
Abstract
Background
In Brazil, studies on animals and humans in mainland areas have shown that most strains of Toxoplasma gondii are pathogenic to mice and exhibit great genetic variability.
Results
In this study, using a set of 11 PCR-RFLP and 15 microsatellite markers, we isolated and genetically characterised T. gondii strains from one cat and three rats on Fernando de Noronha Island. The cat had antibodies to T. gondii, which were revealed using a modified agglutination test (MAT, cut-off 1:25) and the seroprevalence among the 46 rodents was 15.2%. Viable T. gondii was isolated from one cat (TgCatBrFN1), two brown rats (TgRatnoBrFN1 and TgRatnoBrFN2) and one black rat (TgRatraBrFN1). Unlike the strains from mainland Brazil, these isolates were not pathogenic to outbred mice. The genotypes of these strains were compared with strains previously isolated on the island and in mainland Brazil. The analysis based on microsatellite data showed a limited genetic diversity of T. gondii on Fernando de Noronha Island with the majority of strains clustered into the following three groups: type II, III, and Caribbean 1.
Conclusions
There was little variation among strains within the same group, suggesting that the majority of strains circulating on Fernando de Noronha are derived from only a few strains that were recently introduced to the island, likely from imported cats. Except for the strain belonging to the Caribbean 1 group that originates from northeast Brazil, there was little evidence that strains from the other groups were introduced to Fernando de Noronha via mainland Brazil.
Similar content being viewed by others

Background
Toxoplasma gondii infections are prevalent in animals and humans on a global level [1]. Felids are key species in the life-cycle of T. gondii because sexual reproduction in their intestines leads to the production of millions of highly resistant oocysts [2]. The seroprevalence of T. gondii among humans is high in Brazil, reaching 90% in some regions, and may be related to high environmental contamination by oocysts [3]. The worldwide genetic diversity of T. gondii isolates has been studied extensively over the last two decades, showing that the hotspot of diversity is located in South America, particularly in Brazil [4]. Severe cases of ocular and congenital toxoplasmosis in Brazil have been associated with this high diversity [5].
Fernando de Noronha (3°50'28.9''S, 32°24'39.4''W) is an archipelago of 21 islands and islets in the Atlantic Ocean and is located approximately 354 km east of the Brazilian coast. The main island has a population of approximately 3,000 inhabitants [6]. There are no records regarding the juncture of when cats and rats were introduced to the island, but these animals may have come from Europe on ships that arrived on the archipelago, starting at the time of its first human occupation during the sixteenth century.
Previous studies reported high rates of T. gondii seroprevalence in different animal species in Fernando de Noronha, including cats, suggesting that transmission of this zoonotic parasite is active among the animal fauna of this Brazilian archipelago [7,8,9]. Toxoplasma gondii strains isolated from chickens, cats, and cattle egrets on Fernando de Noronha showed unexpected genotyping results. These results showed the presence of atypical genotypes that seemed endemic to the island as well as clonal type II strains that are common in Europe and North America but are virtually absent in mainland Brazil [3, 9, 10]. The objective of this study was to isolate and characterise T. gondii strains from one cat and several rodents on Fernando de Noronha with PCR-FLP and microsatellite markers and examine their genetic relationships with other strains previously isolated on the island and in mainland Brazil.
Methods
Sample collection
On October 5, 2013, an adult male cat with a terminal condition was admitted by its owner to the Animal Surveillance Centre (NVA) of the State District Administration of Fernando de Noronha in the village of Vila do Sueste. The cat was necropsied soon after death, and the heart, brain and thigh muscle samples were collected. A blood sample was obtained from the cardiac cavity and was centrifuged at 1,500× g for 10 min; the serum sample was stored at -20 °C.
Between October 2013 and April 2014, as part of a synanthropic rodent control programme on the Island of Fernando de Noronha, 28 black rats (Rattus rattus) and 18 brown rats (Rattus norvegicus) were captured with Tomahawk live traps at different locations on the island. The rodents were anaesthetised using ketamine hydrochloride (40–90 mg/kg) and xylazine hydrochloride (2–5 mg/kg) and blood samples (3 ml) were collected by means of cardiac puncture. The rodents were individually sacrificed in a hermetically sealed box containing cotton wool soaked with isoflurane. Necropsies were performed to collect brain, heart, and skeletal muscle samples from each rodent.
