA2 mutations were found in 78.2% of BRM-deficient ccRCC cases, although about half of them were silent [74]. All detected mutations were specific for cancerous tissue, especially low differentiated, and none or very low-level mutations were found in BRM-positive tissues as well as adjacent non-malignant tissues [74]. SMARCA2 gene mutations were also identified in about 10% of gastric cancers [79]. In NMSC, a mutation resulting in substitution of glutamine by lysine was discovered. Interestingly, this mutation type—G:C to T:A substitution is observed after UV-irradiation, what is a usual cause of skin cancer [109]. SMARCA2 mutations of unknown effect were also found in leukaemia [113].
Epigenetic modifications
Methylation of CpG sites on SMARCA2 promoter region is the key mechanism of BRM alterations [74]. It was recently found that Polycomb repressive complex 2 (PRC2) causes epigenetic suppression of SMARCA2, by inducing H3K27me3 silencing pattern on its promoter region [119].
In BRM, deficient ccRCC methylation refers to over 40% of cases and was found only in low-differentiated tumour areas [74]. A similarly observation was made in the AD of the lung, where SMARCA2 promoter methylation correlated with poor prognosis [40]. Nonetheless, in some cancers with lack/low BRM level, no mutations and no hypermethylation were found in the SMARCA2 locus [108] suggesting the existence of other mechanisms involved in the control of BRM expression or protein stability.
Additionally, the activity of BRM is modulated by posttranslational modifications, for example acetylation can lead to BRM inactivation [120]. Three acetylation sites were identified in the BRM protein, although their specific functions remain elusive, and whether acetylation of only one or all sites is required for BRM inactivation is unknown [121].
Promoter insertion polymorphisms
Insertions in the promoter sequence of SMARCA2 gene (at positions −741 and −1321) was specified as silencing-type polymorphism, leading to development of many types of cancer [122]. Interestingly, the promoter insertions cause HDACs recruitment and result in SMARCA2 gene silencing [68]. Such insertions were associated with higher risk of lung cancer [68], colorectal cancer [76], and head and neck squamous cell carcinoma [84]. Increased risk of developing upper aerodigestive tract cancers appears only in case of double homozygous variants of such polymorphisms [77]. In the case of pancreatic cancer, the presence of SMARCA2 promoter polymorphisms is associated with poor prognosis for patients with diagnosed cancer rather than specific cancer risk [123].
Chromosomal aberrations
The loss of chromosome 9p, the SMARCA2 gene location, results in BRM loss and enhanced cancer aggressiveness. In ccRCC aberrations of chromosome, 9p (monosomy or deletion) was found in over 40% of analysed BRM-deficient tumours [74]. Currently, the loss of 9p chromosome is used as a prognostic marker for ccRCC [124].
Moreover, loss of the BRM subunit may occur in cells with multiplication of chromosome 9. Multiple abnormalities also may lead to simultaneous silencing of all copies of the gene, e.g. by mutation and CpG methylation that occur in the same cancer cell [74].
BRM overexpression
Although a few malignancies with the upregulation of BRM were identified, a specific mechanism leading to BRM overexpression in neoplasms was not identified. In ACC samples, BRM was elevated on both protein and transcript level [86]. Overexpression of BRM correlated with poor survival and chemoresistance in pancreatic cancer. Similarly, in ovarian cancer, high level of BRM promoted resistance to cisplatin [80, 103].
New treatment strategies in BRM-altered cancers
Only some malignancies with overexpression of BRM protein were identified, whilst in most of the neoplasms, the BRM is strongly downregulated or lost. The restoration of BRM protein in BRM-deficient cancers leads to impeded cancer cell growth [63]. This effect may be obtained with chemical compounds. Thus, the search of such compounds is encouraged [118, 125].
The first study considering direct BRM targeting was development of selective SMARCA2/4 bromodomain inhibitor (PFI-3), although PFI-3 did not reveal antiproliferative effect in cancer cells. This observation indicated that bromodomain is not a proper therapeutic target, and research focused on targeting helicase/ATPase domain in BRM for synthetic-lethality therapy [126]. Recently, small molecules for inhibition of BRM and BRG1 ATPase activity were discovered. In a BRG1-deficient lung cancer xenograft model, these inhibitors downregulated BRM-dependent gene expression and exhibited antiproliferative effect upon oral administration [127]. Another approach based on proteolysis-targeting chimera (PROTAC) has been designed to target BRM/BRG1 subunits of SWI/SNF CRCs employing a bromodomain ligand to recruit the VHL, the E3 ubiquitin ligase facilitating protein degradation [128].
Polymorphisms in SMARCA2 gene in position −747 and −1321 are suggested to be responsible for HDAC recruitment, and HDAC inhibitors (HDACi) cause upregulation of both BRM transcript and protein levels in cell lines [26, 100, 120, 129, 130]. Application of HDACi in patient groups seems to be a promising therapy, especially now as HDACi are approved by Food and Drug Administration and European Medicines Agency, and are becoming a more popular choice of treatment in various types of human cancer [131]. A similar effect on BRM expression was obtained during flavopiridol (synthetic flavonoid) treatment of rhabdoid cell lines [63]. Importantly, some studies demonstrated that whilst utilisation of HDACi effectively induces BRM, it also leads to an increase of BRM acetylation which impairs its function, giving no therapeutic effect overall [120].
In tumours with SMARCA2 depletion caused by PRC2-driven methylation, usage of EZH2 inhibitors seems to be a promising therapy. Effectiveness of this kind of drug was demonstrated for ovarian cancer (SCCOHT) in in vitro models [132]. Interestingly, in SCCOHT, a synergistic effect was observed, when EZH2 inhibitors were used together with HDACi [133]. Unfortunately, such an approach appeared to be not suitable for every type of cancer with BRM depletion: for instance, in lung cancer cell lines and pancreatic cancer cell lines, no therapeutic effect was observed.
BRM appeared as an attractive therapeutic target and induction of its activity may be helpful in cancer treatment. Therefore, a special reporter system was designed to identify novel compounds that restore not only BRM protein level, but also its function, giving hope for effective epigenetically focused treatment [125].
BRM and BRG1 are mutually exclusive ATPase subunits of SWI/SNF CRCs. Therefore, BRM targeting in BRG1-deficient cancer is expected to cause synthetic lethality [134]. So far, this phenomenon has been described for lung and ovarian cancers [126, 127, 135].
Possibilities for incorporation of BRM/SMARCA2-related targeted therapy into the clinic are still developing and new strategies seems to be very promising and effective (Fig. 4).

