
Abstract
Background
Soybean mosaic virus (SMV), a Potyvirus, is the most prevalent viral pathogen of soybean that causes severe yield and seed quality reductions in world soybean production. So far, multiple resistance loci for different SMV strains have been fine-mapped while the candidate genes’ functions need to be verified. However, identification of the resistance or susceptibility genes via stable genetic transformation is time-consuming and labor-intensive, which hinders further exploration of these genes’ functions in soybean. Thus we tried to explore a rapid and efficient method for verification of SMV-related target gene function in soybean.
Results
An Agrobacterium rhizogenes (A. rhizogenes) induced soybean hairy roots versus Soybean mosaic virus (ARISHR-SMV) pathosystem was established. The procedure is characterized with that (1) the soybean hairy roots that can be infected by SMV are induced by A. rhizogenes K599 using soybean cotyledons as explants, (2) the gene to be examined for its function, which may be the endogenous SMV-resistance or -susceptible gene or exogenous SMV-related gene, is transformed into soybean calluses mediated by A. rhizogenes, (3) the transformed calluses on explants further inoculated with the purified tester-SMV virions using pricking method under aseptic conditions, and (4) the measurement of the SMV content in positive hairy roots for evaluating the SMV-related target gene function. The procedure takes about 30 days for one cycle. Utilizing the established procedure, the soybean hairy roots induced by A. rhizogenes was efficiently infected by multiple different SMV strains, the SMV infectivity in soybean hairy roots can be retained at least twice successive transfer cultures and Tomato bushy stunt virus (TBSV) P19 promoting and SMV CP suppressing SMV infection in soybean hairy roots was demonstrated, respectively. This procedure can also be used for identification of resistance to SMV strains for soybean germplasms.
Conclusion
The ARISHR-SMV is an efficient pathosystem that allows a quick and convenient identification of SMV-related target gene function in soybean.
Similar content being viewed by others
Background
Soybean originated in China is a dominant source of plant protein and oil with multiple uses in food, feed, and industrial applications in the world. It usually subjects to various challenges from biotic and abiotic stresses during its whole growth period under natural environments. Among the biotic stresses, Soybean mosaic virus (SMV) disease is the major threat in all soybean-production areas in the world, especially in China, and often leads to serious yield losses (ranging from 8 to 100%) and seed quality reductions [1,2,3]. SMV belongs to Potyvirus with an approximately 9.6 kb, single-stranded, positive-sense RNA genome that contains a known open reading frame encoding at least 11 proteins and 5′ viral protein genome-linked (VPg) cap and 3′ poly(A) tail [4, 5]. A mass of SMV isolates were identified worldwide. Seven SMV strains (G1–G7) isolated from germplasm collections were reported in the United States based on the responses to a set of eight differential soybean genotypes [6]. Similarly, five SMV strains (A–E) in Japan and eleven SMV strains (G1–G7, SMV-N, G5H, G7a and G7H) in Korea have been reported [7, 8]. In China, more than 4500 country-wide SMV isolates were collected and grouped into twenty-two strains based on their responses to ten differential soybean varieties at the National Center for Soybean Improvement [9,10,11,12,13,14].
Host plant resistance is an effective, economic, and environment-friendly strategy in studying resistance to SMV. One of the prerequisites to utilize host plant resistance is to screen out SMV resistance sources to be used for gene mapping, candidate gene identification and breeding for cultivars resistant to SMV. A number of independent SMV resistance loci corresponding to specific strains have been identified in some resistance sources. For instance, in the United States three independent loci, Rsv1, Rsv3, Rsv4 and in China sixteen independent loci, Rsc3, Rsc4, Rsc5, Rsc6, Rsc7, Rsc8, Rsc10, Rsc11, Rsc12, Rsc13, Rsc14, Rsc15, Rsc17, Rsc18, Rsc20, Rsc21 have been identified from different resistance sources [2, 15, 16]. However, few of these genes has been cloned and their relationship has not been fully explored yet. In addition, the breakdown of R gene-mediated resistance occurred frequently with the rapid evolution in IPs/IPRs [17,18,19,20,21]. Therefore, it was urgent to establish a simple and effective system for validating the candidate genes’ functions as a quick excavation of SMV resistance in facing the co-evolution between SMV and soybean.
Recently, several new methods have been developed in verification of anti-SMV candidate genes’ functions. One approach is to transform a target gene obtained from the pathogen or host into soybean through agrobacterium-mediation or particle bombardment transformation [22,23,24]. However, there are still problems with the soybean genetic transformation, such as lack of easy-transformed genotypes, low transformation efficiency, imperfect regeneration system, time-consuming and labor-inefficiency, etc. which hinders the functional analysis of the candidate genes. Moreover, virus-induced gene silencing (VIGS) may substitute for stable transformation to rapidly analyze the function of the candidate genes as an effective method [25, 26]. For example, VIGS vectors based on Bean pod mottle virus have been used to identify components of the soybean resistance response against SMV [27, 28]. Although VIGS has been proven useful for identification of candidate genes’ functions and can overcome gene redundancy in soybean [29], it involves a viral vector which could mask or alter the defense responses of infected plants. On the other hand, the use of the hairy roots system which can be induced by transformation with a gram-negative soil bacterium A. rhizogenes might be also an alternative approach to study gene functions [30]. The morphological characteristics of A. rhizogenes-induced hairy roots were similar to that of wild-type roots, characterized with fast growth and lack of geotropism and lateral branching [31, 32]. The hairy roots were widely used as a transgenic tool to study secondary metabolism and gene function in plants. Lozovaya et al. [33] reported that down regulation of chalcone synthase or isoflavone synthase modified the phenolic metabolism in soybean hairy roots. In aspect of abiotic stress, soybean hairy roots were also applied to explore the genes responsible for salt and drought stress [34, 35]. While for biotic stresses, it has been used for propagating soybean parasites or pathogens and for testing interactions between soybean and the parasite or pathogen, for example, the soybean cyst nematode [36,37,38]. However, since SMV is a leaf disease, until now this approach has not been used in the study for SMV resistance yet.
