
Abstract
Epithelial–mesenchymal transition (EMT) is a critical biological process allowing epithelial cells to de-differentiate into mesenchymal cells. Orchestrated signaling pathways cooperatively induce EMT and effect physiological, sometimes pathological outcomes. Traditional Chinese Medicine (TCM) has been clinically prescribed for thousands of years and recent studies have found that TCM therapies can participate in EMT regulation. In this review, the historical discovery of EMT will be introduced, followed by a brief overview of its major roles in development and diseases. The second section will focus on EMT in organ fibrosis and tissue regeneration. The third section discusses EMT-induced cancer metastasis, and details how EMT contribute to distant dissemination. Finally, new EMT players are described, namely microRNA, epigenetic modifications, and alternative splicing. TCM drugs that affect EMT proven through an evidence-based research approach will be presented in each section.
Similar content being viewed by others
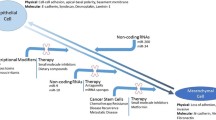


Background
Epithelial–mesenchymal transition (EMT) is the dynamic progression, from well-differentiated epithelial cells into partial or complete fibroblastoid-like mesenchymal cells. Distinct epithelial characteristics are progressively lost, from a specialized apicobasal polarity to a front-rear polarity. Epithelial cell–cell adhesions also dissociate to enable migratory freedom. This reversible EMT process is vital and highly regulated in different stages of embryonic development. Prior to the understanding of EMT, 19th century embryologists had long observed mesenchymal phenotype within epithelial structures of developing tissues. The Matthias Duval’s 1879 hand-drawn atlas of the chicken embryo documented a distinct neural-crest cell population alongside epithelial and mesenchymal cell phenotypes. Ramon y Cajal suggested in 1890 that breast carcinoma cells can delaminate from a cell cluster to invade the adjacent stroma. The concept of EMT was eventually established in the 1960s by Elizabeth Hay through numerous morphological studies conducted on chicken embryos [1,2,3]. The study on EMT in vivo was challenging but Hay and Greenburg inadvertently developed an in vitro model in 1982 to study EMT. Using cells from the adult anterior lens and various embryonic tissues, they observed that epithelial cells undergo major organizational changes when plated on three-dimensional (3D) collagen gels [4]. The environmental conditions generated migratory cells with shape and polarity indistinguishable from mesenchymal cells. This 3D in vitro model allowed further experimentation of EMT processes and the development of different extracellular matrix and diffusible factors [5,6,7,8]. Delving deeper into the underlying mechanisms, Sir Michael Stoker discovered in 1987 that fibroblast-derived scatter factor induces Madin–Darby canine kidney (MDCK) epithelial cell dispersion through a paracrine signaling mechanism [9].
Developmental biologists uncovered multiple signaling pathways controlling EMT during morphogenesis and organogenesis. The fibroblast growth factor (FGF), wingless-type (Wnt), and transforming growth factor-beta (TGF-β) signaling pathways crosstalk at different levels to coordinate development [10,11,12]. The scatter factor discovered by Michael Stoker was eventually identified as the hepatocyte growth factor (HGF) [9, 13, 14]. Together with other growth factors such as FGF-1 and TGFβ [15, 16], normal or transformed epithelial cells were encouraged to disperse. However, extracellular matrix molecules [17] growth factors and extracellular molecules were essential to first stimulate EMT in epithelial cells [18,19,20]. In 1994, the zinc-finger protein SLUG [21] was shown to control EMT in chicken embryo gastrulation. An orthologous master regulatory transcription factor, Snail, was found to mediate mouse gastrulation in a similar fashion. The two related transcription factors, now designated Snai1 (Snail) and Snai2 (Slug), are involved in neural crest ontogeny of different vertebrate embryos [22]. Interestingly, even in invertebrates such as the drosophila, the ortholog of Snai1 designated Snail contributes to cell shape changes in the formation of the ventral furrow during gastrulation [23]. All these findings support the notion that EMT is a universal process that is conserved during evolution.
Unfortunately, the role of EMT extends beyond normal development to disease progression such as tumorigenesis and fibrosis. Discovery of SNAI2-induced EMT in bladder carcinoma cell line [24] reaffirms that EMT promote metastasis in carcinoma through dissolution of desmosomes and remodeling of cytoskeletal proteins. Studies suggest that EMT regulates organ fibrosis [25,26,27] for instance, through inducing cell cycle arrest and parenchymal damage in renal fibrosis [28]. Figure 1a depicts the EMT process.

(permission obtained from original publishers.)
EMT and organ fibrosis. a EMT can be considered as a continued spectrum, where cells present epithelial (E), intermediate (EM), and mesenchymal (M) phenotypes. During EMT, epithelial cells sequentially lose apico-basal polarity and cell–cell adhesions gain front-back polarity and enhanced cell–matrix interactions [27]. b Cell death induces regenerative process, e.g. epithelial cell proliferation. Upregulation of Snai1 and Twist inhibits EC differentiation, leads to myofibroblast and inflammatory cell accumulation and fibrosis [132]
The unwanted implications of EMT in pathogenesis is a motivating factor for scientists to harness viable alternative medicine. Evolved over thousands of years, Traditional Chinese Medicine (TCM) is a medical practice that considers disease as a result of imbalances in the macro-environment. Instead of targeting the local symptoms, treatments are based on a system-wide, multi-target, multi-level holistic approach. Coincidentally, to mitigate the risk of drug resistance, modern medicine is embracing the concept of multi-target therapeutics. The integrative approach in TCM diagnosis and treatment has shown promising effect on EMT in tissue remodeling to cancer progression. With the potential of TCM treatment recognized, majority of cancer patients in China have received TCM in their cancer treatment [29]. Modern drug delivery systems such as targeted drug delivery system (TDDS) also accelerate the applications of TCM on cancer therapy. Passive-targeting TCM preparation, liposomes, emulsion and microspheres offer enormous advantages on traditional drug delivery methods of oral absorption, injection or topical administration [30, 31], allowing for drugs to be concentrated on the target site with higher efficacy and lower side-effects. In this review, we will explore the role of TCM in EMT intervention and discuss novel therapeutic targets for EMT-related organ fibrosis and tumorigenesis.
Tissue remodeling
Organ fibrosis
Organ fibrosis is initiated by a repetitive or persistent injury which in turn trigger inflammatory immune cells to produce cytokines and subsequently recruit and activate quiescent interstitial fibroblasts. Activated contractile fibroblasts, called myofibroblasts, produce excessive extracellular matrix components that deposit as a fibrous network on existing tissue architecture, ultimately augmenting and impairing organ function. Besides arising from chronic injury, myofibroblasts are believed to originate in part from epithelial cells via EMT [32]. In vivo experiments on mouse kidney fibrosis showed that about 30% of fibroblasts are derived from EMT-induced epithelial cells [33]. Studies on fibrotic tissues of the lung, liver, heart and lens also documented EMT as an important source of fibroblasts, as well as perivascular smooth muscle cells and pericytes [34]. Endothelial cells which are progenitor cells of smooth muscle cells and pericytes, are proposed to transform into mesenchymal cells via endothelial–mesenchymal transition (EndMT) [35, 36]. EndMT and EMT are complex biological processes generating mesenchymal-like cells. The cell of origin differs in these two processes, EMT occurs in epithelial cells derived from ectoderm or endoderm. EndEMT occurs in endothelial cells deriving from mesoderm [37]. Nonetheless, EndMT and EMT process activate common transcription factors, such as Snail, Slug, Twist, ZEB1, ZEB2, and Sox2. However, downstream signaling for EndMT results in repressing the expression of endothelial genes (e.g. CDH5 and PECAM1) and subsequently activating the expression of mesenchymal genes (e.g. VIM and COL5A1) [38]. TGF-β1, commonly associated with EMT, is also a major regulator of EndMT in capillary endothelial cells by reducing endothelial markers CD31 and integrin αVβ3 while expressing FSP1, α-SMA, DDR2, collagen I, and vimentin [34]. β-catenin, a central mediator of the Wnt pathway, is an integral member of cell–cell adherens junctions and together with TGF-β1 are common markers used to study the mechanisms in EndMT [39,40,41,42]. Grande et al. studied the mechanisms in detail using a renal fibrosis model and observed that the epithelium does not contribute directly to the increased pool of myofibroblasts [28], Instead, epithelial cells undergo partial EMT, losing epithelial specific markers and its apico-basal polarity while remaining integrated in the tubules [26]. They also produce various cytokines and exosomes that recruit bone marrow mesenchymal progenitors and macrophages exacerbating the fibrotic response [27]. Figure 1b depicts the EMT and organ fibrosis.