All cat and rodent samples were transported under refrigeration by air to the Faculty of Veterinary Medicine of University of São Paulo (FMVZ-USP) for laboratory analyses.
Serological examination and bioassay
The serum samples from the cat and black and brown rats were tested using the modified agglutination test (MAT) [11], with a 1:25 dilution as the cut-off [12].
To conduct the bioassay, brain, heart and skeletal muscle samples from each rodent were homogenised together in a blender with 0.85% NaCl, whereas each cat tissue sample was homogenised separately. The homogenates from each animal were digested with an acidic pepsin solution [13] and subcutaneously inoculated into five or three outbred Swiss mice (1.0 ml per mouse). The surviving mice were bled six weeks post-inoculation (p.i.), and a 1:25 dilution of the serum from each mouse was tested for T. gondii antibodies with MAT. Mice were euthanised (using the same protocol as above) two months p.i., and their brains were examined for T. gondii tissue cysts [1].
Genotyping analyses and neighbour-joining clustering
DNA samples from T. gondii strains collected in this study were extracted from mouse brains with the Qiagen® Dneasy® Blood & Tissue kit and genotyped using 11 PCR-RFLP and 15 microsatellite markers distributed on the 14 chromosomes and plastid of T. gondii, as described previously [14,15,16]. As shown in Additional file 1: Table S1, for comparison, we included the genotyping data of 24 strains previously collected on Fernando de Noronha Island [3, 10] and 41 strains isolated from two different regions of mainland Brazil described in other studies (14 strains from northeast Brazil [10, 17,18,19,20] and 27 from São Paulo [15, 21]). The ENT, ME49, and NED strains were used as reference T. gondii type I, II, and III strains, respectively [22].
Because microsatellite markers are better markers than PCR-RFLP for discriminating related isolates at a limited geographic scale such as an island [14], the neighbour-joining tree was only reconstructed from microsatellite data. The unrooted tree was reconstructed with Populations 1.2.32 (http://bioinformatics.org/populations/) based on the chord-distance Dc evaluation of Cavalli-Sforza and Edwards [23] and generated with MEGA version 6.05 [24] (http://www.megasoftware.net/history.php).
Results
Antibodies to T. gondii were found in the cat, in five out of 28 black rats (18%), and in two out of 18 brown rats (11%). The MAT titres were as follows: < 25 (39 rodents), 25 (one black rat), 100 (cat), 800 (two black rats), 1,600 (two black rats and one brown rat), and 6,400 (one brown rat) (Table 1). Viable T. gondii strains were isolated from the cat, two brown rats and one black rat, and the isolates were named TgCatBrFN1, TgRatnoBrFN1, TgRatnoBrFN2 and TgRatraBrFN1, respectively (Fig. 1). None of the isolates were pathogenic in mice (Table 1).

Map of Fernando de Noronha Island, Brazil, showing the locations where Toxoplasma gondii isolates were obtained. The asterisk indicates T. gondii isolates from chickens genotyped previously [3]
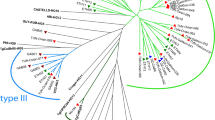
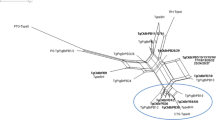
The genotyping results with RFLP and microsatellite markers are presented in Additional file 1: Table S1. The neighbour-joining analysis based on microsatellite data showed that all T. gondii strains but one (TgCkBr220) from Fernando de Noronha Island were highly structured in three groups: three strains (including TgCatBrFN1 and TgRatnoBrFN1 from this study) were clustered with the ME49 strain in the type II group, six strains (including TgRatnoBrFN2 from this study) were clustered with the NED strain in the type III group, and the TgRatRaBrFN1 strain was grouped in the Caribbean 1 group (Fig. 2). Inside each group, differences among strains were limited even with polymorphic microsatellite markers. For example, only one microsatellite marker was able to differentiate strains with the ToxoDB-RFLP genotype #146 in the type III group, even though they were collected from different hosts and at different times and locations on the island (Additional file 1: Table S1). Strains with the ToxoDB-RFLP genotype #3 that belong to the type II group were also very similar, with only three microsatellite markers able to differentiate them, and had the allele 329 at the microsatellite marker N83 (Additional file 1: Table S1). This latter allele is absent in strains from Europe and North America and, to date, has been identified only in Brazilian strains (D. Ajzenberg, personal communication). Strains from common clonal lineages in mainland Brazil (BrI and BrII) were not identified in Fernando de Noronha.