The present study aimed at (1) establishing a ARISHR-SMV pathosystem that allows a rapid and effective identification of SMV-related target gene function in soybean under aseptic conditions, (2) demonstrating whether soybean hairy roots can be efficiently infected by different SMV strains, and (3) utilizing the established ARISHR-SMV pathosystem to confirm the biological function of TBSV P19 and SMV CP protein on SMV infection in soybean hairy roots.
Materials and methods
Plant materials and SMV strains
Soybean cultivar Williams and NN1138-2 were used for A. rhizogenes-mediated soybean cotyledon transformation and propagating SMV, respectively. Soybean plants (NN1138-2) were grown at 25 °C with a 16 h photoperiod in a greenhouse and the unifoliolate leaves of 10-day-old plants were used for virus inoculation. There are 9 strains (SC3, SC7, SC8, SC9, SC10, SC11, SC13, SC15 and SC18) tested in the present study, which were provided by the National Center for Soybean Improvement (Nanjing Agricultural University, Nanjing, China).
Culture of A. rhizogenes
The wild-type or transformed A. rhizogenes strain K599 was cultured in the LB medium (50 μg/ml kanamycin) and was further shaken in 4 ml LB broth (50 μg/ml kanamycin) at 200 rpm for 24 h at 28 °C. Before inoculation, the cultures were centrifuged at 4000 rpm for 5 min and then the tight pellets were gently re-suspended in 10 mM MgCl2. The bacterial suspension was diluted to OD600 = 0.5 with 100 mM MgCl2 containing 150 μM acetosyringone and gently shaken at 45 rpm for at least 30 min at 22 °C before use.
Agrobacterium rhizogenes-mediated soybean cotyledon transformation
The transformation of soybean cotyledons were performed essentially as the described procedures by Subramanian et al. [39] with some modifications. The mature soybean seeds (Williams) were surface-sterilized for 6 h using chlorine gas generated by mixing 3.5 ml of 12 N HCl with 100 ml sodium hypochlorite solution. The sterilized seeds were soaked overnight in sterilized water and the seed coats were carefully removed. The unshelled seeds were further germinated on agar (0.7%) medium in a plant growth incubator with a 16 h photoperiod. When cotyledons became green and just began to separate (after approximately 4 days), the intact cotyledons were harvested and cut into a roughly small circular (about 5 mm diameter) at the petiole end to be used for A. rhizogenes-mediated transformation. The cotyledon pieces were transferred to White’s medium in petri plates and immediately treated with 20 μl of the freshly prepared A. rhizogenes suspension. The plates were then wrapped in Parafilm and placed in an incubator at 25 °C under dark culture. After 10 days, the calluses grown from the incubated cotyledon pieces were used for virus inoculation.
SMV purification and inoculation of callus with virions
Purification of SMV and determination of the viral concentration were performed as described by Li and Pu [40]. The molecular weight of the capsid protein was estimated by SDS-PAGE on a 3.5% stacking gel and a 12% resolving gel. After electrophoresis for 2 h at 120 V at room temperature, the proteins were stained with coomassie brilliant blue. Inoculation of calluses were done aseptically by pricking the callus tissues with a sterile needle, adding 10 μl 0.05 M PB buffer or the purified SMV virions (50 μg/ml) to the wound sites. The inoculated explants were placed onto White’s medium and further cultured for growing soybean hairy roots.
Serological determination, RT-PCR detection and back-inoculation assays of soybean
The enzyme linked immunosorbent assay (ELISA) was used for serological determination of SMV infection. The SMV antiserum (polyclonal rabbit antibodies) was purchased from ACD Inc. (cat #V094-R1, Beijing, China) and the manufacturer’s instructions were followed for the next steps. The total RNA was extracted from soybean hairy roots using RNA isolation kit (Tiangen, Beijing, China) following the manufacturer’s instructions. The quality and concentration of RNA were checked using agarose gel electrophoresis and a spectrophotometer (Nanodrop 2000, Thermo Fisher Scientific), respectively. The first-strand cDNA was synthesized using M-MLV reverse transcriptase (Invitrogen) as recommended by the manufacturer, then the 731 bp viral fragment was amplified with SMV CP gene specific primers (forward primer 5′-GAAGGAGACATGGATGCAG-3′ and reverse primer 5′-CTTGCAGTGTGCCTTTCAG-3′), the soybean EF1α gene was used as the internal control and the forward primer 5′-AAGTCTGTTGAGATGCACCA-3′ and reverse primer 5′-GCCTGTCAATCTTGGTCAAG-3′ were used to amplify the 277 bp fragment. The soybean hairy roots grown from SMV-inoculated calluses were homogenized in 0.01 M phosphate buffer (PH 7.4) in different grinders and the tissue homogenates were back-inoculated the soybean NN1138-2 through mechanical inoculation.