A complex molecular and cellular network orchestrates pathological EMT during fibrosis including TGF-β, EGF and FGF2. Using the CCl4-induced liver fibrosis model, the potential of a multi-component traditional herbal formulae, Han-Dan-Gan-Le, as a prescribed treatment was investigated. The rats administered with Han-Dan-Gan-Le compared to the control group displayed 50% lower CCl4-induced hepatic collagen accumulation [43]. Diwu Yanggan (DWYG), which consists of five Chinese herbs, provoked the reversal of EMT to mesenchymal-to-epithelial transition (MET) in the fibrotic liver tissues, with the up-regulation expression of E-cadherin and down-regulation expression of vimentin [44]. The main constituent of DWYG, curcumin, was found in other studies to inhibit TGF-β-Induced EMT via PPARγ pathway in renal tubular epithelial cells in tubulointerstitial fibrosis [45]. Another example is provided by the Wenyang Huazhuo Tongluo formula, which inhibits fibrosis via suppressing Wnt/β-catenin signaling pathway [46]; and combination of Salvia miltiorrhiza and ligustrazine attenuates bleomycin-induced pulmonary fibrosis in rats via modulating TNF-α and TGF-β [47]. The multi-component, multi-level and multi-targeted properties of TCM treatment limit complete understanding of its mechanisms but it holds great clinical potential in the treatment of fibrosis as demonstrated in the examples above. Table 1 provides a list of TCM herbs and prescriptions regulating pathological EMT.
Tissue regeneration
Upon injury, inflammatory signaling modulators recruit diverse types of cells, most predominantly macrophages. These professional phagocytic cells can trigger EMT through the production of growth factors, such as TGF-β, PDGF, EGF, and FGF-2 [48,49,50]. In addition, these cells also secrete chemokines and metalloproteases (MMPs), notably MMP-2, MMP-3, and MMP-9 [51]. During skin wound-healing, keratinocytes undergo a partial EMT process. These keratinocytes, together with surrounding cells induce basement membrane damage while sustaining an intermediate phenotype through the transient expression of SNAI2 [52]. The recruitment of keratinocytes in the wounded area can be inhibited by blocking MMP activity [53]. EGF signaling is also demonstrated to activate EMT in ovarian surface epithelial cells in each menstrual cycle, via MMPs and ILK/ERK kinase [54]. Subsequent phases of wound-healing involve collagen deposition, granulation tissue formation followed by re-epithelialization. These processes may utilize EMT and MET in an orchestrated manner and mediate cell proliferation while restoring the epithelium integrity. Shikonin, a major component of zicao, a TCM herb, has been reported to stimulate EMT in skin wound-healing [55]. Tanshinone IIA [56] another TCM herb, reduces macrophage infiltration and inflammation by inhibiting the TLR4/MyD88/NF-kB pathway. These findings shed new light on developing therapeutic strategies for tissue regeneration.
Cancer progression
Dynamics of junctional complexes in the progression of carcinoma
Carcinoma, which derive from the malignant transformation of epithelial cells, account for 90% of all cancers. Progression of in situ carcinoma to the invasive stage is lethal since carcinoma cells can disseminate and affect distant organs. EMT equip carcinoma with invasive properties capable of disseminating into surrounding stroma and is hypothesized to be the main contributor of metastatic shift in carcinoma.
Epithelial-specific adhesion structures: tight junctions (TJs), adherens junctions (AJs) and desmosomes [57, 58] are dysregulated during EMT. Selectively permeable, TJs allow trans-epithelial transport of ions but forms a physical barrier to fluid movement between the lumen and the stroma. TJs can be destabilized by TGF-β signaling, leading to RhoA degradation and disruption of actin microfilaments [59, 60]. Adherens junctions localized below the tight junctions in polarized epithelial is the first adhesive structure to form following contact between epithelial cells. The junctions initially appear as E-cadherin-containing punctates, which can ultimately assemble into a cortical belt associated with a unique cortical actin microfilament. The E-cadherin cytoplasmic domain controls the dynamics of actin microfilaments assembly and the decrease or disappearance of this prominent EMT marker is a critical step in EMT regulation [61]. The down-regulation of E-cadherin through epigenetic repression is mediated by transcription factors from the Snail, Zeb and Twist families effecting on CpG islands in the promoter region or histone methyltransferases on chromatin organization [62]. E-cadherin can also be down-regulated by post-translational modifications. HGF, EGF and FGF-activating receptor tyrosine kinases (RTK), induce EMT together with the phosphorylation and subsequent ubiquitination and degradation of E-cadherin [63, 64]. Desmosome adhesion is achieved via desmocollins and desmogleins, members of the calcium-dependent cadherin superfamily. The cytoplasmic domain of the desmosomal cadherins binds to plakoglobin, a member of the catenin family, and to desmoplakin which is connected to cytokeratin intermediate filaments [65]. Serine phosphorylation by members of the protein kinase C (PKC) family helps to maintain desmosome stability. In a laboratory setting, monolayer confluency confers desmosome with a calcium-independent property, and thus, stability even from divalent cation disruption [66].
Well-developed TJs are absent in the early intermediate stage of EMT while AJ destabilization, usually accompanied by desmosome disassembly, is the hallmark of the intermediate late stage of EMT. Initially considered as an epithelial to mesenchymal transformation process, many EMT-induced carcinomas remain in intermediate states during EMT. Intermediaries oscillate in their phenotypes and possess relative ability to reassemble immature AJ. Expression of major AJ constituent, E-cadherin, is repressed by ZEB1, a transcription factor regulated by miR200 family members [67]. The layered feedback loops form a regulatory cascade that affect E-cadherin turnover and subsequently, EMT progression [29]. To reach complete mesenchymal state will require tight control of adhesive components by transcriptional repressors and epigenetic marks [62, 68].
The effect of TCM herbs on EMT is extensive and not well-elucidated but some have been studied in greater detail, especially its effect on EMT-associated adhesion complex. Ginsenoside Rg3, a compound abundant in Ginseng panax, could significantly alter EMT markers by increasing E-cadherin and decreasing expression of SNAI1, N-cadherin and vimentin [69]. Resveratrol, a natural polyphenolic compound found in Polygonum cuspidatum, grapes and peanuts, can inhibit LPS-induced EMT morphological changes in PC-3 and LNCaP PCa prostate cancer cells [70]. LPS-exposure upregulated transcription factor glioma-associated oncogene homolog 1 (Gli1) but resveratrol inhibited this gene. In another study, resveratrol reduced human lung carcinoma cell line A549 spheroid dispersion by 40% compared to the control sample [71]. Triptolide, a compound extracted from the TCM Tripterygium wilfordii, inhibited metastasis of ovarian cancer cells SKOV3 and A2780 via MMP7 and MMP19 repression and E-cadherin activation [51]. The in vivo mouse model conducted in the same study supported the observation that triptolide can inhibit tumor formation and metastasis in nude mice. Thymoquinone, extracted from Nigella sativa, inhibit EMT in cervical cancer cell lines SiHa and CaSki by inactivating EMT signaling, TWIST1 and ZEB1 repression, and preventing cellular migration through E-cadherin activation [72].