Cancer treatment related to BRM protein. Ideas for utilising BRM in anticancer therapy are emerging. Such therapies, taken currently into consideration, are based on BRM level restoration by, e.g. HDAC inhibitors and E2F inhibitors. A very promising but demanding idea is based on a synthetic lethality approach, targeted against BRM ATPase domain or bromodomain
Conclusions
BRM deficiency or downregulation was found in various types of cancer, although its function in cancer development and progression remains elusive. Interestingly, the mutation rate in SMARCA2 gene is quite rare compared to mutations in SMARCA4, suggesting a less important role of BRM ATPase containing SWI/SNF CRCs in carcinogenesis. Some data indicated SMARCA2/BRM as a tumour suppressor or tumour susceptibility gene, whereas overexpression of BRM caused cancer resistance for chemotherapy leading to cancer progression indicating its promoting role. All available data concerning BRM in cancer suggest that BRM function differs depending on cancer type. Thus, BRM acts in some cases as a tumour suppressor and in other cancer types or stages as a tumour or disease-promoting factor. Moreover, in some tumour types, loss or downregulation of BRM occurs during cancer development in late stage or in poorly differentiated/undifferentiated cancer cells suggesting clonal selection of BRM-deficient cancer cells. Additionally, some SMARCA2 missense mutations result not only in BRM loss of function but also may lead to production of gain-of-function BRM protein, likely influencing the whole BRM containing SWI/SNF CRC’s activity.
Based on ample recently published data, BRM seems to be a perfect target for various anticancer therapies including ATPase activity inhibition, synthetic lethality induction, etc. However, in view of current knowledge, i.e. given the high similarity of the ATPase domain among various proteins involved in the control of numerous regulatory processes, such optimism should be moderated by the risk of severe side effects of such therapies through off-target effects. During designing of the BRM-based anticancer therapy, another important fact needs to be urgently taken into consideration, namely, the observation that normal hepatocytes are negative for BRG1 ATPase. In such case, any BRM-targeting small molecule drugs can cause severe and unexpected negative effects on liver activity which may be impossible to detect during initial tests on cancer lines or in mice, as in this model, Brm seems to have a diverse function than in human liver. The damaging effect of BRM-targeting compounds likely may be observed in any tissue characterised by the lack of BRG1 protein. Therefore, additional study on BRM function in normal tissues and cancers is clearly required for better understanding of the interdependence between both ATPases, to precisely and safely treat cancer patients with new anti-BRM compounds or compound-based therapies.
Another important issue in the study of BRM function in development and carcinogenesis is the alternative splicing of SMARCA2 gene, which is relatively unexplored. According to the NCBI database, there are seven alternative transcripts of SMARCA2 gene. A similar situation is observed in mice, where six alternative transcripts exist. There are no data about the tissue or developmental stage-specific expression of these splicing variants either in mice or in humans. The relevance of BRM alternative splice variants in cancer is overlooked, although such a multiplicity of alternative BRM splicing variants suggests far more potential regulatory or pathological functions of the BRM protein which may be specific for certain cancer types or developmental stages. Knowledge about the existence of truncated BRM forms in cancer is very limited, although it is very likely that such incomplete but still partially functional proteins are produced due to unusual splicing events frequently occurring in various cancers. Such truncated BRM forms may have a very strong negative or gain-of-function effect on the functionality of the whole SWI/SNF complex, and thus may lead to the de-regulation of numerous important regulatory cellular processes fine-tuned by SWI/SNF complexes.
Availability of data and materials
Not applicable.
Abbreviations
- ACC:
-
adenoid cystic carcinoma
- AD:
-
adenocarcinoma of the lung
- ATP:
-
adenosine tri-phosphate
- CRC:
-
chromatin-remodelling complex
- ccRCC:
-
clear cell renal cell carcinoma
- CSS:
-
Coffin–Siris syndrome
- E2F:
-
E2 promoter binding factor
- EMT:
-
epithelial–mesenchymal transition
- GI:
-
gastrointestinal tract
- HCC:
-
hepatocellular carcinoma
- HDAC:
-
histone deacetylase
- HDACi:
-
histone deacetylase inhibitors
- HNSCC:
-
head and neck squamous cell carcinoma
- IHC:
-
immunohistochemistry
- LC:
-
lung cancer
- LCCL:
-
large cell carcinoma of the lung
- MITF:
-
microphthalmia-associated transcription factor
- NCBRS:
-
Nicolaides–Baraitser syndrome
- NMSC:
-
non-melanoma skin cancer
- NSCLC:
-
non-small cell lung cancer
- OCC:
-
ovarian cell carcinoma
- OCCC:
-
ovarian clear cell carcinoma
- PL:
-
pleomorphic carcinoma of the lung
- PRC2:
-
polycomb repressive complex 2
- Rb:
-
retinoblastoma protein
- RCC:
-
renal cell carcinoma
- SCC:
-
squamous cell carcinoma
- SCCOHT:
-
small cell carcinoma of the ovary, hypercalcaemic type
- SNP:
-
single-nucleotide polymorphism
- SWI/SNF:
-
switch/sucrose non-fermenting
- UATC:
-
upper aerodigestive tract cancers
References
Muchardt C, Yaniv M. ATP-dependent chromatin remodelling: SWI/SNF and Co. are on the job. J Mol Biol. 1999;293:187–98.
Haber JE, Garvik B. A new gene affecting the efficiency of mating-type interconversions in homothallic strains of Saccharomyces cerevisiae. Genetics. 1977;87:33–50.
Carlson M, Osmond BC, Botstein D. Mutants of yeast defective in sucrose utilization. Genetics. 1981;98:25–40.
Neigeborn L, Carlson M. Genes affecting the regulation of SUC2 gene expression by glucose repression in Saccharomyces cerevisiae. Genetics. 1984;108:845–58.
Peterson CL, Herskowitz I. Characterization of the yeast SWI1, SWI2, and SWI3 genes, which encode a global activator of transcription. Cell. 1992;68:573–83.
Chiba H, Muramatsu M, Nomoto A, Kato H. Two human homologues of saccharomyces cerevisiae SWI2/SNF2 and Drosophila brahma are transcriptional coactivators cooperating with the estrogen receptor and the retinoic acid receptor. Nucleic Acids Res. 1994;22:1815–20.
Muchardt C, Yaniv M. When the SWI/SNF complex remodels the cell cycle. Oncogene. 2001;20:3067–75.
Park JH, Park EJ, Lee HS, Kim SJ, Hur SK, et al. Mammalian SWI/SNF complexes facilitate DNA double-strand break repair by promoting γ-H2AX induction. EMBO J. 2006;25:3986–97.
