
Abstract
Background
Insecticide resistance threatens malaria control in sub-Saharan Africa. Knockdown resistance to pyrethroids and organochlorines in Anopheles gambiae sensu lato (s.l.) is commonly caused by mutations in the gene encoding a voltage-gated sodium channel which is the target site for the insecticide. The study aimed to examine risk factors for knockdown resistance in An. gambiae s.l. and its relationship with malaria infection in children in rural Gambia. Point mutations at the Vgsc-1014 locus, were measured in An. gambiae s.l. during a 2-year trial. Cross-sectional surveys were conducted at the end of the transmission season to measure malaria infection in children aged 6 months–14 years.
Results
Whilst few Anopheles arabiensis and Anopheles coluzzii had Vgsc-1014 mutations, the proportion of An. gambiae sensu stricto (s.s.) mosquitoes homozygous for the Vgsc-1014F mutation increased from 64.8 to 90.9% during the study. The Vgsc-1014S or 1014F mutation was 80% higher in 2011 compared to 2010, and 27% higher in the villages with indoor residual spraying compared to those without. An increase in the proportion of An. gambiae s.l. mosquitoes with homozygous Vgsc-1014F mutations and an increase in the proportion of An. gambiae s.s. in a cluster were each associated with increased childhood malaria infection. Homozygous Vgsc-1014F mutations were, however, most common in An. gambiae s.s. and almost reached saturation during the study meaning that the two variables were colinear.
Conclusions
As a result of colinearity between homozygous Vgsc-1014F mutations and An. gambiae s.s., it was not possible to determine whether insecticide resistance or species composition increased the risk of childhood malaria infection.
Similar content being viewed by others

Background
Between 2000 and 2015, the prevalence of Plasmodium falciparum infection in sub-Saharan Africa (SSA) has halved due to the mass deployment of long-lasting insecticidal nets (LLINs), and to a lesser extent, indoor residual spraying (IRS) [1]. This has, however, increased selection pressure for insecticide resistance in malaria vectors, particularly resistance to pyrethroids, the only insecticide class used currently for treating bed nets. The strength and distribution of insecticide resistance has increased over time and there is growing concern that this will lead to control failure, which has the potential to reverse many of the gains seen in malaria control [2]. One of several mechanisms through which mosquitoes become resistant to insecticides is through mutations in the insecticide target site. Three different point mutations in the voltage-gated sodium channel gene confer knockdown resistance (kdr) to pyrethroids and organochlorines such as dichlorodiphenyltrichloroethane (DDT) in Anopheles gambiae s.l. [3,4,5].
There has been little longitudinal insecticide resistance monitoring in The Gambia, but the general impression is that levels of insecticide resistance are low, but rising. Shortly after the introduction of permethrin-treated bed nets in The Gambia in the early 1990s, there was little or no resistance to DDT or permethrin [6, 7]. Prior to the nationwide DDT IRS campaign in 2009, DDT resistance was found in one site bordering Senegal, but mosquitoes from the site in the Upper River Region (URR) were fully susceptible to permethrin, deltamethrin and DDT [8]. Tests performed during a cluster-randomised controlled trial in the URR which compared the efficacy of LLINs versus LLINs and IRS with DDT against malaria in children found complete susceptibility of An. gambiae s.l. to DDT and permethrin in 2010 and some loss of susceptibility the following year [9]. Another study, conducted in the same year but outside the study area indicates that there may be pockets of high resistance in the URR [10]. Research also suggests that insecticide resistance may be partly responsible for the heterogeneities in malaria transmission across the country [11]. Although malaria has been declining in The Gambia since 2000 [12, 13], there is continued moderately high seasonal transmission in the URR despite high vector control coverage [14].
The study aimed to examine risk factors for kdr resistance in An. gambiae s.l. and its relationship with malaria infection in children in The Gambia.
Methods
Study site
The study was conducted in the URR (regional capital: Basse Santa Su, 13.3167°N, − 14.2167°W), a rural area of open Sudanian savannah which is divided into north and south banks by the River Gambia. Malaria transmission is highly seasonal being associated with the annual rains which occur from June to October. LLIN use by study children was 55% at baseline and IRS with DDT was implemented in 2009, the year prior to study start.
Data collection
This secondary analysis uses data from a cluster-randomised controlled trial assessing the efficacy of LLINs versus LLINs and IRS with DDT against malaria among children aged 6 months–14 years in the URR of The Gambia [9]. The study design and results are described in full elsewhere [9, 15]. In brief, 70 clusters of villages were randomly allocated to receive either LLINs or LLINs plus IRS. Permethrin-treated LLINs (2%; Olyset Nets, Sumitomo Chemicals, Japan) were distributed in both arms at the start of the 2010 transmission season to achieve high coverage of sleeping places. IRS with DDT (2 g/m2, DDT 75% wettable powder; Hindustan Insecticides, New Delhi, India) was applied to dwelling rooms at the start of each transmission season in the IRS-LLIN arm. Surveys conducted at the end of the transmission season in 2010 and 2011 measured the prevalence of P. falciparum infection in a cohort of children.
