
Abstract
Background
In Panama, arboviroses such as dengue fever, and more recently chikungunya fever and Zika disease, are transmitted by Aedes aegypti and Aedes albopictus. Their control is based on the elimination of breeding sites and fogging with pyrethroid insecticides. However, one of the significant issues derived from the prolonged use of pyrethroid insecticide is the development of resistance mechanisms, such as knockdown resistance or kdr. The objective of this study was to evaluate the presence of kdr mutations in a partial region of the VGSC gene in samples of wild-caught Aedes mosquitoes from different locations of the Metropolitan Region of Panama.
Results
Based on the analysis of 194 sequences of the VGSC gene, two kdr mutations (Ile1011Met and Val1016Gly) were detected in a specimen of Ae. aegypti. The frequency of kdr mutations in the evaluated samples of Ae. aegypti was 0.01.
Conclusions
This study provides evidence for a low frequency of kdr mutations in Ae. aegypti populations in Panama. It is possible that these changes have no impact on vector control interventions. To our knowledge, we report, for the first time in America the Val1016Gly mutation documented in Asia. In general terms, this result is highly relevant to the Aedes Control Programme in Panama since it constitutes a feasible approach for the timely detection of resistance as well as for the development of strategies.
Similar content being viewed by others


Background
Aedes aegypti (Linnaeus) and Aedes albopictus (Skuse) are the two most important mosquito species in terms of the transmission of infectious diseases [1, 2]. They are vectors of different arboviruses (viruses transmitted by arthropods) of worldwide relevance that include dengue virus (DENV), yellow fever virus (YFV), chikungunya virus (CHIKV) and Zika virus (ZIKV) [3]. Both of these species of the order Diptera are considered invasive since they have shown success in colonizing many regions outside their endemic areas [2].
Currently, control activities are based on surveillance, chemical application and the elimination of mosquito breeding sites. The pyrethroid insecticides such as deltamethrin and cyfluthrin have been, until now, the most commonly used class of insecticide for vector control against the annual epidemics of diseases transmitted by Aedes in Panama [4, 5]. The rotation in the application of insecticides has also been carried out; deltamethrin has been applied to health regions that have the highest infestation rates and cyfluthrin in the remaining regions. However, even if outbreaks are controlled, concerns about the effect of the continuous use of these insecticides on the populations of Ae. aegypti and Ae. albopictus persist, specifically in relation to the development of resistance mechanisms, which is one of the main problems faced by chemical control programmes [5,6,7].
In Panama, populations of mosquito vectors have been subjected to a continuous selective pressure of organochlorine, carbamate, organophosphate and pyrethroid insecticides. The application of these chemical compounds began in 1957 through the creation of the National Service for the Eradication of Malaria (Servicio Nacional de Erradicación de la Malaria, SNEM) [8, 9]. However, activities for the control of Aedes populations officially began in the year 1969, through the Campaign of Eradication of Ae. aegypti developed by the Ministry of Health. During the 1980s, this was renamed Campaign for the Control of Ae. aegypti and, subsequently, MINSA integrated the SNEM with the Programme of Ae. aegypti and created the Department of Vector Control. Since 1969, temephos (Abate) has been used for the control of larvae (focal treatment) of Aedes. From the 1970s until 1985, malathion spraying was used as a perifocal treatment for the control of adult mosquito populations. From 1985 to the present, spraying with the pyrethroid deltamethrin has been applied [10]. From 1992 to 2016, spraying with cyfluthrin was applied in the regions with the lowest risk of transmission; however, its use was suspended due to supply problems.
Resistance is due to two main mechanisms: a greater metabolic detoxification of chemical agents and insensitivity at the site of action [11]. The mechanisms involving overexpression or qualitative changes in catalytic enzyme sites include non-specific esterases (NSE), glutathione S-transferases (GSTs) and mixed function oxidases (MFOs) [12]. However, knockdown resistance, or kdr, is one of the main types of resistance against pyrethroid insecticides [13]. It is caused by point mutations at the level of the nucleotide sequence of the para gene and leads to changes in some amino acids of the voltage-gated sodium channel (VGSC) protein, which causes a reduction of the binding with the insecticide and, consequently, the loss of its effect [14,15,16,17].