Construction of the TBSV P19 and SMV CP overexpression vectors and transformation of A. rhizogenes
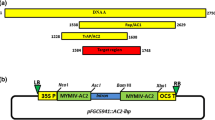
The P19 sequence of Tomato bushy stunt virus (TBSV) [41] and the CP sequence of SMV SC15 strain was cloned into pEarleyGate103 [42] using gateway method, respectively. Likewise, the green fluorescent protein (GFP) derived from pEarleyGate103 was cloned into pEarleyGate202 [42] and this recombinant vector served as control. Additional file 1: Fig. S1 showed the gene arrangement in the constructs. Primers required to construct these vectors are listed in Additional file 2: Table S1. The constructed vectors validated by sequencing were transformed into A. rhizogenes K599 strain by electroporation.
Identification of transgenic hairy roots
The genomic DNA was extracted from soybean hairy roots by cetyltrimethylammonium bromide method with minor modifications [43]. Insertion of the P19 and CP gene into soybean genome was confirmed by PCR using the genomic DNA of hairy root transformants, respectively. Forward primer 5′-ATGGAACGAGCTATACAAGGAA-3′ and reverse primer 5′-CTCGCTTTCTTTTTCGAAGGT-3′ were used to amplify the 516 bp P19 fragment. Forward primer 5′-GAAGGAGACATGGATGCAG-3′ and reverse primer 5′-CTTGCAGTGTGCCTTTCAG-3′ were used to amplify the 731 bp CP fragment. The GFP fluorescence of hairy roots transformed with the vector control, P19 or CP overexpression vector were monitored under stereo fluorescence microscope (OLYMPUS MVX10, Japan). The proteins of GFP-positive transgenic hairy roots were extracted and the protein concentrations were determined by the Bradford assay procedure using bovine serum albumin as the standard [44]: 10 μg protein samples were separated with 10% SDS-PAGE and then transferred onto a polyvinylidene difluoride membrane (Immobilon). Anti-GFP monoclonal antibody (1:3000; Sigma-Aldrich) and IRDye 800CW-conjugated goat (polyclonal) anti-mouse IgG (1:10,000; H + L; LI-COR Biosciences) secondary antibodies were used to further evaluate the gene expression in soybean hairy roots. The membranes were visualized using a LI-COR Odyssey scanner with excitation at 700 and 800 nm.
Real-time quantification of viral RNA
The quantitative real-time polymerase chain reaction (qRT-PCR) was performed using a 7500 Real-time PCR system (Applied Biosystems) in conjunction with SYBR green reagents (TAKARA, Dalian, China). The qRT-PCR analysis of P3 gene of SMV strain SC15 was carried out using the following primers: 5′-GAAAGTTGTAGAAGAAAGCAG-3′ (forward) and 5′-ACAAAGGCTCGCACCGATGC-3′ (reverse). The soybean ELF1β was used as an internal control with the following primers: 5′-GTTGAAAAGCCAGGGGACA-3′ (forward) and 5′-TCTTACCCCTTGAGCGTGG-3′ (reverse), in a 20 μl reaction, including 20 ng cDNA, 0.4 μl of each 10 μM primer, 0.4 μl ROX Reference Dye П (50×), 10 μl Premix Ex Taq (2×) and 6 μl dH2O. Three technical replications in each of the two biological replicates for each sample were taken for real-time PCR analysis. The relative RNA accumulation were calculated using 7500 software (version 2.0.6) supplied by the manufacturer.
Statistical analyses
The ELISA and qRT-PCR data were analyzed for their means, standard deviations and single-factor analyses of variance using the SPSS software (version 18.0). The Fisher’ protected least significant difference tests were performed for comparisons between treated group mean and control group means using the significance level of α = 0.05.
Results
Establishment of the ARISHR-SMV pathosystem
The induction and culture of soybean hairy roots under aseptic condition is shown in Fig. 1. The processes are as the following: the soybean seeds are placed into a desiccator and surface-sterilized using chlorine gas (Fig. 1a); the peeled seeds germinate on 7% agar medium and the seedlings grow to about 6 cm in height, while the two cotyledons have not yet unfolded on about the 4th day (Fig. 1b); a ridge of callus emerges along the severed vein after inoculating the cut surface with A. rhizogenes strain K599 on about the 4th day (Fig. 1c); the yellowish-white callus proliferates along the vein on the 10th day (Fig. 1d) and the hairy roots begin to form on the 12th day (Fig. 1e); then abundant hairy roots form on the 15th day (Fig. 1f), with more new hairy roots begin to emerge (Fig. 1g); and the hairy roots cut from cotyledons grow strongly on White’s basal medium (Fig. 1h).