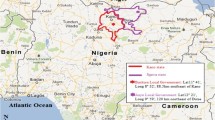
Entomological data were collected in 32 clusters (16 in each arm) in six sentinel rooms per cluster. Clusters were chosen purposively for logistical reasons (Fig. 1). Sampling was performed monthly from June to the end of December in 2010 and 2011, and every 2 months during the intervening dry season. Mosquitoes were collected overnight from sentinel rooms in which an adult slept under an LLIN using a CDC light trap. The epidemiological dataset was restricted to children who resided in the entomological clusters (1543 children in 2010 and 1564 children in 2011).

Spatial distribution of 32 entomological sampling sites in the Upper River Region of The Gambia, in relation to landcover/use
Mosquitoes were identified to species using established keys [16, 17]. Sub-species (Anopheles arabiensis, Anopheles coluzzii, An. gambiae s.s. and hybrid An. gambiae s.s. x An. coluzzii form, hereafter termed ‘hybrid’) and genotype at the Vgsc-1014 locus were determined according to previously described molecular methods [18,19,20]. Leucine to serine (Vgsc-1014S, previously termed kdr-east) or leucine to phenylalanine (Vgsc-1014F, previously termed kdr-west) mutations at this locus confer kdr.
Mapping and spatial analysis
Digitised maps produced by the Japan International Cooperation Agency under The Japanese Government Technical Cooperation Programme and The Government of the Republic of The Gambia from 2002 were obtained. Global Moran’s spatial autocorrelation coefficient, I, was calculated at 1 km intervals between 9 km (the shortest distance at which all sampling point locations had at least one neighbour) and 25 km to examine spatial independence in species distributions. The z-score returned indicated the intensity of clustering. Mapping and spatial analysis was performed using ArcGIS® software (Release 10.4.1, Environmental Systems Research Institute: Redlands, CA).
Statistical analysis
Transmission seasons were defined as 16 August–31 December 2010 and 15 August 2011–1 January 2012 to avoid the months prior to and during application of IRS and the intervening dry season. Proportions of mosquitoes by species and kdr status over time and by village were calculated. Mixed effect logistic regression models including cluster as a random effect were used to determine the relationship between vector species of individual mosquitoes and Euclidean distance of the cluster from the River Gambia, and secondly, the effect of variables such as year and study arm on kdr status of individual mosquitoes, whilst controlling for species. Variables were tested for departure from linear trend where necessary. Stepwise selection procedures and likelihood ratio tests were used to determine the combination of covariates, which fitted the data best. Mixed effect logistic regression models were also used to look at the effect of i) cluster-level kdr status (prevalence of any Vgsc-1014 mutation i.e. any mutation at the Vgsc-1014 locus, and homozygous Vgsc-1014F mutation by cluster) and ii) prevalence of An. gambiae s.s. by cluster on the odds of P. falciparum infection in individual children at the end of transmission season surveys, adjusting for clustering and confounding variables. These three explanatory variables were fit as linear variables and expressed as the odds ratio for the effect of a 1 and 10% increase in these variables on the prevalence of malaria infection. R2 and the variance inflation factor (VIF) were calculated to identify colinearity between variables. Goodness of fit of models evaluating the effect of cluster level prevalence of either An. gambiae s.s. or homozygous Vgsc-1014F mutations on malaria infection were compared using the Akaike information criterion (AIC). Models were also run to evaluate the effect of absolute numbers of mosquitoes per cluster with any Vgsc-1014 mutation and the homozygous Vgsc-1014F mutation, and absolute number of An. gambiae s.s. on the odds of P. falciparum infection in individual children. Statistical analyses were performed using Stata 14 (College Station, TX, USA).
Results
A total of 6853 An. gambiae s.l. were caught in the 32 sampling sites over the two transmission seasons. Of these, 6828 (99.6%) were identified to species: 71.3% were An. arabiensis, 15.0% An. gambiae s.s., 12.3% An. coluzzii, and 0.1% hybrid (Fig. 2). Higher numbers were caught during 2010 when there was unusually high rainfall and extensive flooding, compared to 2011 when flooding was limited to areas beside the river. During the 2010 transmission season, 76.1% of An. gambiae s.l. were An. arabiensis, 12.0% An. gambiae s.s., 10.1% An. coluzzii and 0.2% hybrid (Additional file 1). During the 2011 transmission season, 57.8% of An. gambiae s.l. caught were An. arabiensis, 23.0% An. gambiae s.s., 19.0% An. coluzzii and 0.1% hybrid. Twenty-nine mosquitoes were caught during the dry season (of these 25 were An. arabiensis, 2 An. gambiae s.s. and 2 An. coluzzii). Spatial autocorrelation was found in species distributions with peak autocorrelation operating between 9 and 14 km depending on the species and year.

Number of An. arabiensis, An. gambiae s.s., An. coluzzii and hybrid (An. gambiae s.s. × An. coluzzii) caught using CDC light traps per round during 2010 and 2011 (IRS using DDT was administered between 15–28 July 2010 and 20 July–9 August 2011)
Analysis of species distributions over the two transmission seasons, showed that An. gambiae s.s. was more common further away from the river (Odds ratio, OR for every km away from the river = 1.29, 95% CI 1.21–1.38, p < 0.001) (Figs. 3, 4). Conversely, both An. arabiensis and An. coluzzii were more common closer to the river (An. arabiensis OR = 0.88, 95% CI 0.83–0.94, p < 0.001; An. coluzzii OR = 0.91, 95% CI 0.85–0.98, p = 0.01). Similar patterns were found when each year was analysed separately.