This study aimed to evaluate the resistance in Aedes vectors through molecular tools, given that previous studies with populations of Ae. aegypti have shown metabolic resistance, and its mechanisms have been characterized for different types of insecticides [18, 19]. Furthermore, there is no previous information about the presence of kdr alleles in the studied Aedes populations in Panama.
The study is part of a more comprehensive investigation that includes the performance of susceptibility bioassays standardized by the WHO, and the use of synergistic agents and biochemical tests for the detection of enzymatic mechanisms associated with resistance to insecticides. Each of these approaches satisfies the need of the Aedes Control Programme of MINSA to understand the behavior of resistance and its mechanisms in populations of these vectors.
Methods
Sampling
The sampling locations were selected based on the criteria of a high incidence of dengue and levels of infestation with Aedes mosquitoes from high to moderate [20]. Five locations of the Metropolitan Region of Panama were selected: Nuevo Chorrillo (8°57'36.09"N, 79°41'54.48"W), Princess Mía (8°58'1.29"N, 79°42'8.92"W), Lluvia de Oro (8°57'36.57"N, 79°41'56.28"W), Bethania (9°0'34.04"N, 79°31'45.95"W) and Las Garzas (9°7'6.00"N, 79°15'47.32"W). In each locality, 25–30 houses were selected at random.
The capture of Aedes mosquitoes and eggs was carried out during the months of August to November 2015. For this purpose, two trap types were used: a Mosquito Science Trap-N-Kill™ ovitrap (SpringStar Inc., Woodinville, WA, USA) and a a BG-Sentinel® Trap (Biogents AG, Regensburg, Germany), which were placed in the peridomicile of the homes. Within each BG-Sentinel Trap, BG-Lure™ (Biogents AG, Regensburg, Germany) was used as an attractant to favor the capture of adult mosquitoes. These traps were monitored daily to change the battery and the catch bag, the latter of which was transported to the laboratory in a portable ice cooler in order to preserve specimens for taxonomic identification and subsequent molecular analyses. In the case of the Trap-N-Kill™ ovitrap, paddles were picked up at the end of the week and transported to the laboratory in Ziploc® bags (SC Johnson, Racine, WI, USA), to evaluate the presence of eggs. The paddles with eggs were immersed in trays of water and were reared to adult stage.
Identification of sex and species was carried out with taxonomic keys [21]. Subsequently, the mosquitoes were placed in 1.5 ml conical tubes with 500 μl of DNA/RNA Shield™ (Zymo Research, Irvine, CA, USA) and stored at -80 °C.
DNA isolation
The genomic DNA of the mosquitoes was isolated with a ZR Viral DNA/RNA Kit™ (Zymo Research) according to the manufacturer’s instructions. The quality and concentration of the isolated DNA were evaluated with a NanoDrop™ 2000c spectrophotometer (Thermo Scientific, Wilmington, DE, USA). Finally, samples were labeled and stored at -80 °C.
PCR amplification
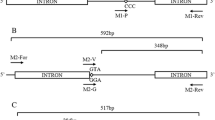
To identify the presence of kdr mutations in Ae. aegypti and Ae. albopictus, the previously proposed oligonucleotides AaSCF1, AaSCR4, AaSCF7 and AaSCR7 were used [22, 23]. These oligonucleotides allow the identification of five kdr sites within the VGSC sequence (Table 1).
The mixture for the PCR was prepared with PCR Master Mix 2× (Promega, Madison, WI, USA) following the manufacturer’s specifications. The amplification process was carried out in a Mastercycler® gradient thermocycler (Eppendorf, Hamburg, Germany) based on a previously described protocol [22]. The quality and integrity of the PCR products were evaluated by agarose gel electrophoresis (AMRESCO, Solon, OH, USA), prepared at 1.5% and stained with Gel red® (Biotium, Fremont, CA, USA). Fragments of approximately 800 bp were obtained with the oligonucleotides AaSCF1 and AaSCR4. Fragments of approximately 700 bp were obtained with the oligonucleotides AaSCF7 and AaSCR7. The samples amplified by PCR were subsequently stored at -20 °C.
Sequencing
The sequencing reactions were performed from the PCR products using the oligonucleotides AaSCF3, AaSCR6 and AaSCR8 [22, 23]. Sequencing was undertaken by Macrogen (Seoul, Korea), and the direct sequencing of the samples was performed on an ABI 3730XL genetic analyzer (Applied Biosystems, Foster City, CA, USA).