Euskirchen GM, Auerbach RK, Davidov E, Gianoulis TA, Zhong G, et al. Diverse roles and interactions of the SWI/SNF chromatin remodeling complex revealed using global approaches. PLoS Genet. 2011;7:e1002008.
Alfert A, Moreno N, Kerl K. The BAF complex in development and disease. Epigenet Chromatin. 2019;12:19.
Wu JI, Lessard J, Crabtree GR. Understanding the words of chromatin regulation. Cell. 2009;136:200–6.
Wang W, Côté J, Xue Y, Zhou S, Khavari PA, et al. Purification and biochemical heterogeneity of the mammalian SWI-SNF complex. EMBO J. 1996;15:5370–82.
Phelan ML, Sif S, Narlikar GJ, Kingston RE. Reconstitution of a core chromatin remodeling complex from SWI/SNF subunits. Mol Cell. 1999;3:247–53.
Mashtalir N, D’Avino AR, Michel BC, Luo J, Pan J, et al. Modular organization and assembly of SWI/SNF family chromatin remodeling complexes. Cell. 2018;175(1272–1288):e20.
Alpsoy A, Dykhuizen EC. Glioma tumor suppressor candidate region gene 1 (GLTSCR15) and its paralog GLTSCR15-like form SWI/SNF chromatin remodeling subcomplexes. J Biol Chem. 2018;293:3892–903.
Gatchalian J, Malik S, Ho J, Lee D-S, Kelso TWR, et al. A non-canonical BRD9-containing BAF chromatin remodeling complex regulates naive pluripotency in mouse embryonic stem cells. Nat Commun. 2018;9:5139.
Lorch Y, Maier-Davis B, Kornberg RD. Mechanism of chromatin remodeling. Proc Natl Acad Sci USA. 2010;107:3458–62.
Narlikar GJ, Sundaramoorthy R, Owen-Hughes T. Mechanisms and functions of ATP-dependent chromatin-remodeling enzymes. Cell. 2013;154:490–503.
Sarnowska EA, Gratkowska DM, Sacharowski SP, Cwiek P, Tohge T, et al. The role of SWI/SNF chromatin remodeling complexes in hormone crosstalk. Trends Plant Sci. 2016;21:594–608.
Huang Z, Li J, Sachs LM, Cole PA, Wong J. A role for cofactor–cofactor and cofactor–histone interactions in targeting p300, SWI/SNF and Mediator for transcription. EMBO J. 2003;22:2146–55.
King HA, Trotter KW, Archer TK. Chromatin remodeling during glucocorticoid receptor regulated transactivation. Biochim Biophys Acta Gene Regul Mech. 2012;1819:716–26.
Sif S, Saurin AJ, Imbalzano AN, Kingston RE. Purification and characterization of mSin3A-containing Brg1 and hBrm chromatin remodeling complexes. Genes Dev. 2001;15:603–18.
Kadam S, Emerson BM. Transcriptional specificity of human SWI/SNF BRG1 and BRM chromatin remodeling complexes. Mol Cell. 2003;11:377–89.
Bultman S, Gebuhr T, Yee D, La Mantia C, Nicholson J, et al. A Brg1 null mutation in the mouse reveals functional differences among mammalian SWI/SNF complexes. Mol Cell. 2000;6:1287–95.
Reyes JC, Barra J, Muchardt C, Camus A, Babinet C, et al. Altered control of cellular proliferation in the absence of mammalian brahma (SNF2α). EMBO J. 1998;17:6979–91.
Glaros S, Cirrincione GM, Muchardt C, Kleer CG, Michael CW, et al. The reversible epigenetic silencing of BRM: implications for clinical targeted therapy. Oncogene. 2007;26:7058–66.
Banerjee R, Bultman SJ, Holley D, Hillhouse C, Bain JR, et al. Non-targeted metabolomics of Brg1/Brm double-mutant cardiomyocytes reveals a novel role for SWI/SNF complexes in metabolic homeostasis. Metabolomics. 2015;11:1287–301.
Sinha S, Verma S, Chaturvedi MM. Differential expression of SWI/SNF chromatin remodeler subunits brahma and brahma-related gene during drug-induced liver injury and regeneration in mouse model. DNA Cell Biol. 2016;35:373–84.
Thompson K, Marquez SB, Lu L, Reisman D. Induction of functional Brm protein from Brm knockout mice. Oncoscience. 2015;2:349.
Yang M, Sun Y, Ma L, Wang C, Wu J, et al. Complex alternative splicing of the Smarca2 gene suggests the importance of Smarca2-B variants. J Cancer. 2011;2:386–400.
Marquez-Vilendrer SB, Rai SK, Gramling SJB, Lu L. Loss of the SWI/SNF ATPase subunits BRM and BRG1 drives lung cancer development. Oncoscience. 2016;3:322–36.
Kobayashi H, Kawahara N, Ogawa K, Yamada Y, Iwai K, et al. Conceptual frameworks of synthetic lethality in clear cell carcinoma of the ovary. Biomed Rep. 2018;9:112–8.
Kosho T, Miyake N, Carey JC. Coffin–Siris syndrome and related disorders involving components of the BAF (mSWI/SNF) complex: historical review and recent advances using next generation sequencing. Am J Med Genet Part C Semin Med Genet. 2014;166:241–51.
Sekiguchi F, Tsurusaki Y, Okamoto N, Teik KW, Mizuno S, et al. Genetic abnormalities in a large cohort of Coffin–Siris syndrome patients. J Hum Genet. 2019;2:1–14.
Wieczorek D, Bögershausen N, Beleggia F, Steiner-Haldenstätt S, Pohl E, et al. A comprehensive molecular study on Coffin–Siris and Nicolaides–Baraitser syndromes identifies a broad molecular and clinical spectrum converging on altered chromatin remodeling. Hum Mol Genet. 2013;22:5121–35.
Tsurusaki Y, Okamoto N, Ohashi H, Kosho T, Imai Y, et al. Mutations affecting components of the SWI/SNF complex cause Coffin–Siris syndrome. Nat Genet. 2012;44:376–8.
Tsurusaki Y, Okamoto N, Ohashi H, Mizuno S, Matsumoto N, et al. Coffin–Siris syndrome is a SWI/SNF complex disorder. Clin Genet. 2014;85:548–54.
Espín-Pérez A, de Kok TMCM, Jennen DGJ, Hendrickx DM, De Coster S, et al. Distinct genotype-dependent differences in transcriptome responses in humans exposed to environmental carcinogens. Carcinogenesis. 2015;36:1154–61.
Raab JR, Runge JS, Spear CC, Magnuson T. Co-regulation of transcription by BRG1 and BRM, two mutually exclusive SWI/SNF ATPase subunits. Epigenet Chromatin. 2017;10:62.