Neighbour-joining clustering of Toxoplasma gondii strains based on 15 microsatellite markers. Squares indicate type I (ENT), II (ME49) and III (NED) reference strains; red points indicate strains from Fernando de Noronha Island, yellow points indicate strains from São Paulo State; blue points indicate strains from northeast Brazil; arrows indicate the four strains collected in the present study. Strains TgCkBr221, 225, 226, 228 and 230 had 15 identical microsatellite markers and were represented by the TgCkBr221 strain in the tree (see Additional file 1: Table S1 for genotyping details); TgCkBr211, TgCkBr183 and TgCkBr210 were identical to TgCkBr224, TgCkBr184 and TgCkBr212-219, 223, 227 and 229, respectively
Discussion
The Island of Fernando de Noronha provides a unique opportunity to study the epidemiology of toxoplasmosis because of restricted animal and human populations. There is a high density of cats (Felis catus) in Fernando de Noronha: of the estimated 848 domestic animals living on the island, 470 are domestic cats [25]. We are not aware of the presence of any wild felid on the island. The number of cats seems to be more important than just their presence because as the number of cats increases, the number of oocysts in the environment increases [1, 26]. Since rats are considered an important food source for cats, we surveyed T. gondii infection in these intermediate hosts. All the rats were bioassayed, irrespective of serological findings, because congenitally infected rats may develop immune tolerance without antibody production [27]. Toxoplasma gondii antibodies were found in 15% of the 46 rats collected in our study, which is lower than the 38% [7] of black rats from Fernando de Noronha with the same test and cut-off; these data confirm that rats are significantly infected with T. gondii and contribute to the cycle of this zoonosis on the island.
Our results confirm the limited genetic diversity of T. gondii strains on the island. The minor variation between strains in the same group with polymorphic microsatellite markers suggests that the majority of T. gondii strains from Fernando de Noronha originated from a few strains that were recently introduced onto the island, likely from imported cats or possibly migratory birds. The TgRatraBrFN1 strain belongs to the Caribbean 1 group that is endemic to the Caribbean and the anthropised coast of French Guiana [28, 29]. In this study, we show that this Caribbean 1 group is also endemic to northeast Brazil, where the TgRatraBrFN1 strain likely originates. The ToxoDB-RFLP genotype #146 seems to be the dominant genotype on the island since it has been isolated from chickens, cats, rats, and even cattle egrets living on the island in prior studies [3, 9, 10] and this study. The neighbour-joining analysis with microsatellite markers indicates that this genotype is related to clonal type III strains, but the fact that this genotype has only been sampled to date on the island [4] and its significant divergence from type III strains with RFLP markers suggests a specific evolutionary history for this genotype. Type II strains have now been isolated from chickens, cats and rats from Fernando de Noronha, which confirms that the most successful clonal lineage from Europe and North America is circulating in Fernando de Noronha. The origin of this genotype on the island remains enigmatic because it is virtually absent from mainland Brazil except in the southernmost part of the country near the Argentinean and Uruguayan borders [30], and the presence of allele 129 at the microsatellite marker N83 excludes importation from Europe or North America.
Conclusions
The State District Administration of Fernando de Noronha (ADEFN) has a programme to promote public health and control synanthropic animals. Our results support the implementation of a toxoplasmosis prevention and control programme based on the guidelines of One Health and Ecological Health (conservation medicine) in the archipelago of Fernando de Noronha.
Abbreviations
- ADEFN:
-
State District Administration of Fernando de Noronha
- FMVZ-USP:
-
Faculty of Veterinary Medicine of University of São Paulo
- MAT:
-
Modified agglutination test
- NVA:
-
Animal surveillance centre
- PCR-RFLP:
-
Polymerase chain reaction - restriction fragment length polymorphism
References
Dubey JP. Toxoplasmosis of animals and humans. 2nd ed. Boca Raton: CRC Press; 2010. p. 338.
Dubey JP, Lappin MR, Thulliez P. Long-term antibody responses of cats fed Toxoplasma gondii tissue cysts. J Parasitol. 1995;81:887–93.