Analysis of results
The chromatograms were visualized with BioEdit v.7.2.5 software [24]; the sequences obtained in ABI format was transformed to FASTA format for editing. The authenticity of the sequences was corroborated through the BLAST program (https://blast.ncbi.nlm.nih.gov/Blast.cgi), considering an identity percentage greater than 95%. The sequences were aligned and edited with MEGA 7.0 software [25]. To calculate the frequency of the kdr mutations, we considered the number of samples that showed point changes in the kdr sites of interest as a function of the total sequences analyzed.
Results
A total of 3432 specimens of Ae. aegypti and 593 specimens of Ae. albopictus were collected. Of these, 149 specimens were sequenced, of which 74 individuals were Ae. aegypti and 75 were Ae. albopictus. From these, 447 sequences were obtained, of which only 194 were of sufficient quality to include in the analysis for detecting mutations at the level of Domain II of the VGSC protein. The kdr sites analyzed were Ser989, Ile1011, Leu1014, Val1016 and Phe1534, resulting in two point mutations (Ile1011Met and Val1016Gly) identified in a single specimen of Ae. aegypti from the locality of Nuevo Chorrillo. Table 2 shows the number of samples evaluated and positive samples in Aedes spp.
The multiple alignments of the partial nucleotide sequence of the para gene in Ae. aegypti are presented, specifically from exons 20 and 21 in Figs. 1 and 2, respectively. These present the contrast of some of the sequences analyzed in the study with reference sequences obtained from the GenBank database.

Partial section of the nucleotide sequence of DIIS6 of the VGSC protein (para gene) in Ae. aegypti (exon 20). Samples Ae079, Ae060 and Ae051 are contrasted with reference sequences Ref. 1 (GenBank: AB914690.1) and Ref. 2 (GenBank: FJ479612.1)

Partial section of the nucleotide sequence of DIIS6 of the VGSC protein (para gene) in Ae. aegypti (exon 21). Samples Ae079, Ae090 and Ae371 are contrasted with reference sequences Ref. 1 (GenBank: AB914690.1) and Ref. 2 (GenBank: AB914689.1)
In sample Ae079, the change in the amino acid isoleucine (Ile) by the amino acid methionine (Met) was observed, which is the product of a transition in the third base of the codon 1011; the non-mutated codon is ATA and the mutated codon is ATG (Additional files 1 and 2).
At position 1016 of sample Ae079, the change of the amino acid valine (Val) to the amino acid glycine (Gly) was observed, a product of a transversion occurring in the second nucleotide of the codon; the non-mutated codon is GTA and the mutated codon is GGT (Additional files 1 and 2). It is important to note that at the level of the third nucleotide there is also a point mutation (A → T), but it is synonymous.
The frequency of kdr mutations in the samples of Ae. aegypti evaluated in this study was 0.01, which corresponds to 1% of the total samples, and 0.06, which corresponds to 6% of the samples from Nuevo Chorrillo (Table 3).
Discussion
Recent reports suggest that the populations of Ae. aegypti in Panama are, in general terms, susceptible to commercial pesticides. Larval resistance bioassays have shown the existence of vector populations that have moderate levels of resistance to the pyrethroid deltamethrin [8]. These results are significant given that, in the absence of adequate management by the vector control programme, the potential for the dissemination of this phenotypic trait may be increased in the future. However, the detection of point mutations Ile1011Met and Val1016Gly in a single specimen of Ae. aegypti using molecular tools demonstrates the importance and feasibility of incorporating molecular techniques for the detection and monitoring of resistance to insecticides in mosquitoes. This approach also complements the results of the bioassays and improves control strategies.
It is important to point out that the detection of mutation Val1016Gly raises new questions about its distribution and introduction at the regional level since, to our knowledge, it is the first time that is reported in America. In fact, the Val1016Ile and Val1016Gly mutations have distinct geographical distributions, with Val1016Gly found in Asia [26,27,28,29,30] and Val1016Ile in the Americas and recently detected in Africa [14, 26, 27, 31,32,33,34]. Based on these results, we hypothesize that the Val1016Gly mutation could have been introduced to Panama via the transit of containers or tires with eggs because the country is a site of commercial and international transit, which is difficult to monitor and control. At first, we hypothesized that the mutation was introduced in America but there is also the possibility that it is a new mutation. If it is a new mutation, an exhaustive phylogeographical analysis will establish the origin of the mutation present in our region.