The transformation procedure and culture of soybean hairy roots under aseptic conditions. a The spotless soybean seeds were surface sterilized using chlorine gas in a desiccator. b The peeling seeds grow to about 6 cm high in 7% agar medium. The performance of cotyledon tissues on 4 days (c), 10 days (d), 12 days (e), 15 days (f), and 19 days (g) after infection with A. rhizogenes strain K599. h Soybean hairy roots grown from calluses were cut and transferred to White’s medium
To establish the ARISHR-SMV pathosystem, the soybean calluses induced by A. rhizogenes K599 on soybean cotyledon explants (for example, that of the cultivar Williams) are used for SMV inoculation, while the purified SMV strain (for example, SC15) virions are used as tester-inoculum. Whether the virus exist or not in hairy roots grown from SMV-inoculated calluses are measured. As shown in Fig. 2, the yellowish-white calluses were pricked with a fine sterile needle at the black arrow position in Fig. 2a and further inoculated with mock or the purified SMV virions as shown in Fig. 2b. After 14 days post inoculation (dpi), the extracts of hairy roots grown from mock- or SC15-inoculated calluses were assayed using ELISA with polyclonal antibodies to detect SMV coat protein. This revealed that SMV was present at high concentrations in the hairy roots grown from these calluses inoculated with SC15, even the viral content levels in soybean hairy roots are less than those in leaves (Fig. 2c). Meanwhile, the presence of viral RNA in hairy roots was measured by RT-PCR using SMV-specific primers. The agarose gel electrophoresis showed the presence of the 731 bp amplicon in hairy roots inoculated with strain SC15 (Fig. 2d). Moreover, the susceptible soybean line NN1138-2 back-inoculated with the sap from SMV infected hairy roots developed typical mosaic symptoms (Fig. 2f) and no symptoms were observed in mock control following mechanical inoculation at 14 dpi (Fig. 2e). As shown in Fig. 3, the entire process takes about 1 month, including 5 days for preparation of the soybean explant, 10 days for growth of soybean callus with which the growth status can just fit for SMV inoculation and 15 days for the period from virus inoculation to harvest of the infected soybean hairy roots.

Calluses inoculation process with SMV and back-inoculation soybean assays with hairy roots sap. a 10 days yellowish-white calluses were pricked with a fine sterile needle at black arrow position. b SDS-PAGE electrophoresis of the purified SMV strain SC15 (an asterisk indicates the positions of SMV capsid protein). c ELISA detection of SMV coat protein in un-, Mock- and virions-inoculated soybean hairy roots with diseased soybean leaves at 14 dpi. The error bars represent standard deviations with N = 10. d Accumulation of viral RNA were measured by RT-PCR at 14 dpi. e, f Plants were back-inoculated with sap from hairy roots grown from Mock- and SMV-inoculated calluses, respectively, and the photographs were taken at 14 dpi

Schematic view of the established ARISHR-SMV pathosystem
Multiple SMV strains can infect soybean hairy roots using the ARISHR-SMV pathosystem
To know whether other SMV strains can infect hairy roots through this ARISHR-SMV pathosystem method, the purified SMV strains SC3, SC7, SC8, SC9, SC10, SC11, SC13 and SC18 virions were used as tester to inoculate soybean calluses, respectively, and the hairy roots grown from these calluses were further assayed with ELISA. The results revealed that SMV was detected in all the soybean hairy roots grown from the calluses inoculated with different SMV strains (Table 1). All SMV-infected soybean hairy roots gave 85.7–100% infection in infectivity assay through back-inoculating the soybean line NN1138-2 (Table 1). As expected, the mock-inoculated hairy roots exhibited negative reactions in serological tests (Table 1). Taken together, the above results suggested that all the 9 SMV strains can be applied to the ARISHR-SMV pathosystem. Besides, the pathosystem can be used also for identification of resistance to these 9 SMV strains in soybean germplasms.
Retention of SMV infectivity in hairy roots with successive transfer cultures
To further develop this ARISHR-SMV pathosystem method, the key aspects of the interaction between SMV and soybean hairy root cells in vitro was investigated. The maintenance of viral stability and infectivity in hairy roots was especially concerned. The SMV infectivity was measured through back-inoculating soybeans with the sap obtained from soybean hairy roots which were transferred to White’s medium one or two times. The results showed that the virus in a majority of hairy roots was infective after one or two successive transfer cultures, it produced 70.2–86.6% infected plants on average for different SMV strains (Table 2). In addition, viral infectivity rate was similar as the results between the two transfers (Table 2). As expected, no virus was detected in the non-inoculated control by infectivity assay (Table 2). The above results indicated that the SMV infectivity in soybean hairy roots can be retained at least twice successive transfer cultures.