EMT signaling in the context of the tumor microenvironment
The tumor microenvironment is considered crucial in the initiation and progression of malignant cells [73]. Recent findings support the notion that cancer cells require support from stromal cells before it can expand autonomously. An EMT fate is dependent on the reciprocal interactions between carcinoma cells and the stroma with the interplay of growth factors, cytokines and chemokines in the microenvironment (Fig. 2). HGF, EGF, FGF, IGF, PDGF, TGF-β, Wnt family members, Sonic Hedgehog, Notch, interleukins and chemokines are signaling molecules that have been shown to induce EMT [74, 75]. EMT signaling molecules can activate diverse pathways such as Ras, Smad and β-catenin. Downstream transcription factors such as SNAI1, SNAI2, zinc finger E-box binding homeobox 1 and 2 (ZEB1 and ZEB2), TWIST1, Forkhead boxC2 (FOXC2), high mobility group A2 (HMGA2) and paired related homeobox 1 (PRX1) are subsequently activated in carcinoma [62, 76,77,78,79,80,81].

Snapshot of EMT signaling pathways. EMT in carcinoma is induced by various pathways, either by growth factors and cytokines from adjacent cells, or via cell interactions with extracellular matrix (ECM). In the paracrine mode, Receptor Tyrosine kinases (RTKs), TGFβ, Wnt and Notch can be activated. EMT could also be induced through receptors such as integrins interacting with ECM proteins like collagens, subsequently activating intracellular kinases such as Src. Signal transduction involves transcription factors and polarity genes [11, 61, 82, 88]
TGF-β is one of the key element that promotes invasion and metastasis by altering apico-basolateral polarity, generating tumor initiating cells, and sheltering the tumor from immune surveillance and apoptosis signals [82,83,84]. Initiated by ligand binding to type II TGF-β receptors, the canonical TGF-β signaling leads to phosphorylation of regulatory SMAD2 and SMAD3. The non-canonical TGF-β pathway involves several non-receptor protein tyrosine kinases (SRC/FAK), p38 MAPK and the GTPase RhoA [85,86,87]. Curcumin inhibits TGF-β1-induced EMT in hepatoma cells by down-regulating downstream SMAD2 phosphorylation. Assembly of phosphorylated SMAD2:SMAD3 with SMAD4 into a heterotrimeric complex [88] and translocation into the nucleus to control gene transcription is prevented, effectively inhibiting EMT [89]. Jianpi Huayu Decoction (JHD), a TCM prescription against cancer, is found to halt EMT hepatocellular carcinoma by regulating the Smad3/Smad7 cascade. Smad7 expression is increased while expression of p-Smad3 and Snail decreased leading to E-cadherin up-regulation and EMT inhibition [90].
EMT and cancer niche
An inflammatory site is often established prior to the appearance of in situ cancer cells and is termed the cancer niche [91]. The normal epithelium cells in this niche are more susceptible to stress triggers, such as chemical carcinogenesis, to undergo mutation and malignancy changes. The recruitment of bone marrow immature myeloid cells usually accompanies the onset of inflammatory stroma expansion adjacent to the dysplastic lesion. Different stromal cell types, fibroblasts, endothelial cells and pericytes, have been identified in tumours associated with angiogenesis, myeloid and lymphoid lineages [71, 92]. The stroma cells interact closely with carcinoma cells to produce many growth factors and cytokines that contribute to the acquisition of a mesenchymal-like phenotype. Although the regulation of adhesion mechanisms is crucial in the control of EMT status in carcinoma, stroma cells operate in a tissue-specific manner that affect EMT outcome. A detailed study was performed to characterize mesenchymal stems cells under ex vivo autocrine and paracrine loops from colon carcinoma cells [93]. The study found that the colon carcinoma line secreted IL-1 which induced prostaglandinE2 (PEG2) production through the cyclooxygenase2 (COX2) and microsomal prostaglandinE2 synthase-1 increased level. Secreted PEG2 promotes the autocrine production of IL-6, IL-8 and Gro-α. In addition, PEG2 induces nuclear translocation of β-catenin through inactivation of GSK3β by AKT activation. The crosstalk between colon carcinoma cells and mesenchymal stem cell led to the emergence of ALDH-1/CD133 double positive tumor initiating cells.
Shaoyao decoction (SYD), a TCM prescription that originated in the Jin-Yuan Dynasty, was found to suppress colitis-associated colorectal cancer. The inflammatory niche in colitis is addressed as SYD repressed proinflammatory cytokines IL-1β, IL-6, and TNF-α. Thus, inhibiting Snail-induced EMT by attenuating TAM infiltration and NF-κB activation [94].
EMT may prevent senescence induced by oncogenes, therefore inducing the dissemination of carcinoma [95, 96].
A ‘precancer niche’ is a prerequisite for the survival and development of EMT-initiated carcinoma cells [91]. Similarly, deriving from murine experimental models of metastasis, creation of a premetastatic niche at presumptive sites of metastasis is critical. It starts with a premetastatic inflammatory environment that will be seeded by micrometastatic circulating tumor cells [97].
These circulating tumor cells (CTC) colonize and recruit endothelial progenitor cells (EPCs) that activate the angiogenic switch and shift micro-metastasis towards macro-metastasis [98]. Figure 3 describes the metastatic cascade in relation to the tumor stroma. EPCs are a subset of bone marrow-derived cells (BMDCs) that possess EndMT capabilities. BMDCs include macrophages, myeloid-derived suppressor cells (MDSCs), TIE2-expressing monocytes (TEMs), mesenchymal stem cells (MSCs), and platelets [34, 99]. Recent findings suggest exosomes from tumors regulate the immune response and hematopoietic system. These exosomes contain proteins and nucleic acids that induce lineage-specific differentiation of bone marrow precursors [91, 100, 101], and recruit inflammatory cells (such as MDSC) to distant sites [102].

EMT in the metastatic cascade. Distinct stromal cells promote the dissemination of carcinoma cells, such as myofibroblasts, natural killer cells, T or B lymphocytes and macrophages. The scheme outlines the transition from an epithelial-like phenotype to a mesenchymal-like phenotype during local invasion and intravasation. Carcinoma cells can re-acquire through MET an epithelial-like phenotype following extravasation colonization of the secondary sites and proliferation. Macrophages can initiate a feedback-loop of EGF and Colony Stimulating Factor 1 (CSF1) to promote EMT. Myofibroblasts produce and reprogram ECM, secrete pro-inflammatory factors, and induce epithelial cell proliferation and invasion. BMDC contribute to the genesis of the premetastatic niche. Lymphocyte trafficking is also an important metastasis regulator. MDSCs interact with innate/adaptive immune cells, and promote cancer escape and angiogenesis in the cancer niche. MDSC and NK cells are also critical for MET process by populate premetastatic niches, where they regulate metastatic dissemination, initiate and promote tumor colonization
Individual cell types within the tumor niche cells play distinct roles on tumor niche formation and cancer metastasis. Tumor-associated macrophages (TAMs) have a range of stromal cell types with significantly different physiological functions represented by polarized states M1, M2a, M2b and M2c [103]. It is hypothesized that TAMs exist either as a mixture of different polarized states or exhibit intermediary stages possessing partial functions of each polarized state [104, 105]. Macrophages can initiate a feedback loop of EGF and Colony Stimulating Factor 1 (CSF1) to promote EMT [106]. Most macrophages found in evolving tumors adopt the M2 polarized state, an immunosuppressive state-of-cell that is cancer promoting. Interestingly, different cell types within each family of macrophages and cytokines act upon specific cancers and sometimes, different regions of the same tumor. Proinflammatory M1-like TAMs are uniquely found in colorectal cancers and unlike M2 macrophages, suppress tumor progression by mediating cell killing [107]. Qingyihuaji Formula (QYHJ), a seven-herb patented TCM prescription, has significant anti-metastatic effects on pancreatic cancer even in gemcitabine resistant ones. QYHJ inhibits cancer-related inflammation in tumors by decreasing infiltration of tumor-associated macrophages and production of IL-6 [67].