In the present study, both the Ile1011Met and Val1016Gly mutations were detected in a sample of Ae. aegypti from the same locality (NC). The co-occurrence of kdr mutations in Ae. aegypti has been reported previously, specifically of the Val1016 and Ser989 mutations in Asian populations of this vector [26, 27]. However, the implications of the combination of kdr mutations are debated; some studies report that their co-occurrence improves resistance [35], but others conclude that there is no additive or synergistic effect [36].
In this study it was not possible to detect kdr mutations in Ae. albopictus sequences, a fact that may be related to technical factors (due to the quality of some sequences obtained) which we consider part of the limitations of this study. It is important to note that of the 194 evaluated sequences, only 20 were found to have sufficient quality to identify point changes at the DIIIS6 level of the VGSC, a region where it has been possible to characterize the Phe1534Cys mutation. Notably, a study conducted in Costa Rica [22] did not detect mutant alleles associated with kdr resistance in a natural population of Ae. albopictus, a characteristic attributed to the recent invasion of this species in that country. The colonization of Ae. albopictus populations in Panama is relatively recent [37] compared to Ae. aegypti [38]; therefore, the failure to detect kdr mutations in Ae. albopictus may be because chemical control with insecticides has not yet put pressure on the populations of this vector. However, in Ae. albopictus populations from other latitudes, it has been possible to detect the Phe1534Cys mutation. For example, in Singapore, Kasai et al. [39] found that 92.3% of mosquitoes exhibited the Phe1534Cys mutation as detected by sequencing, thus estimating a frequency of 73.1% for the Cys1534 allele. Recently, the evaluation of Ae. albopictus populations from Asia, Africa, America and Europe [40] detected two new kdr mutations at the level of domain III of the VGSC, namely mutations Ile1532Thr and Phe1534Ser, the latter presenting a significant association with resistance to deltamethrin.
The frequency of kdr mutations in the analyzed samples was low (1%) compared to that reported in studies conducted in the region. For example, in populations of Ae. aegypti from Grand Cayman and Cuba, frequencies of 79% and 51%, respectively, for the kdr allele Ile1016 were detected through sequencing [41, 42]. Similar results are reported for populations of Ae. aegypti from Venezuela evaluated through the allele-specific PCR technique (PCR-AS) [43] and from Brazil [44]. Another study carried out in Brazil [31] revealed patterns of regional distribution of kdr mutations attributed to positions Val1016 and Phe1534 in Ae. aegypti collected over ten years. According to the authors, the regionalization of the kdr alleles reflects differences in the populations of Ae. aegypti that colonized the continent.
Lastly, we consider that the low frequency detected in our study does not yet have an impact on mosquito control interventions. Determining the distribution of the Ile1011Met and Val1016Gly mutations, as well as other kdr mutations in populations of Ae. aegypti or in other species of mosquito vectors present in Panama, requires a greater sampling effort and an adaptation of the methodology used in this research. The information generated will be of great value in determining the frequency of the mutant alleles.
Conclusions
This study provides evidence for a low frequency of kdr mutations (Ile1011Met and Val1016Gly) in Ae. aegypti populations in Panama. The low frequency recorded is perhaps not enough to have an impact on the interventions of mosquito control. To our knowledge, we report, for the first time in America the Val1016Gly mutation documented in Asia. The finding of the kdr mutations in specimens not previously exposed to resistance bioassays is indicative that the natural populations of this vector could be developing resistance to the insecticides that are being applied in Panama. In general terms, the information on the presence of this kdr mutation in Panama can help monitor the spread of the mutation in America in the case that it becomes a significant problem for vector control. This result is highly relevant to the Aedes Control Programme in Panama since it constitutes a feasible approach for the timely detection of resistance as well as for the development of strategies.