Distribution of members of the An. gambiae s.l. complex in the study area during 2010 (a) and 2011 (b) transmission seasons. Pie charts show percentage composition of species of An. gambiae s.l. complex at CDC light trap sampling sites, (excluding sampling sites with less than 10 mosquitoes caught in total across each transmission season)

Distribution of members of the An. gambiae s.l. species complex (excluding An. arabiensis) in the study area during 2010 (a) and 2011 (b) transmission seasons. Pie charts show percentage An. gambiae s.l. species composition (excluding An. arabiensis) at CDC light trap sampling sites (excluding sampling sites with less than 10 mosquitoes caught in total across each transmission season)
In 2010, An. arabiensis comprised 81.8% of mosquitoes caught in the LLIN only arm (63.3% in 2011) and 67.8% in the IRS-LLIN arm (50.3% in 2011). As a result, there was a significantly lower odds of collecting An. arabiensis in the double intervention arm compared to the LLIN arm of the study in both years (2010: odds ratio, OR = 0.51, 95% CI 0.31–0.84, p = 0.008; 2011: OR = 0.44, 95% CI 0.25–0.78, p = 0.005). In 2010, An. gambiae s.s. comprised 6.5% of mosquitoes in the LLIN only arm (17.9% in 2011) and 19.8% in the IRS-LLIN arm (29.9% in 2011). There was a significantly higher odds of finding An. gambiae s.s. in the IRS-LLIN arm compared to the LLIN arm of the study in both years (2010: OR = 3.02, 95% CI 1.38–6.57, p = 0.005; 2011: OR = 2.71, 95% CI 1.18–6.19, p = 0.02). There was no difference in the odds of catching An. coluzzii between the two study arms in both 2010 and 2011 (2010: OR = 1.12, 95% CI 0.71–1.79, p = 0.63; 2011: OR = 0.81, 95% CI 0.41–1.60, p = 0.54), nor was there a difference in the odds of catching hybrids between the two study arms in 2010 (OR = 1.93, 95% CI 0.43–8.62, p = 0.39).
Vgsc-1014 mutations were found in all species sampled but at differing levels. An. arabiensis were predominantly wild-type (73.1% during 2010 and 58.1% during 2011), although the proportion with heterozygous Vgsc-1014S mutations increased from 20.5% in 2010 to 28.3% in 2011 (OR = 1.58, 95% CI 1.32–1.89, p < 0.001) (Table 1). Anopheles gambiae s.s. were predominantly homozygous Vgsc-1014F and this proportion increased almost to saturation from 64.8% in 2010 to 90.9% in 2011 (OR = 8.24, 95% CI 4.99–13.63, p < 0.001). An. coluzzii were predominantly wild-type (73.1% during 2010 and 70.9% during 2011).
In both years, the odds of having any type of Vgsc-1014 mutation was significantly higher in the IRS-LLIN arm compared to the LLIN only arm (2010: OR = 1.54, 95% CI 1.07–2.22, p = 0.02; 2011: OR = 2.26, 95% CI 1.24–4.11, p = 0.01) (Table 2). This was primarily due to the higher proportion of Vgsc-1014F mutations, particularly homozygous Vgsc-1014F mutations, in the double intervention compared to the single intervention arm. In 2010, IRS-LLIN villages had 2.24 times the odds of mosquitoes carrying homozygous Vgsc-1014F mutations compared to LLIN only villages (95% CI 1.12–4.49, p = 0.02), while in 2011, double intervention villages had 2.52 times the odds (95% CI 1.20–5.29, p = 0.01). There was an increased odds of a mosquito carrying the heterozygous Vgsc-1014F mutation in 2010 (OR = 2.17, 95% CI 1.28–3.68, p = 0.004), but not in 2011 (OR = 1.32, 95% CI 0.75–2.33, p = 0.34). No significant difference in the odds of heterozygous or homozygous Vgsc-1014S mutations was found between IRS-LLIN villages and LLIN villages in 2010 or 2011.
Distribution maps of Vgsc-1014 mutations show an increase in the proportion of mosquitoes carrying homozygous Vgsc-1014F mutations in villages on the south bank and in the northern part of the study area bordering Senegal between 2010 and 2011 (Fig. 5), which mirrors the increase in the proportion of An. gambiae s.s. in these areas (Fig. 3, 4).

Vgsc-1014 mutation status of An. gambiae s.l. in the study area during 2010 (a) and 2011 (b) transmission seasons. Pie charts show percentage wildtype, homozygous and heterozygous Vgsc-1014F and Vgsc-1014S mutations in An. gambiae s.l. complex at CDC light trap sampling sites (excluding sampling sites with less than 10 mosquitoes caught in total across each transmission season)
In a multivariable model, species, study arm and year of survey were associated with odds of any Vgsc-1014 mutation (Table 3). Adjusting for year and study arm, An. gambiae s.s. mosquitoes had 18.49 times the odds of having any Vgsc-1014 mutation compared to An. arabiensis (95% CI 14.48–23.61, p < 0.001), while An. coluzzii had 0.76 times the odds of having any Vgsc-1014 mutation (95% CI 0.63–0.92, p = 0.004) compared to An. arabiensis. Adjusting for species and year, mosquitoes caught in the LLIN-IRS arm had 1.27 times the odds of having any Vgsc-1014 mutation compared to mosquitoes in the LLIN only arm (95% CI 1.03–1.55, p = 0.02). Adjusting for species and study arm, mosquitoes caught in 2011 had 1.80 times the odds of having any Vgsc-1014 mutation compared to those caught in 2010 (95% CI 1.56–2.09, p < 0.001).