Validation of TBSV-P19 promoting and SMV-CP suppressing SMV infection in soybean hairy roots using ARISHR-SMV pathosystem
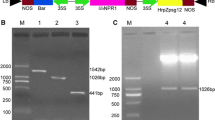
The P19 of TBSV was a suppressor of gene silencing in plants [41, 45], which prevents the onset of post-transcriptional gene silencing that is natural plant antiviral mechanism in eukaryotes, and promotes pathogen infection in the tissue-specific expression of P19 gene. On the other hand, the previous research reported that overexpression of SMV CP gene in soybean enhanced the transgenic plants’ resistance to SMV [23, 46]. The mechanism of pathogen-derived resistance (PDR) was reported involving post-transcriptional RNA silencing [47, 48] or the CP itself [49, 50]. In order to test the ARISHR-SMV pathosystem, we introduced the TBSV P19 and SMV CP gene into the soybean hairy roots separately and purified the SMV strain SC15 as tester. After just 10 days, transformation of the callus with appropriate vector control, P19 and CP overexpression vector were monitored with fluorescence of the GFP marker (Fig. 4a). The callus which can produce more than five fluorescent spots was used to inoculate strain SC15 (Fig. 4a), otherwise, it was discarded. Among the 4 roots shown in Fig. 4b, two of them had been transformed by K599 carrying a P19 or CP overexpression vector (Fig. 4c). The remained two might come from emerged adventitious roots or the transformation of the wild-type K599 strain. We observed that approximately 30% of the hairy roots were successfully transformed with P19 or CP overexpression vector on average. The time or efficiency of root formation was not changed when vector control, P19 or CP overexpression vector presents in K599. Meanwhile, the gDNA PCR results showed the presence of the 516 bp and 731 bp amplicon in transgenic hairy roots using specific primers of P19 or CP gene, respectively (Fig. 4d). The western blot results (Fig. 4e) confirmed the fluorescence observed in Fig. 4c. For determining whether P19 promoted and CP suppressed the accumulation of SMV strain SC15 in transgenic hairy roots, the virus content in positive hairy roots was measured at 10, 15 and 20 dpi using qRT-PCR, respectively. The results revealed that SMV concentration was significantly higher in P19 transgenic hairy roots and lower in CP transgenic hairy roots than that in control at 10, 15 and 20 dpi, respectively (Fig. 4f). Taken together, the ARISHR-SMV pathosystem has certain potential for rapid and effective identification of exogenous SMV-related candidate gene function, or endogenous soybean candidate resistance gene or susceptibility gene function in soybean under aseptic conditions. Meanwhile, pre-overexpression of the P19 in soybean hairy roots was an optional way of enhancing the ability of SMV infection.

Verification of P19 and CP transformation events and the effect of P19 and CP on SMV accumulation in soybean hairy roots, respectively. a Soybean calluses were induced by K599 carrying control or destination vector on 10 days under stereo fluorescence microscope. Soybean hairy roots of a cotyledon transformed with control or destination vector are shown on 15 days under white light (b) and long wavelength UV light with a GFP barrier filter (c). d gDNA PCR detection of P19 and CP amplicon in transgenic hairy roots. NT: no template; EV: vector control; +: plasmid; 1#, 2#, 3#: different lines of transgenic hairy roots. e Western blot confirm the presence of P19 and CP protein in hairy roots. U: no plasmid; EV: vector control; P19: P19 plasmid; CP: CP plasmid. f Amount of viral RNA accumulating in hairy roots at 10, 15, and 20 dpi. The error bars indicate standard deviations calculated from three technical replications
Discussion
The ARISHR-SMV pathosystem is a rapid and efficient method for verification of SMV-related target gene function in soybeans
In recent years, the hairy roots system has been extensively used to study the gene functions of soybeans in different cases. For example, Li et al. [34] reported that GmNAC15 overexpression in hairy roots enhanced salt tolerance in soybean. Wang et al. [35] reported that GmWRKY27 interacted with GmMYB174 to reduced the expression of GmNAC29 for drought tolerance in soybean hairy roots. Liu et al. [36] identified a soybean cyst nematode resistance gene using hairy roots system which can be used to reduce soybean yield losses. However, soybean hairy root system has not been used in the study for SMV infection. In the present study, the ARISHR-SMV pathosystem was established and can be used for studying soybean-virus interactions in hairy roots, which can broaden the research scope of hairy roots system (Figs. 1, 2, 3, 4). Recently, Morriss et al. [37] developed an interaction system between soybean hairy roots and aphid that allowed effective aphid feeding and validation of aphids resistance genes. Combining the present results with the reports of Morriss et al. [37], all three factors (soybean, SMV and aphid) would be integrated into the hairy roots system to study the interaction relationship among them. Meanwhile, the ARISHR-SMV pathosystem just need to take about 1 month for the entire flow and allowed rapid identification of the target genes responsible for SMV resistance or susceptibility, which is more efficient than the normal soybean genetic transformation when screening a mass of SMV-related target genes’ functions. Prior to this, Jiang et al. [51] reported a SMV variant belonging to SMV SC7 strain caused different diseases in Glycine max and Nicotiana benthamiana, and the interaction system between SMV and Nicotiana benthamiana was an ideal model for investigating the resistance genes to SC7 strain, but it is limited merely to the specific strain. Compared with the results of Jiang et al. [51], the newly established ARISHR-SMV pathosystem can be applied for at least 9 SMV strains, therefore, it is characterized with multiple uses in the study of SMV resistance or susceptibility genes (Tables 1 and 2). Latterly, two methods of transiently expressing a target gene using Agrobacterium infiltration in soybean leaf tissue were developed, however, the small amount of infectious tissues and the low levels of replicated experiments may have limited their practical implications [52, 53]. Additionally, Wu and Hanzawa [54] reported that a protocol for the isolation of soybean protoplasts and the application to transient expression studies, while it is not suitable for observation of long term cellular processes. In ARISHR-SMV pathosystem, induction and culture of soybean hairy roots are under aseptic condition, the transformation efficiency of hairy roots is controllable and SMV infectivity can be maintained in hairy roots with successive transfers (Figs. 1, 2, 3, 4, Tables 1, 2). Thus, the ARISHR-SMV pathosystem can be used for observation of gene effect during long-term process of SMV infection which provide convenience for the study on disease-related candidate gene functions. Besides, SMV and its counterpart soybean hairy roots fit well the Invasion Model in which host receptors, termed invasion pattern receptors (IPRs) detect either an externally encoded or modified-self ligand that indicates invasion, termed invasion pattern(s) [IP(s)] [21] and therefore the ARISHR-SMV pathosystem can be used to excavate not only new IPRs but also new IPs in SMV-soybean competition, which supply an opportunity for rapid study the molecular mechanisms of soybean resistance to SMV.