MDSC is another cancer niche cell type and share the same progenitors as TAMs. They suppress immune responses to newly displayed tumor antigens. Thus, an increase in the heterogeneous collection of immature myeloid cells is observed in the bone marrow and blood of cancer patients [108]. MDSCs promote EMT possibly through TGF-β signaling, or via S100 series proteins [109]. Sheng Qi Formula (SQF) is commonly used to minimize chemotherapy side effects and as a complimentary cancer treatment. In vivo mouse model studies found that SQF formula significantly inhibited 4T1 mammary tumor development [81]. SQF treatment inhibited hydrogen peroxide production by Gr1+CD11b+ cells which in turn, reduced MDSCs protective effect on cancer cells. MDSCs interact with adaptive immune cells, to promote cancer escape and angiogenesis in the cancer niche [91, 110]. Feiyanning Decoction (FYN) is an antitumor prescription that restores T cell toxicity. In the Lewis lung carcinoma model, FYN was found to lower the ratio of CD4+CD25+ regulatory T cells (TREG) and down-regulate Foxp3 oncogene expression in TREG [80].
Cancer EMT and chronic inflammation
The presence of chronic inflammation is strongly associated with EMT and the dynamic recruitment of BMDCs [50]. Inflammatory cytokine tumor necrosis factor (TNF-α), through NF-κB signaling, stabilizes SNAI1 for inflammation-induced metastasis [111]. Upon NF-κB p65 and IKK-β activation, EMT transcription factor TWIST1 expression is enhanced [112]. NF-κB signaling forms a positive feedback-loop which maintain high levels of pro-inflammatory cytokine (IL-1, TNF-α, IL-6), chemokine (IL-8, MIP-1, MCP-1), adhesion molecule (e.g. ICAM, VCAM) and growth factors [113]. As a result, inflammatory response cells like TAMs, T-cells and fibroblasts are recruited and accumulation of hypoxia inducible factor 1 (HIF-1) is favored [114]. Hypoxic responses induced by HIF-1 is also associated with activation of Twist1 and Snail1 [115]. Yi Ai Fang (YAF), a TCM formula, has been found to be effective against vasculogenic mimicry (VM) in colorectal cancer via HIF-1α/EMT regulation [116]. Another example is matrine, a natural alkaloid isolated from the traditional herb medicine sophora flavescens. This compound inhibited EMT in pancreatic cancer cells via ROS/NF-κB/MMPs pathway regulation [117].
New “players” in EMT
EMT is a process which involves the dynamic cooperation of different hierarchical pathways involving multiple regulatory layers. Firstly, it is governed by transcriptional/translational changes in an orchestrated manner as discussed in previous paragraphs. Secondly, it is also regulated epigenetically and post-translationally. The activity of EMT transcription factors of the Snail, Zeb and Twist families is dependent on different epigenetic modifiers [79]. Epigenetic control of gene expression via DNA methylation on CpG islands is a conventional mechanism for silencing tumor suppression genes in tumor cells [118]. Epigenetic modification of histones is also involved in EMT where SNAI1 induces repressive histone modifications through histone deacetylase (HDAC1) and HDAC2 [119]. Several other histone modifiers have been associated to SNAI1 functions, such as demethylase LSD1, methyltransferase EZH2 and SUZ12 [62, 79, 120, 121].
MicroRNAs are also important in regulation of EMT inducers [122]. The family of miR-200 and the Zeb family of transcription factors control EMT and MET through negative feedback loops [123, 124]. SNAI1-dependent EMT is directly regulated by miR-34 family [125, 126]. Ginsenoside Rb1, extracted from the same plant as Ginsenoside Rb3, lowered miR-25 levels and inhibited cell migration abilities in SKOV3 and 3AO human ovarian cancer cells. The treatment reversed hypoxia-induced EMT by effecting on EP300 and E‑cadherin [66]. Genistein, a flavonoid found in a TCM herb Sophora japonica, promoted apoptosis while inhibiting prostate cancer cell proliferation, invasion and Wnt signaling in prostate cancer. MiR-1260b is highly expressed in prostate cancer tissues but genistein treatment significantly reduced it. sFRP1 and SMAD4 expression that are decreased in prostate cancer tissues can be partially restored by genistein via DNA demethylation and histone modifications [127].
Alternative splicing was recently reported to control EMT. Two RNA binding proteins, epithelial splicing regulatory protein 1 and 2 were recognized as critical promoters of FGFR2 splicing. The FGFR2b isoform is exclusively expressed in epithelial cell while the FGFR2c in mesenchymal cells [79, 128]. Jin-Fu-An decoction decreased the expression of p120ctn, isoform 1A, and its S288 phosphorylation, which reduced metastasis of H1650 lung cancer cell [129].
Conclusions
In this review, the physiological and pathological EMT driven processes are summarized and the regulatory roles of TCM herbs and prescriptions on these processes are studied. The complexity of the EMT process demands efficient therapeutic interventions that have multiple layers of regulation. This criterion can be met by newly discovered or ancient TCM-derived therapies backed with modern evidence-based studies. Scientists found the treatments useful in reducing local invasion, distant dissemination, chemotherapy resistance, and cancer immune escape. Deep sequencing of cancer genomes has shown that stable driver mutations are difficult to identify in most solid tumors as they contain thousands of mutations on average and constantly evolve during tumor progression. Thus, multi-targeted therapies, such as TCM-derived therapies, regulating the unstable global status of cancer cells is significant in improving treatment response [18, 130, 131]. Thus, TCM is one strategy which can bring up new opportunities of generic treatment with the concept of evidence-based therapy.
Another challenge in cancer research remains in global mapping of EMT epigenetic modifications and related therapeutic opportunities. Technical developments in omics should provide better profiling analysis of translational interactions within cells in the EMT spectrum. TCM drugs have shown efficacy in targeting epigenetic modifications associated with EMT. However, the underlying mechanisms involved in its effect will require greater investigation. Current challenges remain such as isolating and identifying individual effective components from TCM, searching for lead compounds in herbal medicine, minimizing toxicity and establishing standard guidelines for clinical trials. Nonetheless TCM-derived therapies still hold great potential and the significant increase in studies to deconvolute heterogeneous herbal mixtures will aid modern evidence-based study of TCM on EMT and cancer research.
Availability of data and materials
All data included in this article are available from the corresponding author upon request.
Abbreviations
- AJ:
-
adherens junction
- BMDCs:
-
bone marrow-derived cells
- CTC:
-
circulating tumor cells
- CSF1:
-
colony stimulating factor 1
- COX2:
-
cyclooxygenase2
- EPCs:
-
endothelial progenitor cells
- EGF:
-
epidermal growth factor
- EMT:
-
epithelial–mesenchymal transition
- FGF:
-
fibroblast growth factor
- FOXC2 :
-
Forkhead boxC2
- Gli1:
-
glioma-associated oncogene homolog 1
- HGF:
-
hepatocyte growth factor
- HMGA2 :
-
high mobility group A2
- HIF-1:
-
hypoxia inducible factor 1
- IGF:
-
insulin-like growth factor
- LPS:
-
lipopolysaccharides
- MDCK:
-
Madin–Darby canine kidney
- MSCs:
-
mesenchymal stem cells
- MET:
-
mesenchymal-to-epithelial transition
- MMP:
-
metalloprotease
- MDSCs:
-
myeloid-derived suppressor cells
- PRX1 :
-
paired related homeobox 1
- PDGF:
-
platelet-derived growth factor
- PEG2:
-
prostaglandinE2
- PKC:
-
protein kinase C
- RTK:
-
receptor tyrosine kinases
- TREG:
-
regulatory T cells
- 3D:
-
three-dimensional
- TEMs:
-
TIE2-expressing monocytes
- TJ:
-
tight junction
- TCM:
-
Traditional Chinese Medicine
- TGF-β:
-
transforming growth factor-beta
- TNF-α:
-
tumor necrosis factor
- TAMs:
-
tumor-associated macrophages
- VM:
-
vasculogenic mimicry
- Wnt:
-
wingless-type
- ZEB1 and ZEB2 :
-
zinc finger E-box binding homeobox 1 and 2
References
Trelstad RL, Hay ED, Revel JD. Cell contact during early morphogenesis in the chick embryo. Dev Biol. 1967;16(1):78–106.
Trelstad RL, Revel JP, Hay ED. Tight junctions between cells in the early chick embryo as visualized with the electron microscopy. J Cell Biol. 1966;31(1):C6–10.
Hay ED. Organization and fine structure of epithelium and mesenchyme in the developing chick embryo: In: Epithelial–mesenchymal interactions;. 18th Hahnemann Symposium; 1968.