P. falciparum infection status was ascertained for 1543 children at the end of the 2010 transmission season and 1564 children in 2011. Multivariable analysis showed that girls were more likely to be infected with P. falciparum in 2010 (OR = 1.47, 95% CI 1.08–1.98, p = 0.01), but were less likely to be infected in 2011 (OR = 0.73, 95% CI 0.56–0.96, p = 0.02) (Table 4). Older children were more likely to be infected with P. falciparum at the end of the transmission season in both years (OR for 1 year increase in age: 2010 = 1.06, 95% CI 1.02–1.10, p = 0.007, 2011 = 1.12, 95% CI 1.08–1.17, p < 0.001). In 2011, children sleeping under an LLIN the previous night were less likely to be infected than children who had not slept under an LLIN (OR = 0.49, 95% CI 0.28–0.83, p = 0.009), although there was no significant association between LLIN use and infection in 2010. There was no significant association between cluster level prevalence of any Vgsc-1014F mutation and malaria infection in children in either year. In univariable analysis there was a tendency towards an increased odds of P. falciparum infection among children living in clusters with a high proportion of mosquitoes specifically carrying homozygous Vgsc-1014F mutations (OR for a 10% increase in the proportion of An. gambiae s.l. mosquitoes with any Vgsc-1014 mutation 2010: OR = 1.25, 95% CI 0.99–1.58, p = 0.07, 2011: OR = 1.13, 95% CI 1.01–1.27, p = 0.04). There was also a similar magnitude increase in the odds of P. falciparum infection among children living in clusters with a high proportion of mosquitoes identified as An. gambiae s.s. in univariable analysis (OR for a 10% increase in the proportion of An. gambiae s.l. mosquitoes identified as An. gambiae s.s. 2010: OR = 1.27, 95% CI 1.03–1.55, p = 0.02, 2011: OR = 1.12, 95% CI 1.01–1.25, p = 0.03). The proportion of homozygous Vgsc-1014F mutations and proportion of An. gambiae s.s. at each cluster were colinear (2010: R2 = 0.94, VIF = 17.3, 2011: R2 = 0.91, VIF = 10.8). As a result, these variables could not be combined in the multivariable model. Comparison of the AIC for models including either cluster level proportion of An. gambiae s.s. or mosquitoes carrying homozygous Vgsc-1014F mutations was not able to distinguish which model provided better goodness of fit (2010: AIC for model including An. gambiae s.s. = 1195.38, AIC for model including homozygous Vgsc-1014F mutations = 1196.90, 2011: AIC for model including An. gambiae s.s. = 1413.25, AIC for model including homozygous Vgsc-1014F mutations = 1413.71). Looking at total number of mosquitoes collected in each cluster, rather than percentage composition, there was no significant association between either the absolute number of An. gambiae s.s. mosquitoes, mosquitoes with Vgsc-1014 mutations or mosquitoes with homozygous Vgsc-101F mutations and malaria infection in children at the end of the transmission season in both years.
Discussion
These findings illustrate the temporal and spatial pattern of the An. gambiae complex and Vgsc-1014 mutations in the URR of The Gambia and their association with malaria infection in children from 2010 to 2011. To the authors knowledge, this is the first study to adopt a landscape approach with intensive entomological sampling to understand factors related to the distribution of kdr in malaria vectors.
As in previous work in the URR [21,22,23,24], An. arabiensis was the most abundant member of the An. gambiae complex and persisted longer into the dry season than the other species. The frequency of hybrids was 0.1–0.2%, a slightly lower proportion than that shown by others in The Gambia [24, 25]. Anopheles gambiae s.s. were more common in villages away from the River Gambia which corresponds with previous studies that reported An. gambiae s.s. prefers small rain-dependent larval habitats on free-draining soil covered with open woodland savannah or farmland [26, 27]. Anopheles arabiensis was more common near the river suggesting that their aquatic habitats are common in wetlands, such as rain-fed ricefields, adjacent to the river [28]. Indeed, previous studies found An. arabiensis in water bodies along the edge of the alluvial soils, particularly in areas of rice cultivation in the Central River Region [29]. Anopheles coluzzii was also more common closer to the river which supports research showing this species exploits semi-permanent aquatic habitats that are also frequented by An. arabiensis [26, 30, 31]. Although the literature suggests the distribution of species is likely to be due to differential larval habitats, this result may be because An. gambiae s.s. has less flexible host choice behaviours than An. arabiensis [32,33,34] and so contributes a larger proportion of the mosquito catch further away from the river.
Interestingly, fewer An. arabiensis were caught in villages in the double intervention compared to the single intervention arm, while the opposite pattern was seen with An. gambiae s.s. If the Vgsc-1014F mutation translates into phenotypic resistance, this may have given An. gambiae s.s. a competitive advantage over An. arabiensis in the double intervention arm. This is a different pattern from that in East Africa where, with the scale-up of interventions, An. arabiensis is starting to dominate over An. gambiae s.s. since the latter is more endophagic and endophillic and so is thought to be preferentially killed by LLINs [35,36,37].