The ARISHR-SMV pathosystem can be used also for SMV resistance identification of soybean germplasm
Different from the previous method of resistance identification of soybean cultivars to SMV, which evaluating the whole performance of the inoculated plants to assess the resistance or susceptibility of soybean germplasm to SMV [55]. The present study utilizing A. rhizogenes to induce calluses of a certain soybean genotype for inoculating the purified SMV virions and estimating varietal resistance according to the SMV presence of the inoculated soybean calluses or hairy roots (Figs. 1, 2, 3). With this method, a soybean plant (genotype) can be used for multiple SMV strains since a callus can be separated into multiple parts while the conventional method has to spend one plant for one strain, which is impossible if each plant is a unique genotype. Moreover, the previous method was greatly affected by climate, especially the outside temperature, therefore, it cannot be carried out in the whole year, while the ARISHR-SMV method can save manpower and space costs, and is free from the influences of temperature and humidity, therefore, can be performed at any time with more flexibility. However, this method also has some shortcomings in SMV resistance identification of soybean germplasm, i.e., the infection of A. rhizogenes will induce plant defense response and changes in hormone levels, which could lead to a synergistical effect with SMV infection [56]. As a consequence, the results of resistance identification of soybean cultivars to SMV will be possible to appear false positive. Even so, the ARISHR-SMV pathosystem still supply an additional and aided approach for SMV resistance identification in soybean. For a broad application of this new method, whether soybean hairy roots can be infected by more SMV strains needs to be further investigated.
Conclusions
A pathosystem, designated ARISHR-SMV, is established for a rapid and effective identification of SMV-related target gene function, which composes of utilizing A. rhizogenes transformed with the target gene to induce calluses, inoculating the calluses with SMV virions and measuring the viral content in hairy roots derived from the calluses. The soybean hairy roots can be efficiently infected by different SMV strains and SMV infectivity in soybean hairy roots can be retained at least twice successive transfer cultures. In addition, this ARISHR-SMV pathosystem was successfully applied to confirm the function of P19 and CP protein on SMV infection in soybean hairy roots, respectively. This method provided a new option for investigating SMV resistance or susceptibility gene function in soybean.
Availability of data and materials
The data sets supporting the results of this article are included within the article.
Abbreviations
- SMV:
-
Soybean mosaic virus
- A. rhizogenes :
-
Agrobacterium rhizogenes
- ARISHR-SMV:
-
Agrobacterium rhizogenes induced soybean hairy roots-Soybean mosaic virus
- VIGS:
-
virus-induced gene silencing
- TBSV:
-
Tomato bushy stunt virus
- ELISA:
-
enzyme linked immunosorbent assay
- qRT-PCR:
-
quantitative real-time polymerase chain reaction
- dpi:
-
days post-inoculation
References
Hill JH, Whitham SA. Control of virus diseases in soybeans. Adv Virus Res. 2014;90(90):355–90.
Li K, Zhi HJ. Advances in resistance to Soybean mosaic virus disease in soybean. Soybean Sci. 2016;35(4):525–30.
Hill JH. Soybean mosaic virus. In: Hartman GL, Sinclair JB, Rupe JC, editors. Compendium of Soybean diseases. 4th ed. St. Paul: APS Press; 1999. p. 70–1.
Jayaram C, Hill JH, Miller WA. Complete nucleotide sequences of two Soybean mosaic virus strains differentiated by response of soybean containing the Rsv resistance gene. J Gen Virol. 1992;73:2067–77.
Wen RH, Hajimorad MR. Mutational analysis of the putative pipo of Soybean mosaic virus suggests disruption of PIPO protein impedes movement. Virology. 2010;400(1):1–7.
Cho EK, Goodman RM. Strains of Soybean mosaic virus: classification based on virulence in resistant soybean cultivars. Phytopathology. 1979;69:467–90.
Takahashi K, Tanaka T, Iida W, Tsuda Y. Studies on virus diseases and causal viruses of soybean in Japan. Bull Tohoku Natl Agric Exp Stn. 1980;62:1–130.
Seo JK, Lee HG, Choi HS, Lee SH, Kim KH. Infectious in vivo transcripts from a full-length clone of Soybean mosaic virus strain G5H. Plant Pathol J. 2009;25(1):54–61.
Li K, Yang QH, Zhi HJ, Gai JY. Identification and distribution of Soybean mosaic virus strains in southern China. Plant Dis. 2010;94(3):351–7.
Wang DG, Li HW, Zhi HJ, Tian Z, Hu C, Hu GY, et al. Identification of strains and screening of resistance resources to Soybean mosaic virus in Anhui Province. Chin J Oil Crop Sci. 2014;36:374–9.
Guo DQ, Zhi HJ, Wang YW, Gai JY, Zhou XA, Yang CL, et al. Identification and distribution of Soybean mosaic virus strains in Middle and Northern Huang Huai Region of China. Chin J Oil Crop Sci. 2005;27(4):64–8.