Dubey JP, Rajendran C, Costa DGC, Ferreira LR, Kwok OCH, Qu D, et al. New Toxoplasma gondii genotypes isolated from free-range chickens from the Fernando de Noronha, Brazil: unexpected findings. J Parasitol. 2010;96:709–12.
Shwab EK, Zhu X-Q, Majumdar D, Pena HFJ, Gennari SM, Dubey JP, Su C. Geographical patterns of Toxoplasma gondii genetic diversity revealed by multilocus PCR-RFLP genotyping. Parasitol. 2014;141:453–61.
Gilbert RE, Freeman K, Lago EG, Bahia-Oliveira LMG, Tan HK, Wallon M, et al. Ocular sequelae of congenital toxoplasmosis in Brazil compared with Europe. PLoS Negl Trop Dis. 2008;2(8):E277.
IBGE. Instituto Brasileiro de Geografia e Estatística. Estimativa da População. Brasília: IBGE; 2015.
Costa DGC, Marvulo MFV, Silva JSA, Santana SC, Magalhães FJR, Lima Filho CDF, et al. Seroprevalence of Toxoplasma gondii in domestic and wild animals from the Fernando de Noronha, Brazil. J Parasitol. 2012;98:679–80.
Magalhães FJR, Silva JG, Ribeiro-Andrade M, Pinheiro JW, Mota RA. High prevalence of toxoplasmosis in free-range chicken of the Fernando de Noronha Archipelago, Brazil. Acta Trop. 2016;159:58–61.
Melo RPB, Almeida JC, Lima DCV, Pedrosa CM, Magalhães FJR, Alcântara AM, et al. Atypical Toxoplasma gondii genotype in feral cats from the Fernando de Noronha Island, northeastern Brazil. Vet Parasitol. 2016;224:92–5.
Vitaliano SN, Soares HS, Minervino AHH, Santos ALQ, Werther K, Marvulo MFV, et al. Genetic characterization of Toxoplasma gondii from Brazilian wildlife revealed abundant new genotypes. Int J Parasitol Parasites Wildl. 2014;3:276–83.
Dubey JP, Desmonts G. Serological responses of equids fed Toxoplasma gondii oocysts. Equine Vet J. 1987;19:337–9.
DeFeo ML, Dubey JP, Mather TN, Rhodes R. Epidemiologic investigation of seroprevalence of antibodies to Toxoplasma gondii in cats and rodents. Am J Vet Res. 2002;63:1714–7.
Dubey JP. Refinement of pepsin digestion method for isolation of Toxoplasma gondii from infected tissues. Vet Parasitol. 1998;74:75–7.
Ajzenberg D, Collinet F, Mercier A, Vignoles P, Dardé ML. Genotyping of Toxoplasma gondii isolates with 15 microsatellite markers in a single multiplex PCR assay. J Clin Microbiol. 2010;48:4641–5.
Pena HFJ, Gennari SM, Dubey JP, Su C. Population structure and mouse-virulence of Toxoplasma gondii in Brazil. Int J Parasitol. 2008;38:561–9.
Su C, Shwab EK, Zhou P, Zhu XQ, Dubey JP. Moving towards an integrated approach to molecular detection and identification of Toxoplasma gondii. Parasitol. 2010;137:1–11.
Dubey JP, Velmurugan GV, Chockalingam A, Pena HFJ, Nunes-Oliveira L, Leifer CA, et al. Genetic diversity of Toxoplasma gondii isolates from chickens from Brazil. Vet Parasitol. 2008;157:299–305.
Gennari SM, Esmerini PO, Lopes MG, Soares HS, Vitaliano SN, Cabral AD, et al. Occurrence of antibodies against Toxoplasma gondii and its isolation and genotyping in donkeys, mules, and horses in Brazil. Vet Parasitol. 2015;209:129–32.
Pena HFJ, Marvulo MFV, Horta MC, Silva MA, Silva JCR, Siqueira DB, et al. Isolation and genetic characterisation of Toxoplasma gondii from a red-handed howler monkey (Alouatta belzebul), a jaguarundi (Puma yagouaroundi), and a black-eared opossum (Didelphis aurita) from Brazil. Vet Parasitol. 2011;175:377–81.