Abbreviations
- A:
-
Adenine
- C:
-
Cytosine
- CHIKV:
-
Chikungunya virus
- DENV:
-
Dengue virus
- G:
-
Guanine
- kdr :
-
Knockdown resistance
- MINSA:
-
Ministerio de Salud (Ministry of Health)
- PCR:
-
Polymerase chain reaction
- PCR-AS:
-
Polymerase chain reaction allele-specific
- T:
-
Thymine
- VGSC:
-
Voltage-gated sodium channel
- YFV:
-
Yellow fever virus
- ZIKV:
-
Zika virus
References
Figueroa DP, Scott S, Hamilton-West C, González CR, Canals M. Mosquitoes: disease vectors in context of climate change in Chile. Parasitol Latinoam. 2015;64:30–40.
Rey JR, Lounibos P. Ecología de Aedes aegypti y Aedes albopictus en América y la transmisión de enfermedades. Biomédica. 2015;35:177–85.
Kraemer MU, Sinka ME, Duda KA, Mylne AQ, Shearer FM, Barker CM, et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. Elife. 2015;4:e08347.
Carrera J-P, Díaz Y, Denis B, Barahona de Mosca I, Rodriguez D, Cedeño I, et al. Unusual pattern of chikungunya virus epidemic in the Americas, the Panamanian experience. PLoS Negl Trop Dis. 2017;11:e0005338.
Valderrama A, Chaves LF, Futami K. Evaluation of kdr mutations in Aedes albopictus (Skuse) in Panama. (Preliminary data). Rev Med Panama. 2016;36:30–2.
David J-P, Ismail HM, Chandor-Proust A, Paine MJI. Role of cytochrome P450s in insecticide resistance: impact on the control of mosquito-borne diseases and use of insecticides on Earth. Philos Trans R Soc Lond B Biol Sci. 2013;368:20120429.
Smith LB, Kasai S, Scott JG. Pyrethroid resistance in Aedes aegypti and Aedes albopictus: Important mosquito vectors of human diseases. Pestic Biochem Phys. 2016;133:1–12.
Cáceres L, Rovira J, García A, Torres R, Cruz MD. Determinación de la sensibilidad a insecticidas organofosforados, carbamato y piretroides en poblaciones de Aedes aegypti Linnaeus, 1762 (Díptera: Culicidae) de Panamá. Biomédica. 2013;33:70–81.
Cáceres L. La lucha antimalárica y el desarrollo de la resistencia del Anopheles albimanus a los insecticidas en Panamá. 1st ed. Panamá: Poligrafía SA; 1999.
Organización Panamericana de la Salud. Guías para el abordaje integral del dengue en Panamá, 2014. Panamá: 1st ed. Panamá: Editora Sibauste SA; 2014.
Labbé P, Alout H, Djogbénou L, Pasteur N, Weill M. Evolution of resistance to insecticide in disease vectors. In: Tibayrenc M, editor. Genetics and Evolution of Infectious Diseases. London: Elsevier; 2011. p. 363–409.
Bisset JA, Rodríguez MM, French L, Severson DW, Gutiérrez G, Hurtado D, et al. Insecticide resistance and metabolic mechanisms involved in larval and adult stages of Aedes aegypti insecticide-resistant reference strains from Cuba. J Am Mosq Control Assoc. 2014;30:298–304.
Aguirre-Obando OA, Bona ACD, Duque JE, Navarro-Silva MA. Insecticide resistance and genetic variability in natural populations of Aedes (Stegomyia) aegypti (Diptera: Culicidae) from Colombia. Zoologia. 2015;32:14–22.
Alvarez LC, Ponce G, Saavedra-Rodriguez K, Lopez B, Flores AE. Frequency of V1016I and F1534C mutations in the voltage-gated sodium channel gene in Aedes aegypti in Venezuela. Pest Manag Sci. 2015;71:863–9.
Dong K, Du Y, Rinkevich F, Nomura Y, Xu P, Wang L, et al. Molecular biology of insect sodium channels and pyrethroid resistance. Insect Biochem Mol Biol. 2014;50:1–17.
Donnelly MJ, Corbel V, Weetman D, Wilding CS, Williamson MS, Black WC IV. Does kdr genotype predict insecticide-resistance phenotype in mosquitoes? Trends Parasitol. 2009;25:213–9.
Hemingway J, Ranson H. Insecticide resistance in insect vectors of human disease. Annu Rev Entomol. 2000;45:371–91.
Bisset JA, Rodríguez MM, Cáceres L. Niveles de resistencia a insecticidas y sus mecanismos en 2 cepas de Aedes aegypti de Panamá. Rev Cubana Med Trop. 2003;55:191–5.