Species, year of survey and study arm were associated with odds of any Vgsc-1014 mutation. There was a 1.27 increase in the odds of any Vgsc-1014 mutation in the double intervention compared to the single intervention arm and a 1.80 increase in the odds of any Vgsc-1014 mutation between 2010 and 2011. The odds of any Vgsc-1014 mutation was 18.49 times higher in An. gambiae s.s. compared to An. arabiensis. Taken together this suggests that (i) LLINs and DDT used together provide more selection pressure than LLINs alone and (ii) there was an increase in selection pressure over the two years most likely due to the second IRS round, and (iii) selection pressure favours An. gambiae s.s. since it has a higher frequency of kdr. Several studies have shown an increase in the frequency of kdr mutations following implementation of vector control interventions [38,39,40,41]. High levels of kdr in An. gambiae s.s. compared to An. arabiensis may be explained by a greater propensity for indoor resting and feeding of An. gambiae s.s. and, therefore, potential for increased contact with insecticides on walls or LLINs [17, 34, 42]. The study used DDT for IRS and pyrethroid-treated LLINs, and as such is it unsurprising that selection pressure for development of Vgsc-1014 mutations was high in the double intervention arm. The IRS in the study was performed by government teams using the insecticide selected by the National Malaria Control Programme but an alternative insecticide class, such as an organophosphate or carbamate would have been a better option to reduce selection pressure [10]. In fact, based on insecticide resistance monitoring, the control programme recently started to implement rotation of IRS insecticides beginning with bendiocarb in 2015 and 2016 and pirimiphos-methyl in 2017.
Older children had a higher odds of P. falciparum infection in both of the end of season surveys. It is unclear why females were at increased risk of infection in the first survey but at lower risk in the second survey and this may be an anomalous result. In 2011, children sleeping under an LLIN had half the odds of being infected compared to children not sleeping under a net, but no significant difference was observed in 2010. This result is probably due to chance because of low numbers of children not using an LLIN in this study. There was no significant association between the cluster level proportion of mosquitoes with any Vgsc-1014F mutations and malaria infection in children. However, univariable analysis did show an association between P. falciparum infection in children and the cluster level proportion of An. gambiae s.s. and homozygous Vgsc-1014F mutations specifically, especially in the second year of the study. However, due to colinearity between An. gambiae s.s. and homozygous Vgsc-1014F mutations these variables could not be combined in the multivariable model. Calculation of the AIC was not able to distinguish between models including these two variables and so it was not possible to say whether high proportions of An. gambiae s.s. or homozygous Vgsc-1014F mutations increased P. falciparum infection in children.
This study has several other limitations. Firstly, the analysis used secondary data which meant that the original study was not primarily designed to measure the spatial epidemiology of kdr resistance. Secondly, it did not verify the phenotype of the mosquitoes in bioassays or investigate other resistance markers, such as those involved in metabolic resistance. Insecticide resistance is typically driven by complex interactions between multiple alleles and this dataset only looks at a few alleles. Thirdly, spatial autocorrelation was present in species distributions and therefore the assumption that clusters were independent was false. This may have inflated the value of test statistics and increased the chance of a type I error.
Increased malaria infection in the study children may be explained by differences in the species distribution in the villages, specifically possible higher efficiency transmission by An. gambiae s.s. Anopheles gambiae s.s. is a more efficient vector than An. arabiensis, although it is less clear whether there is a difference in transmission efficiency between An. gambiae s.s. and An. coluzzii [43,44,45]. Alternatively, heterogeneity in malaria infection could also be due to the impact of kdr. Indeed, previous studies have highlighted a lack of decline in malaria in the URR [14, 46] and a study of paired high and low malaria prevalence villages in The Gambia suggested that heterogeneous transmission may be partly due to insecticide resistance [11]. Opondo et al. showed that DDT mortality for An. gambiae s.s. was significantly lower in high prevalence compared to low prevalence villages and that there was a significant association between the Vgsc-1014F mutation in An. gambiae s.s. and resistance to DDT and deltamethrin [11]. This mutation was a strong predictor of insecticide resistance and effectively masked the effect of other mutations in this study such as those associated with metabolic resistance. However, the role of kdr is not clear cut [47, 48] and several studies show that pyrethroid LLINs were still able to kill An. gambiae despite high kdr frequencies [49,50,51,52]. Analysis did not show any significant relationship between childhood malaria infection and the absolute number of An. gambiae s.s. mosquitoes, mosquitoes with any Vgsc-1014 mutation or mosquitoes with the homozygous Vgsc-1014F mutation per cluster. This is most likely because of low vector numbers in some of the village clusters.
Conclusions
In conclusion, the homozygous Vgsc-1014F mutation occurred predominantly in An. gambiae s.s. and increased almost to saturation during the course of the study. It also occurred at higher frequencies where IRS was used in addition to LLINs, probably because the kdr mutation confers a selective advantage in the presence of insecticides. There was a 13% increase in the odds of malaria infection in children associated with a 10% increase in the proportion of An. gambiae s.l. carrying the Vgsc-1014F mutation in 2011. Moreover there was a 27% increase in the odds of malaria infection with a 10% increase in the proportion of An. gambiae s.s. mosquitoes in 2010 and a 12% increase in 2011. It was, however, impossible to determine whether resistance or species increased the odds of childhood malaria infection since the homozygous Vgsc-1014F mutation was colinear with An. gambiae s.s.