Hay GGE. Epithelia suspended in collagen gels can lose polarity and express characteristics of migrating mesenchymal cells. J Cell Biol. 1982;95:333–9.
Wessells NK. Tissue interactions and development. Menlo Park: W. A. Benjamin; 1977.
Hay ED. Cell biology of extracellular matrix. New York: Springer; 1991.
Birk DE, Trelstad RL. Extracellular compartments in matrix morphogenesis: collagen fibril, bundle, and lamellar formation by corneal fibroblasts. J Cell Biol. 1984;99(6):2024–33.
Gurdon JB. Embryonic induction–molecular prospects. Development (Cambridge, England). 1987;99(3):285–306.
Stoker M, Gherardi E. Factors affecting epithelial interactions. Ciba Found Symp. 1987;125:217–39.
Ciruna B, Rossant J. FGF signaling regulates mesoderm cell fate specification and morphogenetic movement at the primitive streak. Dev Cell. 2001;1(1):37–49.
Lim J, Thiery JP. Epithelial–mesenchymal transitions: insights from development. Development (Cambridge, England). 2012;139(19):3471–86.
Mayor R, Theveneau E. The neural crest. Development (Cambridge, England). 2013;140(11):2247–51.
Stoker M, Gherardi E, Perryman M, Gray J. Scatter factor is a fibroblast-derived modulator of epithelial cell mobility. Nature. 1987;327(6119):239–42.
Naldini L, Weidner KM, Vigna E, Gaudino G, Bardelli A, Ponzetto C, Narsimhan RP, Hartmann G, Zarnegar R, Michalopoulos GK, et al. Scatter factor and hepatocyte growth factor are indistinguishable ligands for the MET receptor. EMBO J. 1991;10(10):2867–78.
Valles AM, Boyer B, Badet J, Tucker GC, Barritault D, Thiery JP. Acidic fibroblast growth factor is a modulator of epithelial plasticity in a rat bladder carcinoma cell line. Proc Natl Acad Sci USA. 1990;87(3):1124–8.
Moustakas A, Heldin CH. The regulation of TGFbeta signal transduction. Development (Cambridge, England). 2009;136(22):3699–714.
Kelley LC, Lohmer LL, Hagedorn EJ, Sherwood DR. Traversing the basement membrane in vivo: a diversity of strategies. J Cell Biol. 2014;204(3):291–302.
Pastushenko I, Brisebarre A, Sifrim A, Fioramonti M, Revenco T, Boumahdi S, Van Keymeulen A, Brown D, Moers V, Lemaire S, et al. Identification of the tumour transition states occurring during EMT. Nature. 2018;556(7702):463–8.
Liu J, Ke F, Cheng H, Zhou J. Traditional Chinese medicine as targeted treatment for epithelial–mesenchymal transition-induced cancer progression. J Cell Biochem. 2018;120:1068–79.
Thiery JP. Epithelial–mesenchymal transitions in tumour progression. Nat Rev Cancer. 2002;2(6):442–54.
Nieto MA, Sargent MG, Wilkinson DG, Cooke J. Control of cell behavior during vertebrate development by Slug, a zinc finger gene. Science (New York, NY). 1994;264(5160):835–9.
Barrallo-Gimeno A, Nieto MA. The Snail genes as inducers of cell movement and survival: implications in development and cancer. Development (Cambridge, England). 2005;132(14):3151–61.
Leptin M, Roth S. Autonomy and non-autonomy in Drosophila mesoderm determination and morphogenesis. Development (Cambridge, England). 1994;120(4):853–9.
Savagner P, Yamada KM, Thiery JP. The zinc-finger protein slug causes desmosome dissociation, an initial and necessary step for growth factor-induced epithelial–mesenchymal transition. J Cell Biol. 1997;137(6):1403–19.
Zeisberg M, Neilson EG. Mechanisms of tubulointerstitial fibrosis. J Am Soc Nephrol JASN. 2010;21(11):1819–34.
Grande MT, Sanchez-Laorden B, Lopez-Blau C, De Frutos CA, Boutet A, Arevalo M, Rowe RG, Weiss SJ, Lopez-Novoa JM, Nieto MA. Snail1-induced partial epithelial-to-mesenchymal transition drives renal fibrosis in mice and can be targeted to reverse established disease. Nat Med. 2015;21(9):989–97.
Nieto MA, Huang RY, Jackson RA, Thiery JP. Emt: 2016. Cell. 2016;166(1):21–45.
Lovisa S, LeBleu VS, Tampe B, Sugimoto H, Vadnagara K, Carstens JL, Wu CC, Hagos Y, Burckhardt BC, Pentcheva-Hoang T, et al. Epithelial-to-mesenchymal transition induces cell cycle arrest and parenchymal damage in renal fibrosis. Nat Med. 2015;21(9):998–1009.
Chen Z, Gu K, Zheng Y, Zheng W, Lu W, Shu XO. The use of complementary and alternative medicine among Chinese women with breast cancer. J Altern Complement Med. 2008;14(8):1049–55.
Xu W, Xing FJ, Dong K, You C, Yan Y, Zhang L, Zhao G, Chen Y, Wang K. Application of traditional Chinese medicine preparation in targeting drug delivery system. Drug Deliv. 2015;22(3):258–65.
Li L, Feng Y, Hong Y, Lin X, Shen L. Recent advances in drug delivery system for bioactive glycosides from Traditional Chinese Medicine. Am J Chin Med. 2018;46(8):1791–824.
Iwano M, Plieth D, Danoff TM, Xue C, Okada H, Neilson EG. Evidence that fibroblasts derive from epithelium during tissue fibrosis. J Clin Investig. 2002;110(3):341–50.
Zavadil J, Bottinger EP. TGF-beta and epithelial-to-mesenchymal transitions. Oncogene. 2005;24(37):5764–74.
Kalluri R, Weinberg RA. The basics of epithelial–mesenchymal transition. J Clin Investig. 2009;119(6):1420–8.
Zeisberg EM, Tarnavski O, Zeisberg M, Dorfman AL, McMullen JR, Gustafsson E, Chandraker A, Yuan X, Pu WT, Roberts AB, et al. Endothelial-to-mesenchymal transition contributes to cardiac fibrosis. Nat Med. 2007;13(8):952–61.
Potenta S, Zeisberg E, Kalluri R. The role of endothelial-to-mesenchymal transition in cancer progression. Br J Cancer. 2008;99(9):1375–9.
Saito A. EMT and EndMT: regulated in similar ways? J Biochem. 2013;153(6):493–5.
Man S, Sanchez Duffhues G, Ten Dijke P, Baker D. The therapeutic potential of targeting the endothelial-to-mesenchymal transition. Angiogenesis. 2019;22(1):3–13.
Galichon P, Hertig A. Epithelial to mesenchymal transition as a biomarker in renal fibrosis: are we ready for the bedside? Fibrogenesis Tissue Repair. 2011;4:11.
Kumarswamy R, Volkmann I, Jazbutyte V, Dangwal S, Park DH, Thum T. Transforming growth factor-beta-induced endothelial-to-mesenchymal transition is partly mediated by microRNA-21. Arterioscler Thromb Vasc Biol. 2012;32(2):361–9.
Strutz F, Okada H, Lo CW, Danoff T, Carone RL, Tomaszewski JE, Neilson EG. Identification and characterization of a fibroblast marker: FSP1. J Cell Biol. 1995;130(2):393–405.
Hosper NA, van den Berg PP, de Rond S, Popa ER, Wilmer MJ, Masereeuw R, Bank RA. Epithelial-to-mesenchymal transition in fibrosis: collagen type I expression is highly upregulated after EMT, but does not contribute to collagen deposition. Exp Cell Res. 2013;319(19):3000–9.
Wang JXY, Hu XX, Wang Y. Handan Ganle inhibits PI3K/Akt signaling pathway in liver fibrosis in rats. World Chin J Digestol. 2014;22(14):1915–20.
Shen X, Cheng S, Peng Y, Song H, Li H. Attenuation of early liver fibrosis by herbal compound “Diwu Yanggan” through modulating the balance between epithelial-to-mesenchymal transition and mesenchymal-to-epithelial transition. BMC Complementary Altern Med. 2014;14:418.