Ragozo AMA, Pena HFJ, Yai LEO, Su C, Gennari SM. Genetic diversity among Toxoplasma gondii isolates of small ruminants from Brazil: Novel genotypes revealed. Vet Parasitol. 2010;170:307–12.
Cabral AD, Gama AR, Sodré MM, Savani ESMM, Galvão-Dias MA, Jordão LR, et al. First isolation and genotyping of Toxoplasma gondii from bats (Mammalia: Chiroptera). Vet Parasitol. 2013;193:100–4.
Ajzenberg D, Cogné N, Paris L, Bessières MH, Thulliez P, Filisetti D, et al. Genotype of 86 Toxoplasma gondii isolates associated with human congenital toxoplasmosis, and correlation with clinical findings. J Infect Dis. 2002;186:684–9.
Cavalli-Sforza LL, Edwards AW. Phylogenetic analysis. Models and estimation procedures. Am J Hum Genet. 1967;19:233–57.
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol. 2013;30:2725–9.
IBGE. Instituto Brasileiro de Geografia e Estatística. Censo demográfico. Brasília: IBGE; 2010.
Weigel RM, Dubey JP, Siegel AM. Risk factors for transmission of Toxoplasma gondii on swine farms in Illinois. J Parasitol. 1995;81:736–41.
Dubey JP, Shen SK, Kwok OCH, Thulliez P. Toxoplasmosis in rats (Rattus norvegicus): congenital transmission to first and second generation offspring and isolation of Toxoplasma gondii from seronegative rats. Parasitol. 1997;115:9–14.
Ajzenberg D, Lamaury I, Demar M, Vautrin C, Cabié A, Simon S, et al. Performance testing of PCR assay in blood samples for the diagnosis of toxoplasmic encephalitis in AIDS patients from the French Departments of America and genetic diversity of Toxoplasma gondii: a prospective and multicentric study. PLoS Negl Trop Dis. 2016. doi:10.1371/journal.pntd.0004790.
Mercier A, Ajzenberg D, Devillard S, Demar MP, Thoisy B, Bonnabau H, et al. Human impact on genetic diversity of Toxoplasma gondii: example of the anthropized environment from French Guiana. Infect Genet Evol. 2011;11:1378–87.
da Silva RC, Langoni H, Su C, Silva AV. Genotypic characterization of Toxoplasma gondii in sheep from Brazilian slaughterhouses: New atypical genotypes and the clonal type II strain identified. Vet Parasitol. 2011;175:173–7.
Acknowledgements
The authors are grateful to Paulo Rogerio Mangini; Tatiane Micheletti Ribeiro Silva; Fernando Ferreira da Silva Filho; Daniel Paulo de Oliveira; Ricardo Araújo; Policarpo Tertuliano Felipe; Carina Abreu; Eduardo Cavalcante de Macedo; and the Administration of Fernando de Noronha, state district, Pernambuco state and Instituto Chico Mendes de Conservação da Biodiversidade – ICMBio.
Funding
This work was supported in part by scholarships from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Brazil (JCRS, RAD, LCA, RAM, SMG, DA and FF), and by a departmental grant from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Brazil (grant number 2327/2015).
Availability of data and materials
The datasets supporting the conclusions of this article are included within the article. The genotyping results with RFLP and microsatellite markers are presented in Additional file 1.
Authors’ contributions
JCRS, FF and HFJP conceived and designed the experiment; JCRS, MFVM, CDFLF and FJRM conducted sample collections; SO, HSS, TFF and JA performed the experiments; RAD, DA and HFJP analysed the data; JCRS, HFJP and DA wrote the manuscript; and LCA, RAM, JPD and SMG critically revised the manuscript. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval
This study was approved by the Ethics Committee of the FMVZ-USP (certificate no. 1921061113), following the National Research Council Guide for the Care and Use of Laboratory Animals.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Additional file
Additional file 1: Table S1.
Toxoplasma gondii genotyping results with RFLP and microsatellite markers from strains from Fernando de Noronha Island and mainland Brazil. (XLSX 30 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Silva, J.C.R., Ferreira, F., Dias, R.A. et al. Cat-rodent Toxoplasma gondii Type II-variant circulation and limited genetic diversity on the Island of Fernando de Noronha, Brazil. Parasites Vectors 10, 220 (2017). https://doi.org/10.1186/s13071-017-2150-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-017-2150-4