Wang XQ, Gai JY, Pu ZQ. Classification and distribution of strain groups of Soybean mosaic virus in middle and lower Huang-Huai and Changjiang valleys. Soybean Sci. 2003;22(2):102–7.
Wang YW, Zhi HJ, Guo DQ, Gai JY, Chen QS, Li K, et al. Classification and distribution of strain groups of Soybean mosaic virus in northern China spring planting soybean region. Soybean Sci. 2005;24:263–8.
Zhan Y, Zhi HJ, Yu DY, Gai JY. Identification and distribution of SMV strains in Huang-Huai valleys. Sci Agric Sin. 2006;39(10):2009–15.
Liu JZ, Fang Y, Pang HX. The current status of the soybean-Soybean mosaic virus (SMV) pathosystem. Front Microbiol. 2016;7:1906.
Karthikeyan A, Li K, Jiang H, Ren R, Li C, Zhi HJ, et al. Inheritance, fine-mapping, and candidate gene analyses of resistance to Soybean mosaic virus strain SC5 in soybean. Mol Genet Genom. 2017;292(4):811–22.
Saruta M, Kikuchi A, Okabe A, Sasaya T. Molecular characterization of A2 and D strains of Soybean mosaic virus, which caused a recent virus outbreak in soybean cultivar Sachiyutaka in Chugoku and Shikoku regions of Japan. J Gen Plant Pathol. 2005;71(6):431–5.
Koo JM, Choi BK, Ahn HJ, Yum HJ, Choi CW. First report of an Rsv resistance-breaking isolate of Soybean mosaic virus in Korea. Plant Pathol. 2005;54(4):573–573.
Choi BK, Koo JM, Ahn HJ, Yum HJ, Choi CW, Ryu KH, et al. Emergence of Rsv-resistance breaking Soybean mosaic virus isolates from Korean soybean cultivars. Virus Res. 2005;112(1):42–51.
Gagarinova AG, Babu M, Poysa V, Hill JH, Wang AM. Identification and molecular characterization of two naturally occurring Soybean mosaic virus isolates that are closely related but differ in their ability to overcome Rsv4 resistance. Virus Res. 2008;138(1):50–6.
Cook DE, Mesarich CH, Thomma BPHJ. Understanding plant immunity as a surveillance system to detect invasion. Annu Rev Phytopathol. 2015;53:541–63.
Wang XY, Eggenberger AL, Nutter FW Jr, Hill JH. Pathogen-derived transgenic resistance to Soybean mosaic virus in soybean. Mol Breed. 2001;8(2):119–27.
Furutani N, Hidaka S, Kosaka Y, Shizukawa Y, Kanematsu S. Coat protein gene-mediated resistance to soybean mosaic virus in transgenic soybean. Breed Sci. 2006;56(2):119–24.
Kim HJ, Kim MJ, Pak JH, Jung HW, Choi HK, Lee YH, et al. Characterization of SMV resistance of soybean produced by genetic transformation of SMV-CP gene in RNAi. Plant Biotechnol Rep. 2013;7(4):425–33.
Lacomme C. Strategies for altering plant traits using virus-induced gene silencing technologies. Methods Mol Biol. 2015;1287:25–41.
Pflieger S, Richard MMS, Blanchet S, Meziadi C, Geffroy V. VIGS technology: an attractive tool for functional genomics studies in legumes. Funct Plant Biol. 2013;40(12):1234–48.
Zhang CQ, Grosic S, Whitham SA, Hill JH. The requirement of multiple defense genes in soybean Rsv1-mediated extreme resistance to Soybean mosaic virus. Mol Plant Microbe Interact. 2012;25(10):1307–13.
Huang SH, Zheng GJ, Yang YQ, Zhi HJ. Analysis on the candidate resistance gene GmZ15 to Soybean mosaic virus by VIGS. Soybean Sci. 2015;26(26):400–7.
Liu JZ, Graham MA, Pedley KF, Whitham SA. Gaining insight into soybean defense responses using functional genomics approaches. Brief Funct Genom. 2015;14(4):283–90.
Veena V, Taylor CG. Agrobacterium rhizogenes: recent developments and promising applications. Vitro Cell Dev Biol Plant. 2007;43(5):383–403.
Shanks JV, Morgan J. Plant ‘hairy root’ culture. Curr Opin Biotechnol. 1999;10(2):151–5.
Makhzoum AB, Sharma P, Bernards MA, Trémouillaux-Guiller J. Hairy roots: an ideal platform for transgenic plant production and other promising applications. In: Gang DR (ed) Phytochemicals, plant growth, and the environment. Springer; 2013. p. 95–142.
Lozovaya VV, Lygin AV, Zernova OV, Ulanov AV, Li SX, Hartman GL, et al. Modification of phenolic metabolism in soybean hairy roots through down regulation of chalcone synthase or isoflavone synthase. Planta. 2007;225(3):665–79.
Li M, Hu Z, Jiang QY, Sun XJ, Guo Y, Qi JC, et al. GmNAC15 overexpression in hairy roots enhances salt tolerance in soybean. J Integr Agric. 2017;16:60345–7.
Wang F, Chen HW, Li QT, Wei W, Li W, Zhang WK, et al. GmWRKY27 interacts with GmMYB174 to reduce expression of GmNAC29 for stress tolerance in soybean plants. Plant J. 2015;83(2):224–36.