Change history
07 June 2018
Unfortunately, the original article [1] contained an error mistakenly carried forward by the Production department handling this article whereby some figures and their captions were interchanged. The correct figures (Figs. 1, 2, 3, 4, 5) and captions are presented in this erratum. The original article has also been updated to reflect this correction.
Abbreviations
- AIC:
-
akaike information criterion
- CDC:
-
Centers for Disease Control
- CI:
-
confidence interval
- DDT:
-
dichlorodiphenyltrichloroethane
- IRS:
-
indoor residual spraying
- kdr :
-
knockdown resistance
- LLIN:
-
long-lasting insecticidal net
- OR:
-
odds ratio
- SSA:
-
sub-Saharan Africa
- URR:
-
Upper River Region
- VIF:
-
variance inflation factor
References
Bhatt S, Weiss DJ, Cameron E, Bisanzio D, Mappin B, Dalrymple U, et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature. 2015;526:207–11.
Ranson H, Lissenden N. Insecticide resistance in African Anopheles mosquitoes: a worsening situation that needs urgent action to maintain malaria control. Trends Parasitol. 2016;32:187–96.
Martinez-Torres D, Chandre F, Williamson MS, Darriet F, Berge JB, Devonshire AL, et al. Molecular characterization of pyrethroid knock-down resistance (kdr) in the major malaria vector Anopheles gambiae s.s. Insect Mol Biol. 1998;7:179–84.
Ranson H, Jensen B, Vulule JM, Wang X, Hemingway J, Collins FH. Identification of a point mutation in the voltage-gated sodium channel gene of Kenyan Anopheles gambiae associated with resistance to DDT and pyrethroids. Insect Mol Biol. 2000;9:491–7.
Jones CM, Liyanapathirana M, Agossa FR, Weetman D, Ranson H, Donnelly MJ, et al. Footprints of positive selection associated with a mutation (N1575Y) in the voltage-gated sodium channel of Anopheles gambiae. Proc Natl Acad Sci USA. 2012;109:6614–9.
Hemingway J, Lindsay SW, Small GJ, Jawara M, Collins FH. Insecticide susceptibility status in individual species of the Anopheles gambiae complex (Diptera: Culicidae) in an area of The Gambia where pyrethroid impregnated bednets are used extensively for malaria control. Bull Entomol Res. 1995;85:229–34.
Lindsay SW, Alonso PL, Armstrong Schellenberg JRM, Hemingway J, Thomas PJ, Shenton FC, et al. A malaria control trial using insecticide-treated bed nets and targeted chemoprophylaxis in a rural area of The Gambia, West Africa 3. Entomological characteristics of the study area. Trans R Soc Trop Med Hyg. 1993;87:19–23.
Betson M, Jawara M, Awolola TS. Status of insecticide susceptibility in Anopheles gambiae s.l. from malaria surveillance sites in The Gambia. Malar J. 2009;8:1–8.
Pinder M, Jawara M, Jarju LBS, Salami K, Jeffries D, Bojang K, et al. Efficacy of indoor residual spraying with dichlorodiphenyltrichloroethane against malaria in Gambian communities with high usage of long-lasting insecticidal mosquito nets: a cluster-randomised controlled trial. Lancet. 2014. https://doi.org/10.1016/S0140-6736(14)61007-2.
Tangena JA, Adiamoh M, D’Alessandro U, Jarju L, Jawara M, Jeffries D, et al. Alternative treatments for indoor residual spraying for malaria control in a village with pyrethroid- and DDT-resistant vectors in The Gambia. PLoS ONE. 2013;8:e74351.
Opondo KO, Weetman D, Jawara M, Diatta M, Fofana A, Crombe F, et al. Does insecticide resistance contribute to heterogeneities in malaria transmission in The Gambia? Malar J. 2016;15:166.
Ceesay SJ, Casals-Pascual C, Erskine J, Anya SE, Duah NO, Fulford AJ, et al. Changes in malaria indices between 1999 and 2007 in The Gambia: a retrospective analysis. Lancet. 2008;372:1545–54.
Ceesay SJ, Casals-Pascual C, Nwakanma DC, Walther M, Gomez-Escobar N, Fulford AJ, et al. Continued decline of malaria in The Gambia with implications for elimination. PLoS ONE. 2010;5:e12242. https://doi.org/10.1371/journal.pone.0012242.
Mwesigwa J, Okebe J, Affara M, Di Tanna GL, Nwakanma D, Janha O, et al. On-going malaria transmission in The Gambia despite high coverage of control interventions: a nationwide cross-sectional survey. Malar J. 2015;14:314.
Pinder M, Jawara M, Jarju LB, Kandeh B, Jeffries D, Lluberas MF, et al. To assess whether indoor residual spraying can provide additional protection against clinical malaria over current best practice of long-lasting insecticidal mosquito nets in The Gambia: study protocol for a two-armed cluster-randomised study. Trials. 2011;12:e147.
Gillies MT, DeMeillon B. The Anophelinae of Africa south of the Sahara (Ethiopian zoogeographical region). Johannesburg: The South African Institute for Medical Research; 1968.
Gillies MT, Coetzee M. A supplement to the Anophelinae of Africa south of the Sahara. Johannessburg: The South African Institute for Medical Research; 1987.
Bass C, Nikou D, Donnelly MJ, Williamson MS, Ranson H, Ball A, et al. Detection of knockdown resistance kdr mutations in Anopheles gambiae: a comparison of two new high-throughput assays with existing methods. Malar J. 2007;6:111.