Cáceres L. Determinación de la resistencia a insecticidas y sus mecanismos en poblaciones de Aedes aegypti (Díptera: Culicidae) en algunos países América Central. Thesis. Instituto de Medicina Tropical Pedro Kourí, Cuba; 2013. http://tesis.repo.sld.cu/681/.
Ministerio de Salud. Boletín epidemiológico No 34: Dengue, 2015. 2015. http://www.minsa.gob.pa/informacion-salud/boletines-semanales-2015. Accessed 7 Nov 2016.
Clark-Gil S, Darsie RF. The mosquitoes of Guatemala. Mosq Syst. 1983;13:151–284.
Chaves LF, Kawashima E, Futami K, Minakawa N, Rodriguez RM. Lack of kdr mutations in a population of Asian tiger mosquitoes from Costa Rica. B Insectol. 2015;68:61–3.
Kawada H, Higa Y, Komagata O, Kasai S, Tomita T, Thi Yen N, et al. Widespread distribution of a newly found point mutation in voltage-gated sodium channel in pyrethroid-resistant Aedes aegypti populations in Vietnam. PLoS Negl Trop Dis. 2009;3:e0000527.
Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl Acids Symp. Ser. 1999;41:95–8.
Kumar S, Stecher G, Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for bigger datasets. Mol Biol Evol. 2016;33:1870–4.
Moyes CL, Vontas J, Martins AJ, Ng LC, Koou SY, Dusfour I, et al. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS Negl Trop Dis. 2017;11:e0005625.
Kawada H, Higa Y, Futami K, Muranami Y, Kawashima E, Osei JHN, et al. Discovery of point mutations in the voltage-gated sodium channel from African Aedes aegypti populations: potential phylogenetic reasons for gene introgression. PLoS Negl Trop Dis. 2016;10:e0004780.
Ishak IH, Jaal Z, Ranson H, Wondji CS. Contrasting patterns of insecticide resistance and knockdown resistance (kdr) in the dengue vectors Aedes aegypti and Aedes albopictus from Malaysia. Parasit Vectors. 2015;8:181.
Pang SC, Chiang LP, Tan CH, Vythilingam I, Lam-Phua SG, Ng LC. Low efficacy of deltamethrin treated net against Singapore Aedes aegypti is associated with kdr-type resistance. Trop Biomed. 2015;32:140–50.
Stenhouse SA, Plernsub S, Yanola J, Lumjuan N, Dantrakool A, Choochote W, et al. Detection of the V1016G mutation in the voltage-gated sodium channel gene of Aedes aegypti (Diptera: Culicidae) by allele-specific PCR assay, and its distribution and effect on deltamethrin resistance in Thailand. Parasit Vectors. 2013;6:253.
Linss JG, Brito LP, Garcia GA, Araki AS, Bruno RV, Lima JB, et al. Distribution and dissemination of the Val1016Ile and Phe1534Cys kdr mutations in Aedes aegypti Brazilian natural populations. Parasit Vectors. 2014;7:25.
Saavedra-Rodriguez K, Urdaneta-Marquez L, Rajatileka S, Moulton M, Flores AE, Fernandez-Salas I, et al. A mutation in the voltage-gated sodium channel gene associated with pyrethroid resistance in Latin American Aedes aegypti. Insect Mol Biol. 2007;16:785–98.
Marcombe S, Mathieu RB, Pocquet N, Riaz M-A, Poupardin R, Sélior S, et al. Insecticide resistance in the dengue vector Aedes aegypti from Martinique: distribution, mechanisms and relations with environmental factors. PLoS One. 2012;7:e30989.
Martins AJ, Lima JB, Peixoto AA, Valle D. Frequency of Val1016Ile mutation in the voltage-gated sodium channel gene of Aedes aegypti Brazilian populations. Trop Med Int Health. 2009;14:1351–5.
Srisawat R, Komalamisra N, Eshita Y, Zheng M, Ono K, Itoh TQ, et al. Point mutations in domain II of the voltage-gated sodium channel gene in deltamethrin-resistant Aedes aegypti (Diptera: Culicidae). Appl Entomol Zool. 2010;45:275–82.