Wu J, He K, Zhang Y, Song J, Shi Z, et al. Inactivation of SMARCA2 by promoter hypermethylation drives lung cancer development. Gene. 2019;687:193–9.
Dunaief JL, Strober BE, Guha S, Khavari PA, Ålin K, et al. The retinoblastoma protein and BRG1 form a complex and cooperate to induce cell cycle arrest. Cell. 1994;79:119–30.
Strober BE, Dunaief JL, Goff SP. Functional interactions between the hBRM/hBRG1 transcriptional activators and the pRB family of proteins. Mol Cell Biol. 1996;16:1576–83.
Trouche D, Le Chalony C, Muchardt C, Yaniv M, Kouzarides T. RB and hbrm cooperate to repress the activation functions of E2F1. Proc Natl Acad Sci USA. 1997;94:11268–73.
Reisman DN, Strobeck MW, Betz BL, Sciariotta J, Funkhouser W Jr, et al. Concomitant down-regulation of BRM and BRG1 in human tumor cell lines: differential effects on RB-mediated growth arrest vs CD44 expression. Oncogene. 2002;21:1196–207.
Roesley SNA, La Marca JE, Deans AJ, Mckenzie L, Suryadinata R, et al. Phosphorylation of Drosophila Brahma on CDK-phosphorylation sites is important for cell cycle regulation and differentiation. Cell Cycle. 2018;17:1559–78.
Zhang HS, Gavin M, Dahiya A, Postigo AA, Ma D, et al. Exit from G1 and S phase of the cell cycle is regulated by repressor complexes containing HDAC-Rb-hSWI/SNF and Rb-hSWI/SNF. Cell. 2000;101:79–89.
Marquez-Vilendrer SB, Rai SK, Gramling SJB, Lu L, Reisman DN. BRG1 and BRM loss selectively impacts RB and P53, respectively: BRG1 and BRM have differential functions in vivo. Oncoscience. 2016;3:337–50.
Lee D, Kim JW, Seo T, Hwang SG, Choi E-J, et al. SWI/SNF complex interacts with tumor suppressor p53 and is necessary for the activation of p53-mediated transcription. J Biol Chem. 2002;277:22330–7.
Xu Y, Zhang J, Chen X. The activity of p53 is differentially regulated by Brm- and Brg1-containing SWI/SNF chromatin remodeling complexes. J Biol Chem. 2007;282:37429–35.
Brownlee PM, Meisenberg C, Downs JA. The SWI/SNF chromatin remodelling complex: its role in maintaining genome stability and preventing tumourigenesis. DNA Repair. 2015;32:127–33.
Ribeiro-Silva C, Aydin ÖZ, Mesquita-Ribeiro R, Slyskova J, Helfricht A, et al. DNA damage sensitivity of SWI/SNF-deficient cells depends on TFIIH subunit p62/GTF2H1. Nat Commun. 2018;9:4067.
Bochar DA, Wang L, Beniya H, Kinev A, Xue Y, et al. BRCA1 is associated with a human SWI/SNF-related complex. Cell. 2000;102:257–65.
Batsché E, Yaniv M, Muchardt C. The human SWI/SNF subunit Brm is a regulator of alternative splicing. Nat Struct Mol Biol. 2006;13:22–9.
Sakurai K, Furukawa C, Haraguchi T, Inada K-I, Shiogama K, et al. MicroRNAs miR-199a-5p and -3p target the Brm subunit of SWI/SNF to generate a double-negative feedback loop in a variety of human cancers. Cancer Res. 2011;71:1680–9.
Kobayashi K, Sakurai K, Hiramatsu H, Inada K, Shiogama K, et al. The miR-199a/Brm/EGR1 axis is a determinant of anchorage-independent growth in epithelial tumor cell lines. Sci Rep. 2015;5:8428.
Zhang Z, Li J, Guo H, Wang F, Ma L, et al. BRM transcriptionally regulates miR-302a-3p to target SOCS5/STAT3 signaling axis to potentiate pancreatic cancer metastasis. Cancer Lett. 2019;449:215–25.
Zhang Z, Wang F, Du C, Guo H, Ma L, et al. BRM/SMARCA2 promotes the proliferation and chemoresistance of pancreatic cancer cells by targeting JAK2/STAT3 signaling. Cancer Lett. 2017;402:213–24.
Link KA, Burd CJ, Williams E, Marshall T, Rosson G, et al. BAF57 governs androgen receptor action and androgen-dependent proliferation through SWI/SNF. Mol Cell Biol. 2005;25:2200–15.
Marshall TW, Link KA, Petre-Draviam CE, Knudsen KE. Differential requirement of SWI/SNF for androgen receptor activity. J Biol Chem. 2003;278:30605–13.
Wang S, Zhang B, Faller DV. BRG1/BRM and prohibitin are required for growth suppression by estrogen antagonists. EMBO J. 2004;23:2293–303.
Kim KH, Roberts CWM. Mechanisms by which SMARCB1 loss drives rhabdoid tumor growth. Cancer Genet. 2014;207:365–72.
Roberts CWM, Biegel JA. The role of SMARCB1/INI1 in development of rhabdoid tumor. Cancer Biol Ther. 2009;8:412–6.
Kahali B, Yu J, Marquez SB, Thompson KW, Liang SY, et al. The silencing of the SWI/SNF subunit and anticancer gene BRM in Rhabdoid tumors. Oncotarget. 2014;5:3316–32.
Herpel E, Rieker RJ, Dienemann H, Muley T, Meister M, et al. SMARCA4 and SMARCA2 deficiency in non–small cell lung cancer: immunohistochemical survey of 316 consecutive specimens. Ann Diagn Pathol. 2017;26:47–51.
Matsubara D, Kishaba Y, Ishikawa S, Sakatani T, Oguni S, et al. Lung cancer with loss of BRG1/BRM, shows epithelial mesenchymal transition phenotype and distinct histologic and genetic features. Cancer Sci. 2013;104:266–73.
Yoshimoto T, Matsubara D, Nakano T, Tamura T, Endo S, et al. Frequent loss of the expression of multiple subunits of the SWI/SNF complex in large cell carcinoma and pleomorphic carcinoma of the lung. Pathol Int. 2015;65:595–602.
Fukuoka J, Fujii T, Shih JH, Dracheva T, Meerzaman D, et al. Chromatin remodeling factors and BRM/BRG1 expression as prognostic indicators in non-small cell lung cancer. Clin Cancer Res. 2004;10:4314–24.
Liu G, Gramling SJB, Munoz D, Cheng D, Azad AK, et al. Two novel BRM insertion promoter sequence variants are associated with loss of BRM expression and lung cancer risk. Oncogene. 2011;30:3295–304.