Li R, Wang Y, Liu Y, Chen Q, Fu W, Wang H, Cai H, Peng W, Zhang X. Curcumin inhibits transforming growth factor-beta1-induced EMT via PPARgamma pathway, not Smad pathway in renal tubular epithelial cells. PLoS ONE. 2013;8(3):e58848.
Wang Q, Zang W, Han L, Yang L, Ye S, Ouyang J, Zhang C, Bi Y, Zhang C, Bian H. Wenyang Huazhuo Tongluo formula inhibits fibrosis via suppressing Wnt/beta-catenin signaling pathway in a Bleomycin-induced systemic sclerosis mouse model. Chin Med. 2018;13:17.
Huang C, Wu X, Wang S, Wang W, Guo F, Chen Y, Pan B, Zhang M, Fan X. Combination of Salvia miltiorrhiza and ligustrazine attenuates bleomycin-induced pulmonary fibrosis in rats via modulating TNF-alpha and TGF-beta. Chin Med. 2018;13:36.
Strutz F, Zeisberg M, Ziyadeh FN, Yang CQ, Kalluri R, Muller GA, Neilson EG. Role of basic fibroblast growth factor-2 in epithelial–mesenchymal transformation. Kidney Int. 2002;61(5):1714–28.
Duffield JS, Lupher M, Thannickal VJ, Wynn TA. Host responses in tissue repair and fibrosis. Annu Rev Pathol. 2013;8:241–76.
Lopez-Novoa JM, Nieto MA. Inflammation and EMT: an alliance towards organ fibrosis and cancer progression. EMBO Mol Med. 2009;1(6–7):303–14.
Zhao H, Yang Z, Wang X, Zhang X, Wang M, Wang Y, Mei Q, Wang Z. Triptolide inhibits ovarian cancer cell invasion by repression of matrix metalloproteinase 7 and 19 and upregulation of E-cadherin. Exp Mol Med. 2012;44(11):633–41.
Arnoux V, Nassour M, L’Helgoualc’h A, Hipskind RA, Savagner P. Erk5 controls Slug expression and keratinocyte activation during wound healing. Mol Biol Cell. 2008;19(11):4738–49.
Yang J, Shultz RW, Mars WM, Wegner RE, Li Y, Dai C, Nejak K, Liu Y. Disruption of tissue-type plasminogen activator gene in mice reduces renal interstitial fibrosis in obstructive nephropathy. J Clin Investig. 2002;110(10):1525–38.
Ahmed N, Maines-Bandiera S, Quinn MA, Unger WG, Dedhar S, Auersperg N. Molecular pathways regulating EGF-induced epithelio-mesenchymal transition in human ovarian surface epithelium. Am J Physiol Cell Physiol. 2006;290(6):C1532–42.
Yin SY, Peng AP, Huang LT, Wang YT, Lan CW, Yang NS. The phytochemical shikonin stimulates epithelial–mesenchymal transition (EMT) in skin wound healing. Evid Based Complement Alternat Med. 2013;2013:262796.
Wu DM, Wang YJ, Han XR, Wen X, Li L, Xu L, Lu J, Zheng YL. Tanshinone IIA prevents left ventricular remodelling via the TLR4/MyD88/NF-kappaB signalling pathway in rats with myocardial infarction. J Cell Mol Med. 2018;22(6):3058–72.
Medigeshi GR, Hirsch AJ, Brien JD, Uhrlaub JL, Mason PW, Wiley C, Nikolich-Zugich J, Nelson JA. West nile virus capsid degradation of claudin proteins disrupts epithelial barrier function. J Virol. 2009;83(12):6125–34.
Huang RY, Guilford P, Thiery JP. Early events in cell adhesion and polarity during epithelial–mesenchymal transition. J Cell Sci. 2012;125(Pt 19):4417–22.
Barrios-Rodiles M, Brown KR, Ozdamar B, Bose R, Liu Z, Donovan RS, Shinjo F, Liu Y, Dembowy J, Taylor IW, et al. High-throughput mapping of a dynamic signaling network in mammalian cells. Science (New York, NY). 2005;307(5715):1621–5.
Ozdamar B, Bose R, Barrios-Rodiles M, Wang HR, Zhang Y, Wrana JL. Regulation of the polarity protein Par6 by TGFbeta receptors controls epithelial cell plasticity. Science (New York, NY). 2005;307(5715):1603–9.
Thiery JP, Sleeman JP. Complex networks orchestrate epithelial–mesenchymal transitions. Nat Rev Mol Cell Biol. 2006;7(2):131–42.
Tam WL, Weinberg RA. The epigenetics of epithelial–mesenchymal plasticity in cancer. Nat Med. 2013;19(11):1438–49.
Fujita Y, Hogan C, Braga VM. Regulation of cell-cell adhesion by Rap1. Methods Enzymol. 2006;407:359–72.
Mukherjee N, Bhattacharya N, Alam N, Roy A, Roychoudhury S, Panda CK. Subtype-specific alterations of the Wnt signaling pathway in breast cancer: clinical and prognostic significance. Cancer Sci. 2012;103(2):210–20.
Thomason HA, Scothern A, McHarg S, Garrod DR. Desmosomes: adhesive strength and signalling in health and disease. Biochem J. 2010;429(3):419–33.
Liu D, Liu T, Teng Y, Chen W, Zhao L, Li X. Ginsenoside Rb1 inhibits hypoxia-induced epithelial–mesenchymal transition in ovarian cancer cells by regulating microRNA-25. Exp Ther Med. 2017;14(4):2895–902.
Zhang J, Wang P, Ouyang H, Yin J, Liu A, Ma C, Liu L. Targeting cancer-related inflammation: Chinese herbal medicine inhibits epithelial-to-mesenchymal transition in pancreatic cancer. PLoS ONE. 2013;8(7):e70334.
Moreno-Bueno G, Portillo F, Cano A. Transcriptional regulation of cell polarity in EMT and cancer. Oncogene. 2008;27(55):6958–69.
Tian L, Shen D, Li X, Shan X, Wang X, Yan Q, Liu J. Ginsenoside Rg3 inhibits epithelial–mesenchymal transition (EMT) and invasion of lung cancer by down-regulating FUT4. Oncotarget. 2016;7(2):1619–32.
Li J, Chong T, Wang Z, Chen H, Li H, Cao J, Zhang P, Li H. A novel anticancer effect of resveratrol: reversal of epithelialmesenchymal transition in prostate cancer cells. Mol Med Rep. 2014;10(4):1717–24.
Niu Y, Bai J, Kamm RD, Wang Y, Wang C. Validating antimetastatic effects of natural products in an engineered microfluidic platform mimicking tumor microenvironment. Mol Pharm. 2014;11(7):2022–9.
Li J, Khan MA, Wei C, Cheng J, Chen H, Yang L, Ijaz I, Fu J. Thymoquinone inhibits the migration and invasive characteristics of cervical cancer cells SiHa and CaSki in vitro by targeting epithelial to mesenchymal transition associated transcription factors Twist1 and Zeb1. Molecules. 2017;22(12):2105.
Jain RK. Normalizing tumor microenvironment to treat cancer: bench to bedside to biomarkers. J Clin Oncol. 2013;31(17):2205–18.
Bai J, Tu TY, Kim C, Thiery JP, Kamm RD. Identification of drugs as single agents or in combination to prevent carcinoma dissemination in a microfluidic 3D environment. Oncotarget. 2015;6(34):36603–14.
Aref AR, Huang RY, Yu W, Chua KN, Sun W, Tu TY, Bai J, Sim WJ, Zervantonakis IK, Thiery JP, et al. Screening therapeutic EMT blocking agents in a three-dimensional microenvironment. Integr Biol. 2013;5(2):381–9.
Jechlinger M, Grunert S, Beug H. Mechanisms in epithelial plasticity and metastasis: insights from 3D cultures and expression profiling. J Mammary Gland Biol Neoplasia. 2002;7(4):415–32.
Shi Y, Massague J. Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell. 2003;113(6):685–700.