Du Y, Nomura Y, Satar G, Hu Z, Nauen R, He SY, et al. Molecular evidence for dual pyrethroid-receptor sites on a mosquito sodium channel. Proc Natl Acad Sci USA. 2013;110:11785–90.
Futami K, Valderrama A, Baldi M, Minakawa N, Rodríguez RM, Chaves LF. New and common haplotypes shape genetic diversity in Asian tiger mosquito populations from Costa Rica and Panamá. J Econ Entomol. 2015;108:761–8.
Powell JR, Tabachnick WJ. History of domestication and spread of Aedes aegypti - a review. Mem Inst Oswaldo Cruz. 2013;108(Suppl. 1):11–7.
Kasai S, Ng LC, Lam-Phua SG, Tang CS, Itokawa K, Komagata O, et al. First detection of a putative knockdown resistance gene in major mosquito vector, Aedes albopictus. Jpn J Infect Dis. 2011;64:217–21.
Xu J, Bonizzoni M, Zhong D, Zhou G, Cai S, Li Y, et al. Multi-country survey revealed prevalent and novel F1534S mutation in voltage-gated sodium channel (VGSC) gene in Aedes albopictus. PLoS Negl Trop Dis. 2016;10:e0004696.
Bariami V, Jones CM, Poupardin R, Vontas J, Ranson H. Gene amplification, ABC transporters and cytochrome p450s: unraveling the molecular basis of pyrethroid resistance in the dengue vector, Aedes aegypti. PLoS Negl Trop Dis. 2012;6:e1692.
Harris AF, Rajatileka S, Ranson H. Pyrethroid resistance in Aedes aegypti from Grand Cayman. Am J Trop Med Hyg. 2010;83:277–84.
Álvarez González L, Ponce García G, Oviedo M, Briceño A, Flores Suarez AE. Mecanismos asociados a la resistencia al derribo “kdr” a la deltametrina en Aedes aegypti del occidente de Venezuela. Bol Malariol Salud Ambient. 2014;54:58–67.
Aguirre-Obando OA, Pietrobon AJ, Bona ACD, Navarro-Silva MA. Contrasting patterns of insecticide resistance and knockdown resistance (kdr) in Aedes aegypti populations from Jacarezinho (Brazil) after a dengue outbreak. Rev Bras Entomol. 2016;60:94–100.
Acknowledgments
We thank the staff of the Department of Research in Medical Entomology (Departamento de Investigación en Entomología Médica, DIEM) of the Gorgas Memorial Institute of Health Studies (Instituto Conmemorativo Gorgas de Estudios de la Salud, ICGES). We also thank the Secretaría Nacional de Ciencia y Tecnología e Innovación (SENACYT) for funding the Masters studies of OM and Sistema Nacional de Investigadores. We are grateful the Department of Vector Control of the Ministry of Health, particularly Galindo Ruíz and José Lasso, for their valuable collaboration.
Funding
Financial support was provided by the project “The Panama Dengue Fever Research” to Sandra Lopez Vergés (PI). Secretaría Nacional de Ciencia y Tecnología e Innovación (SENACYT), through the Sistema Nacional de Investigación (SNI), supported research activities by AV and LC. Financial Support was also provided by the Panama Science and Innovation Fund 2015 grant (UK).
Availability of data and materials
Data supporting the conclusions of this article are included within the article. The newly generated sequences are provided in Additional files 1 and 2.
Author information
Authors and Affiliations
Contributions
AV and OM conceived and participated in coordinating the study. LC, JY and AV designed the samplings. SK, JY, RM and DP performed the samplings and species identification. BH, AC and OM carried out the molecular methods. OM and AV performed the sequence alignment and molecular data interpretation. OM and AV interpreted results and drafted the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1:
Alignment of the newly generated sequences. Partial section of the nucleotide sequence of DIIS6 of the VGSC protein (para gene) in Ae. aegypti (exon 20). (FAS 0.608)
Additional file 2:
Alignment of the newly generated sequences. Partial section of the nucleotide sequence of the DIIS6 of the VGSC protein (para gene) in Ae. aegypti (exon 21). (FAS 0.243)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Murcia, O., Henríquez, B., Castro, A. et al. Presence of the point mutations Val1016Gly in the voltage-gated sodium channel detected in a single mosquito from Panama. Parasites Vectors 12, 62 (2019). https://doi.org/10.1186/s13071-019-3309-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-019-3309-y