Zernickel E, Sak A, Riaz A, Klein D, Groneberg M, et al. Targeting of BRM sensitizes BRG1 -mutant lung cancer cell lines to radiotherapy. Mol Cancer Ther. 2019;18:656–66.
Sarnowska EA, Szymanski M, Rusetska N, Ligaj M, Jancewicz I, et al. Evaluation of the role of downregulation of SNF5/INI1 core subunit of SWI/SNF complex in clear cell renal cell carcinoma development. Am J Cancer Res. 2017;7:2275.
The Cancer Genome Atlas Network. Comprehensive molecular characterization of clear cell renal cell carcinoma. Nature. 2013;499:43–9.
Xia Q-Y, Rao Q, Cheng L, Shen Q, Shi S-S, et al. Loss of BRM expression is a frequently observed event in poorly differentiated clear cell renal cell carcinoma. Histopathology. 2014;64:847–62.
Agaimy A, Amin MB, Gill AJ, Popp B, Reis A, et al. SWI/SNF protein expression status in fumarate hydratase–deficient renal cell carcinoma: immunohistochemical analysis of 32 tumors from 28 patients. Hum Pathol. 2018;77:139–46.
Xia Q-Y, Zhan X-M, Fan X-S, Ye S-B, Shi S-S, et al. BRM/SMARCA2-negative clear cell renal cell carcinoma is associated with a high percentage of BRM somatic mutations, deletions and promoter methylation. Histopathology. 2016;70:711–21.
Agaimy A, Daum O, Markl B, Lichtmannegger I, Michal M, et al. SWI/SNF complex-deficient undifferentiated/rhabdoid carcinomas of the gastrointestinal tract: a series of 13 cases highlighting mutually exclusive loss of SMARCA4 and SMARCA2 and frequent co-inactivation of SMARCB1 and SMARCA2. Am J Surg Pathol. 2016;40:544–53.
Yu Y, Cheng D, Parfrey P, Liu G, Savas S. Two functional indel polymorphisms in the promoter region of the Brahma gene (BRM) and disease risk and progression-free survival in colorectal cancer. PLoS ONE. 2018;13:1–17.
Wong KM, Qiu X, Cheng D, Azad AKALA, Habbous S, et al. Two BRM promoter insertion polymorphisms increase the risk of early-stage upper aerodigestive tract cancers. Cancer Med. 2014;3:426–33.
Yamamichi N, Inada KI, Ichinose M, Yamamichi-Nishina M, Mizutani T, et al. Frequent loss of Brm expression in gastric cancer correlates with histologic features and differentiation state. Cancer Res. 2007;67:10727–35.
Takeshima H, Niwa T, Takahashi T, Wakabayashi M, Yamashita S, et al. Frequent involvement of chromatin remodeler alterations in gastric field cancerization. Cancer Lett. 2015;357:328–38.
Numata M, Morinaga S, Watanabe T, Tamagawa H, Yamamoto N, et al. The clinical significance of SWI/SNF complex in pancreatic cancer. Int J Oncol. 2013;42:403–10.
Shain AH, Giacomini CP, Matsukuma K, Karikari CA, Bashyam MD, et al. Convergent structural alterations define SWItch/Sucrose Non-Fermentable (SWI/SNF) chromatin remodeler as a central tumor suppressive complex in pancreatic cancer. Proc Natl Acad Sci. 2012;109:E252–9.
Endo M, Yasui K, Zen Y, Gen Y, Zen K, et al. Alterations of the SWI/SNF chromatin remodelling subunit-BRG1 and BRM in hepatocellular carcinoma. Liver Int. 2013;33:105–17.
Pasic I, Wong KM, Lee JJ, Espin-Garcia O, Brhane Y, et al. Two BRM promoter polymorphisms predict poor survival in patients with hepatocellular carcinoma. Mol Carcinog. 2018;57:106–13.
Wang JR, Gramling SJB, Goldstein DP, Cheng D, Chen D, et al. Association of two BRM promoter polymorphisms with head and neck squamous cell carcinoma risk. Carcinogenesis. 2013;34:1012–7.
Ho AS, Kannan K, Roy DM, Morris LGT, Ganly I, et al. The mutational landscape of adenoid cystic carcinoma. Nat Genet. 2013;45:791–8.
Jagielska B, Sarnowska E, Rusetska N, Jancewicz I, Durzynska M, et al. Advanced adenoid cystic carcinoma (ACC) is featured by SWI/SNF chromatin remodeling complex aberrations. J Cancer Res Clin Oncol. 2019;145:201–11.
Cohet N, Stewart KM, Mudhasani R, Asirvatham AJ, Mallappa C, et al. SWI/SNF chromatin remodeling enzyme ATPases promote cell proliferation in normal mammary epithelial cells. J Cell Physiol. 2010;223:667–78.
Wu Q, Madany P, Akech J, Dobson JR, Douthwright S, et al. The SWI/SNF ATPases are required for triple negative breast cancer cell proliferation. J Cell Physiol. 2015;230:2683–94.
Guerrero-Martínez JA, Reyes JC. High expression of SMARCA4 or SMARCA2 is frequently associated with an opposite prognosis in cancer. Sci Rep. 2018;8:1–17.
Yang Y, Liu L, Fang M, Bai H, Xu Y. The chromatin remodeling protein BRM regulates the transcription of tight junction proteins: implication in breast cancer metastasis. Biochim Biophys Acta Gene Regul Mech. 2019;1862:547–56.
Cho H, Kim JS-Y, Chung H, Perry C, Lee H, et al. Loss of ARID1A/BAF250a expression is linked to tumor progression and adverse prognosis in cervical cancer. Hum Pathol. 2013;44:1365–74.
Serber DW, Rogala A, Makarem M, Rosson GB, Simin K, et al. The BRG1 chromatin remodeler protects against ovarian cysts, uterine tumors, and mammary tumors in a lineage-specific manner. PLoS ONE. 2012;7:1–10.
Guan B, Wang TL, Shih IM. ARID1A, a factor that promotes formation of SWI/SNF-mediated chromatin remodeling, is a tumor suppressor in gynecologic cancers. Cancer Res. 2011;71:6718–27.
Shadeo A, Chari R, Lonergan KM, Pusic A, Miller D, et al. Up regulation in gene expression of chromatin remodelling factors in cervical intraepithelial neoplasia. BMC Genomics. 2008;9:64.
Lin DI, Chudnovsky Y, Duggan B, Zajchowski D, Greenbowe J, et al. Comprehensive genomic profiling reveals inactivating SMARCA4 mutations and low tumor mutational burden in small cell carcinoma of the ovary, hypercalcemic-type. Gynecol Oncol. 2017;147:626–33.