Scott JA, Brogdon W, Collins FH. Identification of single specimens of the Anopheles gambiae complex by the polymerase chain reaction. Am J Trop Med Hyg. 1993;4:520–9.
Fanello C, Santolamazza F, Della Torre A. Simultaneous identification of species and molecular forms of the Anopheles gambiae complex by PCR-RFLP. Med Vet Entomol. 2002;16:461–4.
Oyewole IO, Awolola TS, Ibidapo CA, Oduola AO, Okwa OO, Obansa JA. Behaviour and population dynamics of the major anopheline vectors in a malaria endemic area in southern Nigeria. J Vector Borne Dis. 2007;44:56–64.
Lindsay SW, Wilkins HA, Zieler HA, Daly RJ, Petrarca V, Byass P. Ability of Anopheles gambiae mosquitoes to transmit malaria during the dry and wet seasons in an area of irrigated rice cultivation in The Gambia. J Trop Med Hyg. 1991;94:313–24.
Lindsay SW, Parson L, Thomas CJ. Mapping the ranges and relative abundance of the two principal African malaria vectors, Anopheles gambiae sensu stricto and An. arabiensis, using climate data. Proc Biol Sci. 1998;265:847–54.
Caputo B, Nwakanma D, Jawara M, Adiamoh M, Dia I, Konate L, et al. Anopheles gambiae complex along The Gambia river, with particular reference to the molecular forms of An. gambiae s.s. Malar J. 2008;7:182.
Nwakanma DC, Neafsey DE, Jawara M, Adiamoh M, Lund E, Rodrigues A, et al. Breakdown in the process of incipient speciation in Anopheles gambiae. Genetics. 2013;193:1221–31.
Della Torre A, Costantini C, Besansky NJ, Caccone A, Petrarca V, Powell JR, et al. Speciation within Anopheles gambiae—the glass is half full. Science. 2002;298:115–7.
Gimonneau G, Pombi M, Choisy M, Morand S, Dabiré RK, Simard F. Larval habitat segregation between the molecular forms of the mosquito Anopheles gambiae in a rice field area of Burkina Faso, West Africa. Med Vet Entomol. 2012;26:9–17.
Thomas CJ, Lindsay SW. Local-scale variation in malaria infection amongst rural Gambian children estimated by satellite remote sensing. Trans R Soc Trop Med Hyg. 2000;94:159–63.
Bøgh C, Clarke SE, Jawara M, Thomas CJ, Lindsay SW. Localized breeding of the Anopheles gambiae complex (Diptera: Culicidae) along the River Gambia, West Africa. Bull Entomol Res. 2003;93:279–87.
Costantini C, Ayala D, Guelbeogo WM, Pombi M, Some CY, Bassole IH, et al. Living at the edge: biogeographic patterns of habitat segregation conform to speciation by niche expansion in Anopheles gambiae. BMC Ecol. 2009;9:16.
Simard F, Ayala D, Kamdem GC, Pombi M, Etouna J, Ose K, et al. Ecological niche partitioning between Anopheles gambiae molecular forms in Cameroon: the ecological side of speciation. BMC Ecol. 2009;9:17.
White GB. Anopheles gambiae complex and disease transmission in Africa. Trans R Soc Trop Med Hyg. 1974;68:278–301.
Bryan JH. Anopheles gambiae and A. melas at Brefet, The Gambia, and their role in malaria transmission. Ann Trop Med Parasitol. 1983;77:1–12.
Sinka ME, Bangs MJ, Manguin S, Coetzee M, Mbogo CM, Hemingway J, et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: occurrence data, distribution maps and bionomic précis. Parasit Vectors. 2010;3:117.
Bayoh MN, Mathias DK, Odiere MR, Mutuku FM, Kamau L, Gimnig JE, et al. Anopheles gambiae: historical population decline associated with regional distribution of insecticide-treated bed nets in western Nyanza Province, Kenya. Malar J. 2010;9:62.
Mwangangi JM, Mbogo CM, Orindi BO, Muturi EJ, Midega JT, Nzovu J, et al. Shifts in malaria vector species composition and transmission dynamics along the Kenyan coast over the past 20 years. Malar J. 2013;12:13.
Russell TL, Lwetoijera DW, Maliti D, Chipwaza B, Kihonda J, Charlwood JD, et al. Impact of promoting longer-lasting insecticide treatment of bed nets upon malaria transmission in a rural Tanzanian setting with pre-existing high coverage of untreated nets. Malar J. 2010;9:1–14.
Aïzoun N, Aïkpon R, Akogbéto M. Evidence of increasing L1014F kdr mutation frequency in Anopheles gambiae s.l. pyrethroid resistant following a nationwide distribution of LLINs by the Beninese National Malaria Control Programme. Asian Pac. J Trop Biomed. 2014;4:239–43.
Padonou GG, Sezonlin M, Ossé R, Aizoun N, Oké-Agbo F, Oussou O, et al. Impact of three years of large scale Indoor Residual Spraying (IRS) and Insecticide Treated Nets (ITNs) interventions on insecticide resistance in Anopheles gambiae s.l. in Benin. Parasit Vectors. 2012;5:72.
Czeher C, Labbo R, Arzika I, Duchemin J. Evidence of increasing Leu-Phe knockdown resistance mutation in Anopheles gambiae from Niger following a nationwide long-lasting insecticide-treated nets implementation. Malar J. 2008;7:189.