Medici D, Hay ED, Olsen BR. Snail and Slug promote epithelial–mesenchymal transition through beta-catenin-T-cell factor-4-dependent expression of transforming growth factor-beta3. Mol Biol Cell. 2008;19(11):4875–87.
De Craene B, Berx G. Regulatory networks defining EMT during cancer initiation and progression. Nat Rev Cancer. 2013;13(2):97–110.
Guo J, Wang JY, Zheng Z, Wang Q, Dong CS. Effects of Chinese herbal medicine Feiyanning decoction on the ratio of CD4+ CD25+ regulatory T cells and expression of transcription factor Foxp3 in mice bearing Lewis lung carcinoma. Zhong xi yi jie he xue bao J Chin Integr Med. 2012;10(5):584–90.
Li J, Chen X, Lin HS, Oppenheim JJ, Howard OZ. Inhibitory effect of Sheng Qi Formula (SQF) on Gr-1+ CD11b+ myeloid immunosuppressor cells (MIC) in the 4T1 murine mammary cancer model. In: AACR annual meeting. San Diego: American Association for Cancer Research; 2008.
Heldin CH, Vanlandewijck M, Moustakas A. Regulation of EMT by TGF beta in cancer. FEBS Lett. 2012;586(14):1959–70.
Massague J. TGFbeta in cancer. Cell. 2008;134(2):215–30.
Taylor MA, Parvani JG, Schiemann WP. The pathophysiology of epithelial–mesenchymal transition induced by transforming growth factor-beta in normal and malignant mammary epithelial cells. J Mammary Gland Biol Neoplasia. 2010;15(2):169–90.
Bhowmick NA, Ghiassi M, Bakin A, Aakre M, Lundquist CA, Engel ME, Arteaga CL, Moses HL. Transforming growth factor-beta1 mediates epithelial to mesenchymal transdifferentiation through a RhoA-dependent mechanism. Mol Biol Cell. 2001;12(1):27–36.
Bhowmick NA, Zent R, Ghiassi M, McDonnell M, Moses HL. Integrin beta 1 signaling is necessary for transforming growth factor-beta activation of p38MAPK and epithelial plasticity. J Biol Chem. 2001;276(50):46707–13.
Parvani JG, Taylor MA, Schiemann WP. Noncanonical TGF-beta signaling during mammary tumorigenesis. J Mammary Gland Biol Neoplasia. 2011;16(2):127–46.
Morrison CD, Parvani JG, Schiemann WP. The relevance of the TGF-beta Paradox to EMT-MET programs. Cancer Lett. 2013;341(1):30–40.
Cao MT, Liu HF, Liu ZG, Xiao P, Chen JJ, Tan Y, Jiang XX, Jiang ZC, Qiu Y, Huang HJ, et al. Curcumin downregulates the expression of Snail via suppressing Smad2 pathway to inhibit TGF-beta1-induced epithelial–mesenchymal transitions in hepatoma cells. Oncotarget. 2017;8(65):108498–508.
Zhong C, Zhang YF, Huang JH, Wang ZY, Chen QY, Su LT, Liu ZT, Xiong CM, Tao Z, Guo RP. The Chinese medicine, Jianpi Huayu Decoction, inhibits the epithelial mesenchymal transition via the regulation of the Smad3/Smad7 cascade. Am J Transl Res. 2017;9(6):2694–711.
Barcellos-Hoff MH, Lyden D, Wang TC. The evolution of the cancer niche during multistage carcinogenesis. Nat Rev Cancer. 2013;13(7):511–8.
Mizui Y, Sakai T, Iwata M, Uenaka T, Okamoto K, Shimizu H, Yamori T, Yoshimatsu K, Asada M. Pladienolides, new substances from culture of Streptomyces platensis Mer-11107. III. In vitro and in vivo antitumor activities. J Antibiot. 2004;57(3):188–96.
Effenberger KA, Anderson DD, Bray WM, Prichard BE, Ma N, Adams MS, Ghosh AK, Jurica MS. Coherence between cellular responses and in vitro splicing inhibition for the anti-tumor drug pladienolide B and its analogs. J Biol Chem. 2014;289(4):1938–47.
Lin X, Yi Z, Diao J, Shao M, Zhao L, Cai H, Fan Q, Yao X, Sun X. ShaoYao decoction ameliorates colitis-associated colorectal cancer by downregulating proinflammatory cytokines and promoting epithelial–mesenchymal transition. J Transl Med. 2014;12:105.
Smit MA, Peeper DS. Deregulating EMT and senescence: double impact by a single twist. Cancer Cell. 2008;14(1):5–7.
Ansieau S, Bastid J, Doreau A, Morel AP, Bouchet BP, Thomas C, Fauvet F, Puisieux I, Doglioni C, Piccinin S, et al. Induction of EMT by twist proteins as a collateral effect of tumor-promoting inactivation of premature senescence. Cancer Cell. 2008;14(1):79–89.
Kaplan RN, Rafii S, Lyden D. Preparing the “soil”: the premetastatic niche. Can Res. 2006;66(23):11089–93.
Psaila B, Lyden D. The metastatic niche: adapting the foreign soil. Nat Rev Cancer. 2009;9(4):285–93.
Joyce JA, Pollard JW. Microenvironmental regulation of metastasis. Nat Rev Cancer. 2009;9(4):239–52.
Lakkaraju A, Rodriguez-Boulan E. Itinerant exosomes: emerging roles in cell and tissue polarity. Trends Cell Biol. 2008;18(5):199–209.
Liu Y, Xiang X, Zhuang X, Zhang S, Liu C, Cheng Z, Michalek S, Grizzle W, Zhang HG. Contribution of MyD88 to the tumor exosome-mediated induction of myeloid derived suppressor cells. Am J Pathol. 2010;176(5):2490–9.
Becker A, Thakur BK, Weiss JM, Kim HS, Peinado H, Lyden D. Extracellular vesicles in cancer: cell-to-cell mediators of metastasis. Cancer Cell. 2016;30(6):836–48.
Martinez FO, Sica A, Mantovani A, Locati M. Macrophage activation and polarization. Front Biosci J Virtual Library. 2008;13:453–61.
Bai J, Adriani G, Dang TM, Tu TY, Penny HX, Wong SC, Kamm RD, Thiery JP. Contact-dependent carcinoma aggregate dispersion by M2a macrophages via ICAM-1 and beta2 integrin interactions. Oncotarget. 2015;6(28):25295–307.
Adriani G, Bai J, Wong S-C, Kamm R, Thiery J. M2a macrophages induce contact-dependent dispersion of carcinoma cell aggregates. Macrophage. 2016;3:e1222.
Condeelis J, Pollard JW. Macrophages: obligate partners for tumor cell migration, invasion, and metastasis. Cell. 2006;124(2):263–6.
Ong SM, Tan YC, Beretta O, Jiang D, Yeap WH, Tai JJ, Wong WC, Yang H, Schwarz H, Lim KH, et al. Macrophages in human colorectal cancer are pro-inflammatory and prime T cells towards an anti-tumour type-1 inflammatory response. Eur J Immunol. 2012;42(1):89–100.
Ostrand-Rosenberg S, Sinha P, Beury DW, Clements VK. Cross-talk between myeloid-derived suppressor cells (MDSC), macrophages, and dendritic cells enhances tumor-induced immune suppression. Semin Cancer Biol. 2012;22(4):275–81.
Toh B, Wang X, Keeble J, Sim WJ, Khoo K, Wong WC, Kato M, Prevost-Blondel A, Thiery JP, Abastado JP. Mesenchymal transition and dissemination of cancer cells is driven by myeloid-derived suppressor cells infiltrating the primary tumor. PLoS Biol. 2011;9(9):e1001162.
Gajewski TF, Schreiber H, Fu YX. Innate and adaptive immune cells in the tumor microenvironment. Nat Immunol. 2013;14(10):1014–22.
Wu Y, Deng J, Rychahou PG, Qiu S, Evers BM, Zhou BP. Stabilization of snail by NF-kappaB is required for inflammation-induced cell migration and invasion. Cancer Cell. 2009;15(5):416–28.