Witkowski L, Carrot-Zhang J, Albrecht S, Fahiminiya S, Hamel N, et al. Germline and somatic SMARCA4 mutations characterize small cell carcinoma of the ovary, hypercalcemic type. Nat Genet. 2014;46:438–43.
Ramos P, Karnezis AN, Craig DW, Sekulic A, Russell ML, et al. Small cell carcinoma of the ovary, hypercalcemic type, displays frequent inactivating germline and somatic mutations in SMARCA4. Nat Genet. 2014;46:427–9.
Jelinic P, Mueller JJ, Olvera N, Dao F, Scott SN, et al. Recurrent SMARCA4 mutations in small cell carcinoma of the ovary. Nat Genet. 2014;46:424–6.
Kupryjańczyk J, Dansonka-Mieszkowska A, Moes-Sosnowska J, Plisiecka-Hałasa J, Szafron Ł, et al. Ovarian small cell carcinoma of hypercalcemic type—evidence of germline origin and smarca4 gene inactivation. A pilot study. Polish J Pathol. 2013;64:238–46.
Karnezis AN, Wang Y, Ramos P, Hendricks WPD, Oliva E, et al. Dual loss of the SWI/SNF complex ATPases SMARCA4/BRG1 and SMARCA2/BRM is highly sensitive and specific for small cell carcinoma of the ovary, hypercalcaemic type. J Pathol. 2016;238:389–400.
Jelinic P, Schlappe BA, Conlon N, Tseng J, Olvera N, et al. Concomitant loss of SMARCA2 and SMARCA4 expression in small cell carcinoma of the ovary, hypercalcemic type. Mod Pathol. 2016;29:60–6.
Itamochi H, Oishi T, Oumi N, Takeuchi S, Yoshihara K, et al. Whole-genome sequencing revealed novel prognostic biomarkers and promising targets for therapy of ovarian clear cell carcinoma. Br J Cancer. 2017;117:717–24.
Xu X, Zheng Z, Jia L, Suo S, Liu B, et al. Overexpression of SMARCA2 or CAMK2D is associated with cisplatin resistance in human epithelial ovarian cancer. Oncol Lett. 2018;16:3796–804.
Kothandapani A, Gopalakrishnan K, Kahali B, Reisman D, Patrick SM. Downregulation of SWI/SNF chromatin remodeling factor subunits modulates cisplatin cytotoxicity. Exp Cell Res. 2012;318:1973–86.
Ramalingam P, Croce S, McCluggage WG. Loss of expression of SMARCA4 (BRG1), SMARCA2 (BRM) and SMARCB1 (INI1) in undifferentiated carcinoma of the endometrium is not uncommon and is not always associated with rhabdoid morphology. Histopathology. 2017;70:359–66.
Gustavsson H, Jennbacken K, Welén K, Damber J-E. Altered expression of genes regulating angiogenesis in experimental androgen-independent prostate cancer. Prostate. 2008;68:161–70.
Shen H, Powers N, Saini N, Comstock CESS, Sharma A, et al. The SWI/SNF ATPase Brm is a gatekeeper of proliferative control in prostate cancer. Cancer Res. 2008;68:10154–62.
Bock VL, Lyons JG, Huang XXJ, Jones AM, McDonald LA, et al. BRM and BRG1 subunits of the SWI/SNF chromatin remodelling complex are downregulated upon progression of benign skin lesions into invasive tumours. Br J Dermatol. 2011;164:1221–7.
Moloney FJ, Lyons JG, Bock VL, Huang XX, Bugeja MJ, et al. Hotspot mutation of Brahma in non-melanoma skin cancer. J Invest Dermatol. 2009;129:1012–5.
Farrell AW, Halliday GM, Lyons JG. Brahma deficiency in keratinocytes promotes UV carcinogenesis by accelerating the escape from cell cycle arrest and the formation of DNA photolesions. J Dermatol Sci. 2018;92:254–63.
Vachtenheim J, Ondrušová L, Borovanský J. SWI/SNF chromatin remodeling complex is critical for the expression of microphthalmia-associated transcription factor in melanoma cells. Biochem Biophys Res Commun. 2010;392:454–9.
Buscarlet M, Krasteva V, Ho L, Simon C, Hebert J, et al. Essential role of BRG, the ATPase subunit of BAF chromatin remodeling complexes, in leukemia maintenance. Blood. 2014;123:1720–8.
Doménech E, Gómez-López G, Gzlez-Peña D, López M, Herreros B, et al. New mutations in chronic lymphocytic leukemia identified by target enrichment and deep sequencing. PLoS ONE. 2012;7:2–7.
Eisfeld A-K, Kohlschmidt J, Mrózek K, Volinia S, Blachly JS, et al. mutational landscape and gene expression patterns in adult acute myeloid leukemias with monosomy 7 as a sole abnormality. Cancer Res. 2017;77:207–18.
Li L, Fan XS, Xia QY, Rao Q, Liu B, et al. Concurrent loss of INI1, PBRM1, and BRM expression in epithelioid sarcoma: implications for the cocontributions of multiple SWI/SNF complex members to pathogenesis. Hum Pathol. 2014;45:2247–54.
Amankwah EK, Thompson RC, Nabors LB, Olson JJ, Browning JE, et al. SWI/SNF gene variants and glioma risk and outcome. Cancer Epidemiol. 2013;37:162–5.
Kadoch C, Hargreaves DC, Hodges C, Elias L, Ho L, et al. Proteomic and bioinformatic analysis of mammalian SWI/SNF complexes identifies extensive roles in human malignancy. Nat Genet. 2013;45:592–601.
Gramling S, Rogers C, Liu G, Reisman D. Pharmacologic reversal of epigenetic silencing of the anticancer protein BRM: a novel targeted treatment strategy. Oncogene. 2011;30:3289–94.
Januario T, Ye X, Bainer R, Alicke B, Smith T, et al. PRC2-mediated repression of SMARCA2 predicts EZH2 inhibitor activity in SWI/SNF mutant tumors. Proc Natl Acad Sci. 2017;114:12249–54.
Bourachot B, Yaniv M, Muchardt C. Growth inhibition by the mammalian SWI-SNF subunit Brm is regulated by acetylation. EMBO J. 2003;22:6505–15.
Marquez SB, Thompson KW, Lu L, Reisman D. Beyond mutations: additional mechanisms and implications of SWI/SNF complex inactivation. Front Oncol. 2015;4:1–20.
Ouyang X, Ye XL, Wei HB. BRM promoter insertion polymorphisms increase the risk of cancer: a meta-analysis. Gene. 2017;626:420–5.