Liu SM, Kandoth PK, Warren SD, Yeckel G, Heinz R, Alden J, et al. A soybean cyst nematode resistance gene points to a new mechanism of plant resistance to pathogens. Nature. 2012;492(7428):256–60.
Morriss SC, Studham ME, Tylka GL, MacIntosh GC. Validation of a hairy roots system to study soybean–soybean aphid interactions. PLoS ONE. 2017;12(3):e0174914.
Ma ZC, Zhu L, Song TQ, Wang Y, Zhang Q, Xia YQ, et al. A paralogous decoy protects Phytophthora sojae apoplastic effector PsXEG1 from a host inhibitor. Science. 2017;355(6326):710–4.
Subramanian S, Graham MY, Yu O, Graham TL. RNA interference of soybean isoflavone synthase genes leads to silencing in tissues distal to the transformation site and to enhanced susceptibility to Phytophthora sojae. Plant Physiol. 2005;137(4):1345–53.
Li WM, Pu ZQ. Studies on the comparison of three methods for purification of soybean mosaic virus. J Nanjing Agric Univ. 1990;13(4):56–9.
Voinnet O, Pinto YM, Baulcombe DC. Suppression of gene silencing: a general strategy used by diverse DNA and RNA viruses of plants. Proc Natl Acad Sci. 1999;96(24):14147–52.
Earley KW, Haag JR, Pontes O, Opper K, Juehne T, Song KM, et al. Gateway-compatible vectors for plant functional genomics and proteomics. Plant J. 2006;45(4):616–29.
Allen GC, Flores-Vergara MA, Krasynanski S, Kumar S, Thompson WF. A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide. Nat Protoc. 2006;1(5):2320–5.
Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72(1–2):248–54.
Voinnet O, Rivas S, Mestre P, Baulcombe D. An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. Plant J. 2003;33:949–56.
Wang X, Eggenberger AL, Nutter FW, Hill JH. Pathogen-derived transgenic resistance to soybean mosaic virus in soybean. Mol Breed. 2001;8(2):119–27.
Voinnet O. Post-transcriptional RNA silencing in plant–microbe interactions: a touch of robustness and versatility. Curr Opin Plant Biol. 2008;11(4):464–70.
Prins M, Laimer M, Noris E, Schubert J, Wassenegger M, Tepfer M. Strategies for antiviral resistance in transgenic plants. Mol Plant Pathol. 2008;9(1):73–83.
Asurmendi S, Berg RH, Smith TJ, Bendahmane M, Beachy RN. Aggregation of TMV CP plays a role in CP functions and in coat-protein-mediated resistance. Virology. 2007;366(1):98–106.
Dinant S, Kusiak C, Cailleteau B, Verrier J, Chupeau M, Chupeau Y, et al. Field resistance against potato virus Y infection using natural and genetically engineered resistance genes. Eur J Plant Pathol. 1998;104(4):377–82.
Jiang H, Li K, Dou DL, Gai JY. Characterization of a Soybean mosaic virus variant causing different diseases in Glycine max and Nicotiana benthamiana. Arch Virol. 2017;162(2):549–53.
Willman M. Transient expression efficiency of a novel Agrobacterium strain in soybean leaf tissue. Denman Undergraduate Research Forum. 2015.
Azhakanandam K, Su P. Methods and compositions for a soybean in-planta transient expression system: U. S. Patent 9,068,191 [P]. 2015-6-30.
Wu FQ, Hanzawa Y. A simple method for isolation of soybean protoplasts and application to transient gene expression analyses. J Vis Exp JoVE. 2018;131:e57258.
Zhi H, Gai J. Performances and germplasm evaluation of quantitative resistance to soybean mosaic virus in soybeans. Agric Sci China. 2004;3(4):247–53.
Bulgakov VP, Shkryl YN, Veremeichik GN, Gorpenchenko TY, Vereshchagina YV. Recent advances in the understanding of Agrobacterium rhizogenes—derived genes and their effects on stress resistance and plant metabolism. Adv Biochem Eng Biotechnol. 2013;134:1–22.
Acknowledgements
None.
Funding
This work was financially supported through the grants from the Natural Science Foundation of China (31801388), the Fundamental Research Funds for the Central Universities (KJQN201909), National Key R & D Program for Crop Breeding in China (2017YFD0101500), the MOE 111 Project (B08025), the MOE Program for Changjiang Scholars and Innovative Research Team in University (PCSIRT_17R55), the MOA CARS-04 program, the Jiangsu Higher Education PAPD Program and the Jiangsu JCIC-MCP.
Author information
Authors and Affiliations
Contributions
GJY conceived and designed the method and experiments. JH and LK performed the experiments and analyzed the data. GJY contributed reagents/materials and interpretation of the results. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors are consent for publication.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1: Fig. S1.
The gene arrangement in T-DNA constructs used in this study.
Additional file 2: Table S1
The primers are used to construct control and destination vectors.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Jiang, H., Li, K. & Gai, J. Agrobacterium rhizogenes-induced soybean hairy roots versus Soybean mosaic virus (ARISHR-SMV) is an efficient pathosystem for studying soybean–virus interactions. Plant Methods 15, 56 (2019). https://doi.org/10.1186/s13007-019-0442-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13007-019-0442-8