Mathias DK, Ochomo E, Atieli F, Ombok M, Bayoh MN, Olang G, et al. Spatial and temporal variation in the kdr allele L1014S in Anopheles gambiae s.s. and phenotypic variability in susceptibility to insecticides in Western Kenya. Malar J. 2011;10:10.
White GB. The Anopheles gambiae complex and malaria transmission around Kisumu, Kenya. Trans R Soc Trop Med Hyg. 1972;66:572–81.
Taylor KA, Koros JK, Nduati J, Copeland RS, Collins FH, Brandling-Bennett AD. Plasmodium falciparum infection rates in Anopheles gambiae, An. arabiensis, and An. funestus in western Kenya. Am J Trop Med Hyg. 1990;43:124–9.
Ndiath MO, Cohuet A, Gaye A, Konate L, Mazenot C, Faye O, et al. Comparative susceptibility to Plasmodium falciparum of the molecular forms M and S of Anopheles gambiae and Anopheles arabiensis. Malar J. 2011;10:269.
Boissière A, Gimonneau G, Tchioffo MT, Abate L, Bayibeki A, Awono-Ambéné PH, et al. Application of a qPCR assay in the investigation of susceptibility to malaria infection of the M and S molecular forms of An. gambiae s.s. in Cameroon. PLoS ONE. 2013;8:e54820.
Okebe J, Affara M, Correa S, Muhammad AK, Nwakanma D, Drakeley C, et al. School-based countrywide seroprevalence survey reveals spatial heterogeneity in malaria transmission in The Gambia. PLoS ONE. 2014;9:e110926.
Brooke BD. kdr: can a single mutation produce an entire insecticide resistance phenotype? Trans R Soc Trop Med Hyg. 2008;102:524–5.
Donnelly MJ, Corbel V, Weetman D, Wilding CS, Williamson MS, Black WC 4th. Does kdr genotype predict insecticide-resistance phenotype in mosquitoes? Trends Parasitol. 2009;25:213–9.
Darriet F, N’Guessan R, Koffi AA, Konan L, Doannio JM, Chandre F, et al. Impact of pyrethrin resistance on the efficacy of impregnated mosquito nets in the prevention of malaria: results of tests in experimental cases with deltamethrin SC. Bull Soc Pathol Exot. 2000;93:131–4.
Asidi AN, N’Guessan R, Hutchinson RA, Traore-Lamizana M, Carnevale P, Curtis CF. Experimental hut comparisons of nets treated with carbamate or pyrethroid insecticides, washed or unwashed, against pyrethroid-resistant mosquitoes. Med Vet Entomol. 2004;18:134–40.
Asidi AN, N’Guessan R, Koffi AA, Curtis CF, Hougard JM, Chandre F, et al. Experimental hut evaluation of bednets treated with an organophosphate (chlorpyrifos-methyl) or a pyrethroid (lambdacyhalothrin) alone and in combination against insecticide-resistant Anopheles gambiae and Culex quinquefasciatus mosquitoes. Malar J. 2005;4:25.
Henry MC, Assi SB, Rogier C, Dossou-Yovo J, Chandre F, Guillet P, et al. Protective efficacy of lambdacyhalothrin treated nets in Anopheles gambiae pyrethroid resistance areas of Côte d’Ivoire. Am J Trop Med Hyg. 2005;75:859–64.
Authors’ contributions
Conceived of the study/analysis: ALW, MP, MJ, SWL. Conducted field and laboratory work: MP, MH-A, MJ, EJR, KS, LBSJ, BK. Conducted data analysis: ALW, MP, JB, DJ. Contributed to and approved the final manuscript: ALW, MP, JB, MJD, MH-A, LBSJ, MJ, DJ, BK, EJR, KS, UD, SWL. All authors read and approved the final manuscript.
Acknowledgements
We are grateful for the support of the villagers, the village health workers, the regional health team, and staff in the health clinics during conduct of the clinical trial. We also thank the Medical Research Council Unit in The Gambia for their technical and logistical support, especially the field, laboratory and data management staff. We also thank members of the Trial Steering Committee and the Data Safety and Monitoring Board.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets analysed during the current study are available from the Principal Investigator (S.W. Lindsay) on reasonable request.
Consent for publication
Not applicable.
Ethics approval and consent to participate
The trial was approved by the Gambian Government and Medical Research Council Unit Joint Ethics Committee and the London School of Hygiene & Tropical Medicine Ethics Committee. Prior to study start, meetings were held with village heads to explain about the study. Caregivers provided informed consent for collection of epidemiological data from children and householders provided informed consent for entomological data collection.
Funding
The study was funded by the UK Medical Research Council (Grant Number MRC GO900220). The funding body had no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Additional information
The original version of this article was revised; for further details, please see the Correction article https://doi.org/10.1186/s12936-018-2374-6.
Additional file
Additional file 1.
Characteristics of village clusters and proportion species composition during 2010 and 2011 transmission seasons.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Wilson, A.L., Pinder, M., Bradley, J. et al. Emergence of knock-down resistance in the Anopheles gambiae complex in the Upper River Region, The Gambia, and its relationship with malaria infection in children. Malar J 17, 205 (2018). https://doi.org/10.1186/s12936-018-2348-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-018-2348-8