Segedi M, Anderson LN, Espin-Garcia O, Borgida A, Bianco T, et al. BRM polymorphisms, pancreatic cancer risk and survival. Int J Cancer. 2016;139:2474–81.
La Rochelle J, Klatte T, Dastane A, Rao N, Seligson D, et al. Chromosome 9p deletions identify an aggressive phenotype of clear cell renal cell carcinoma. Cancer. 2010;116:4696–702.
Gramling SJB, Reisman D. Discovery of BRM targeted therapies: novel reactivation of an anticancer gene. Lett Drug Des Discov. 2011;8:93–9.
Oike T, Ogiwara H, Tominaga Y, Ito K, Ando O, et al. A synthetic lethality-based strategy to treat cancers harboring a genetic deficiency in the chromatin remodeling factor BRG1. Cancer Res. 2013;73:5508–18.
Papillon JPN, Nakajima K, Adair CD, Hempel J, Jouk AO, et al. Discovery of orally active inhibitors of brahma homolog (BRM)/SMARCA2 ATPase activity for the treatment of brahma related gene 1 (BRG1)/SMARCA4-mutant cancers. J Med Chem. 2018;61:10155–72.
Farnaby W, Koegl M, Roy MJ, Whitworth C, Diers E, et al. BAF complex vulnerabilities in cancer demonstrated via structure-based PROTAC design. Nat Chem Biol. 2019;15:672–80.
Reisman DN, Glaros S, Thompson EA. The SWI/SNF complex and cancer. Oncogene. 2009;28:1653–68.
Yamamichi N, Yamamichi-Nishina M, Mizutani T, Watanabe H, Minoguchi S, et al. The Brm gene suppressed at the post-transcriptional level in various human cell lines is inducible by transient HDAC inhibitor treatment, which exhibits antioncogenic potential. Oncogene. 2005;24:5471–81.
Kahali B, Gramling SJB, Marquez SB, Thompson KW, Lu L, et al. Identifying targets for the restoration and reactivation of BRM. Oncogene. 2014;33:653–64.
Chan-Penebre E, Armstrong K, Drew A, Grassian AR, Feldman I, et al. Selective killing of SMARCA2- and SMARCA4-deficient small cell carcinoma of the ovary, hypercalcemic type cells by inhibition of EZH2: in vitro and in vivo preclinical models. Mol Cancer Ther. 2017;16:850–60.
Wang Y, Chen SY, Colborne S, Lambert G, Shin CY, et al. Histone deacetylase inhibitors synergize with catalytic inhibitors of EZH2 to exhibit antitumor activity in small cell carcinoma of the ovary, hypercalcemic type. Mol Cancer Ther. 2018;17:2767–79.
O’Neil NJ, Bailey ML, Hieter P. Synthetic lethality and cancer. Nat Rev Genet. 2017;18:613–23.
Hoffman GR, Rahal R, Buxton F, Xiang K, McAllister G, et al. Functional epigenetics approach identifies BRM/SMARCA2 as a critical synthetic lethal target in BRG1-deficient cancers. Proc Natl Acad Sci. 2014;111:3128–33.
Pascolini G, Valiante M, Bottillo I, Laino L, Fleischer N, et al. Striking phenotypic overlap between Nicolaides–Baraitser and Coffin–Siris syndromes in monozygotic twins with ARID1B intragenic deletion. Eur J Med Genet. 2019;14:103739.
Dai Y, Ngo D, Jacob J, Forman LW, Faller DV. Prohibitin and the SWI/SNF ATPase subunit BRG1 are required for effective androgen antagonist-mediated transcriptional repression of androgen receptor-regulated genes. Carcinogenesis. 2008;29:1725–33.
Lempiäinen JK, Niskanen EA, Vuoti K-M, Lampinen RE, Göös H, et al. Agonist-specific protein interactomes of glucocorticoid and androgen receptor as revealed by proximity mapping. Mol Cell Proteomics. 2017;16:1462–74.
van de Wijngaart DJ, Dubbink HJ, Molier M, de Vos C, Trapman J, et al. Functional screening of FxxLF-like peptide motifs identifies SMARCD1/BAF60a as an androgen receptor cofactor that modulates TMPRSS2 expression. Mol Endocrinol. 2009;23:1776–86.
Hill DA, de la Serna IL, Veal TM, Imbalzano AN. BRCA1 interacts with dominant negative SWI/SNF enzymes without affecting homologous recombination or radiation-induced gene activation of p21 or Mdm2. J Cell Biochem. 2004;91:987–98.
Fontana GA, Rigamonti A, Lenzken SC, Filosa G, Alvarez R, et al. Oxidative stress controls the choice of alternative last exons via a Brahma-BRCA1-CstF pathway. Nucleic Acids Res. 2017;45:902–14.
Kondo T, Raff M. Chromatin remodeling and histone modification in the conversion of oligodendrocyte precursors to neural stem cells. Genes Dev. 2004;18:2963–72.
Wang G-L, Iakova P, Wilde M, Awad S, Timchenko NA. Liver tumors escape negative control of proliferation via PI3 K/Akt-mediated block of C/EBP alpha growth inhibitory activity. Genes Dev. 2004;18:912–25.
Lavigne M, Eskeland R, Azebi S, Saint-André V, Jang SM, et al. Interaction of HP1 and Brg1/Brm with the globular domain of histone H3 is required for HP1-mediated repression. PLoS Genet. 2009;5:e1000769.
Nozawa R-S, Nagao K, Masuda H-T, Iwasaki O, Hirota T, et al. Human POGZ modulates dissociation of HP1α from mitotic chromosome arms through Aurora B activation. Nat Cell Biol. 2010;12:719.
Steunou A-L, Ducoux-Petit M, Lazar I, Monsarrat B, Erard M, et al. Identification of the hypoxia-inducible factor 2α nuclear interactome in melanoma cells reveals master proteins involved in melanoma development. Mol Cell Proteomics. 2013;12:736–48.
Zhang Y, Cheng M, Zhang Y, Zhong X, Dai H, et al. A switch from hBrm to Brg1 at IFNγ-activated sequences mediates the activation of human genes. Cell Res. 2010;20:1345–60.
Ichinose H, Garnier J-M, Chambon P, Losson R. Ligand-dependent interaction between the estrogen receptor and the human homologues of SWI2/SNF2. Gene. 1997;188:95–100.
Jiang X-X, Nguyen Q, Chou Y, Wang T, Nandakumar V, et al. Control of B cell development by the histone H2A deubiquitinase MYSM1. Immunity. 2011;35:883–96.