Li CW, Xia W, Huo L, Lim SO, Wu Y, Hsu JL, Chao CH, Yamaguchi H, Yang NK, Ding Q, et al. Epithelial–mesenchymal transition induced by TNF-alpha requires NF-kappaB-mediated transcriptional upregulation of Twist1. Can Res. 2012;72(5):1290–300.
Blackwell TS, Christman JW. The role of nuclear factor-kappa B in cytokine gene regulation. Am J Respir Cell Mol Biol. 1997;17(1):3–9.
Mantovani A, Romero P, Palucka AK, Marincola FM. Tumour immunity: effector response to tumour and role of the microenvironment. Lancet. 2008;371(9614):771–83.
Yang J, Weinberg RA. Epithelial–mesenchymal transition: at the crossroads of development and tumor metastasis. Dev Cell. 2008;14(6):818–29.
Hou F, Li W, Shi Q, Li H, Liu S, Zong S, Ren J, Chai J, Xu J. Yi Ai Fang, a traditional Chinese herbal formula, impacts the vasculogenic mimicry formation of human colorectal cancer through HIF-1alpha and epithelial mesenchymal transition. BMC Complementary Altern Med. 2016;16(1):428.
Huang M, Xin W. Matrine inhibiting pancreatic cells epithelial–mesenchymal transition and invasion through ROS/NF-kappaB/MMPs pathway. Life Sci. 2018;192:55–61.
Espada J, Peinado H, Lopez-Serra L, Setien F, Lopez-Serra P, Portela A, Renart J, Carrasco E, Calvo M, Juarranz A, et al. Regulation of SNAIL1 and E-cadherin function by DNMT1 in a DNA methylation-independent context. Nucleic Acids Res. 2011;39(21):9194–205.
Peinado H, Ballestar E, Esteller M, Cano A. Snail mediates E-cadherin repression by the recruitment of the Sin3A/histone deacetylase 1 (HDAC1)/HDAC2 complex. Mol Cell Biol. 2004;24(1):306–19.
Lin T, Ponn A, Hu X, Law BK, Lu J. Requirement of the histone demethylase LSD1 in Snai1-mediated transcriptional repression during epithelial–mesenchymal transition. Oncogene. 2010;29(35):4896–904.
Herranz N, Pasini D, Diaz VM, Franci C, Gutierrez A, Dave N, Escriva M, Hernandez-Munoz I, Di Croce L, Helin K, et al. Polycomb complex 2 is required for E-cadherin repression by the Snail1 transcription factor. Mol Cell Biol. 2008;28(15):4772–81.
Cano A, Nieto MA. Non-coding RNAs take centre stage in epithelial-to-mesenchymal transition. Trends Cell Biol. 2008;18(8):357–9.
Gregory PA, Bert AG, Paterson EL, Barry SC, Tsykin A, Farshid G, Vadas MA, Khew-Goodall Y, Goodall GJ. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat Cell Biol. 2008;10(5):593–601.
Park SM, Gaur AB, Lengyel E, Peter ME. The miR-200 family determines the epithelial phenotype of cancer cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes Dev. 2008;22(7):894–907.
Kim NH, Kim HS, Li XY, Lee I, Choi HS, Kang SE, Cha SY, Ryu JK, Yoon D, Fearon ER, et al. A p53/miRNA-34 axis regulates Snail1-dependent cancer cell epithelial–mesenchymal transition. J Cell Biol. 2011;195(3):417–33.
Siemens H, Jackstadt R, Hunten S, Kaller M, Menssen A, Gotz U, Hermeking H. miR-34 and SNAIL form a double-negative feedback loop to regulate epithelial–mesenchymal transitions. Cell Cycle (Georgetown, Tex). 2011;10(24):4256–71.
Hirata H, Hinoda Y, Shahryari V, Deng G, Tanaka Y, Tabatabai ZL, Dahiya R. Genistein downregulates onco-miR-1260b and upregulates sFRP1 and Smad4 via demethylation and histone modification in prostate cancer cells. Br J Cancer. 2014;110(6):1645–54.
Warzecha CC, Sato TK, Nabet B, Hogenesch JB, Carstens RP. ESRP1 and ESRP2 are epithelial cell-type-specific regulators of FGFR2 splicing. Mol Cell. 2009;33(5):591–601.
Sun Z, Cao Y, Hu G, Zhao J, Chen M, Wang S, Ye Z, Chen H, Wang W, Wang Y. Jinfu’an decoction inhibits invasion and metastasis in human lung cancer cells (H1650) via p120ctn-mediated induction and kaiso. Med Sci Monit. 2018;24:2878–86.
Alexandrov LB, Nik-Zainal S, Wedge DC, Aparicio SA, Behjati S, Biankin AV, Bignell GR, Bolli N, Borg A, Borresen-Dale AL, et al. Signatures of mutational processes in human cancer. Nature. 2013;500(7463):415–21.
Topalian SL, Hodi FS, Brahmer JR, Gettinger SN, Smith DC, McDermott DF, Powderly JD, Carvajal RD, Sosman JA, Atkins MB, et al. Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N Engl J Med. 2012;366(26):2443–54.
Huang S, Susztak K. Epithelial plasticity versus EMT in kidney fibrosis. Trends Mol Med. 2016;22(1):4–6.
Chen SR, Chen XP, Lu JJ, Wang Y, Wang YT. Potent natural products and herbal medicines for treating liver fibrosis. Chin Med. 2015;10:7.
Wang QL, Tao YY, Yuan JL, Shen L, Liu CH. Salvianolic acid B prevents epithelial-to-mesenchymal transition through the TGF-beta1 signal transduction pathway in vivo and in vitro. BMC Cell Biol. 2010;11:31.
Wang QL, Yuan JL, Tao YY, Zhang Y, Liu P, Liu CH. Fuzheng Huayu recipe and vitamin E reverse renal interstitial fibrosis through counteracting TGF-beta1-induced epithelial-to-mesenchymal transition. J Ethnopharmacol. 2010;127(3):631–40.
Li CX, Li L, Lou J, Yang WX, Lei TW, Li YH, Liu J, Cheng ML, Huang LH. The protective effects of traditional Chinese medicine prescription, han-dan-gan-le, on CCl4-induced liver fibrosis in rats. Am J Chin Med. 1998;26(3–4):325–32.
Jiao D, Wang J, Lu W, Tang X, Chen J, Mou H, Chen QY. Curcumin inhibited HGF-induced EMT and angiogenesis through regulating c-Met dependent PI3 K/Akt/mTOR signaling pathways in lung cancer. Mol Ther Oncolytics. 2016;3:16018.
Lou W, Chen Y, Zhu KY, Deng H, Wu T, Wang J. Polyphyllin I overcomes EMT-associated resistance to erlotinib in lung cancer cells via IL-6/STAT3 pathway inhibition. Biol Pharm Bull. 2017;40(8):1306–13.
Pei Z, Fu W, Wang G. A natural product toosendanin inhibits epithelial–mesenchymal transition and tumor growth in pancreatic cancer via deactivating Akt/mTOR signaling. Biochem Biophys Res Commun. 2017;493(1):455–60.
Li M, Li P, Zhang M, Ma F. Brucine suppresses breast cancer metastasis via inhibiting epithelial mesenchymal transition and matrix metalloproteinases expressions. Chin J Integr Med. 2018;24(1):40–6.
Wu TT, Lu J, Zheng PQ, Liu SL, Wu J, Sun W, Sun QM, Ma NX, Ding XL, Chen M, et al. Yiqi Huayu Jiedu decoction inhibits the invasion and metastasis of gastric cancer cells through TGF-beta/Smad pathway. Evid Based Complementary Altern Med eCAM. 2017;2017:1871298.
Acknowledgements
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
JB, JPT organized the outline, drafted the manuscript and designed the figures. WCK contributed to the final version of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Bai, J., Kwok, W.C. & Thiery, JP. Traditional Chinese Medicine and regulatory roles on epithelial–mesenchymal transitions. Chin Med 14, 34 (2019). https://doi.org/10.1186/s13020-019-0257-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13020-019-0257-6