Nakamura T, Mori T, Tada S, Krajewski W, Rozovskaia T, et al. ALL-1 Is a histone methyltransferase that assembles a supercomplex of proteins involved in transcriptional regulation. Mol Cell. 2002;10:1119–28.
Li J, Xi Y, Li W, McCarthy RL, Stratton SA, et al. TRIM28 interacts with EZH2 and SWI/SNF to activate genes that promote mammosphere formation. Oncogene. 2017;36:2991–3001.
Yamamichi N, Inada K, Furukawa C, Sakurai K, Tando T, et al. Cdx2 and the Brm-type SWI/SNF complex cooperatively regulate villin expression in gastrointestinal cells. Exp Cell Res. 2009;315:1779–89.
Wang F, Zhang R, Beischlag TV, Muchardt C, Yaniv M, et al. Roles of Brahma and Brahma/SWI2-related gene 1 in hypoxic induction of the erythropoietin gene. J Biol Chem. 2004;279:46733–41.
Secombe J, Li L, Carlos L, Eisenman RN. The Trithorax group protein Lid is a trimethyl histone H3K4 demethylase required for dMyc-induced cell growth. Genes Dev. 2007;21:537–51.
Havugimana PC, Hart GT, Nepusz T, Yang H, Turinsky AL, et al. A census of human soluble protein complexes. Cell. 2012;150:1068–81.
Pal S, Yun R, Datta A, Lacomis L, Erdjument-Bromage H, et al. mSin3A/histone deacetylase 2- and PRMT5-containing Brg1 complex is involved in transcriptional repression of the Myc target gene cad. Mol Cell Biol. 2003;23:7475–87.
Cao Q, Wang X, Zhao M, Yang R, Malik R, et al. The central role of EED in the orchestration of polycomb group complexes. Nat Commun. 2014;5:3127.
Kowenz-Leutz E, Leutz A. A C/EBPβ Isoform Recruits the SWI/SNF Complex to Activate Myeloid Genes. Mol Cell. 1999;4:735–43.
Ito T, Yamauchi M, Nishina M, Yamamichi N, Mizutani T, et al. Identification of SWI.SNF complex subunit BAF60a as a determinant of the transactivation potential of Fos/Jun dimers. J Biol Chem. 2001;276:2852–7.
Kim BR, Coyaud E, Laurent EMN, St-Germain J, Van de Laar E, et al. Identification of the SOX2 interactome by BioID reveals EP300 as a mediator of SOX2-dependent squamous differentiation and lung squamous cell carcinoma growth. Mol Cell Proteomics. 2017;16:1864–88.
Cox JL, Wilder PJ, Gilmore JM, Wuebben EL, Washburn MP, et al. The SOX2-interactome in brain cancer cells identifies the requirement of MSI2 and USP9X for the growth of brain tumor cells. PLoS ONE. 2013;8:e62857.
Cho H, Orphanides G, Sun X, Yang XJ, Ogryzko V, et al. A human RNA polymerase II complex containing factors that modify chromatin structure. Mol Cell Biol. 1998;18:5355–63.
Sauter JL, Graham RP, Larsen BT, Jenkins SM, Roden AC, et al. SMARCA4-deficient thoracic sarcoma: a distinctive clinicopathological entity with undifferentiated rhabdoid morphology and aggressive behavior. Mod Pathol. 2017;30:1422–32.
Bookhout C, Bouldin TW, Ellison DW. Atypical teratoid/rhabdoid tumor with retained INI1 (SMARCB1) expression and loss of BRG1 (SMARCA4). Neuropathology. 2018;38:305–8.
Mei L, Alikhan M, Mujacic I, Parilla M, Antic T. Genomic alterations in undifferentiated malignant tumors with rhabdoid phenotype and loss of BRG1 immunoexpression identified by fine needle aspirates. Acta Cytol. 2019;63:438–44.
Orvis T, Hepperla A, Walter V, Song S, Simon J, et al. BRG1/SMARCA4 inactivation promotes non-small cell lung cancer aggressiveness by altering chromatin organization. Cancer Res. 2014;74:6486–98.
Medina PP, Romero OA, Kohno T, Montuenga LM, Pio R, et al. Frequent BRG1/SMARCA4-inactivating mutations in human lung cancer cell lines. Hum Mutat. 2008;29:617–22.
Rao Q, Xia Q-Y, Shen Q, Shi S-S, Tu P, et al. Coexistent loss of INI1 and BRG1 expression in a rhabdoid renal cell carcinoma (RCC): implications for a possible role of SWI/SNF complex in the pathogenesis of RCC. Int J Clin Exp Pathol. 2014;7:1782–7.
Sentani K, Oue N, Kondo H, Kuraoka K, Motoshita J, et al. Increased expression but not genetic alteration of < i>BRG1 </i > a component of the SWI/SNF complex, is associated with the advanced stage of human gastric carcinomas. Pathobiology. 2001;69:315–20.
Gunduz E, Gunduz M, Ouchida M, Nagatsuka H, Beder L, et al. Genetic and epigenetic alterations of BRG1 promote oral cancer development. Int J Oncol. 2005;26:201–10.
Bai J, Mei P, Zhang C, Chen F, Li C, et al. BRG1 is a prognostic marker and potential therapeutic target in human breast cancer. PLoS ONE. 2013;8:1–9.
Lin H, Wong RPC, Martinka M, Li G. BRG1 expression is increased in human cutaneous melanoma. Br J Dermatol. 2010;163:502–10.
Saladi S, Keenen B, Marathe HG, Qi H, Chin K-V, et al. Modulation of extracellular matrix/adhesion molecule expression by BRG1 is associated with increased melanoma invasiveness. Mol Cancer. 2010;9:280.
Acknowledgements
We thank Richard Bayliss and Sharon Yeoh for critical comments to the manuscript and correction of English language.
Funding
This work was supported by the Polish Ministry of Science and Higher Education, under Diamond Grant no. 0229/DIA/2015/44 (IJ) any by the National Science Center Grant no. UMO-2016/23/N/NZ1/01138 (IJ) and Polpharma Scientific Foundation 5/XVII/18 (TJS).
Author information
Authors and Affiliations
Contributions
IJ wrote the main part of the manuscript. ES, JAS and TJS made a substantive revision of the text and added essential paragraphs and conclusions. All the authors corrected and approved the final version of the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing of interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Jancewicz, I., Siedlecki, J.A., Sarnowski, T.J. et al. BRM: the core ATPase subunit of SWI/SNF chromatin-remodelling complex—a tumour suppressor or tumour-promoting factor?. Epigenetics & Chromatin 12, 68 (2019). https://doi.org/10.1186/s13072-019-0315-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13072-019-0315-4