
Abstract
Background
Clonal plants are important in maintaining wetland ecosystems. The main growth types of clonal plants are the guerrilla and phalanx types. However, little is known about the effects of these different clonal growth types on plant plasticity in response to heterogeneous resource distribution. We compared the growth performance of clonal wetland plants exhibiting the two growth forms (guerrilla growth form: Scirpus yagara, Typha orientalis, Phragmites australis and Sparganium stoloniferum; phalanx growth form: Acorus calamus, Schoenoplectus tabernaemontani and Butomus umbellatus) grown in soil substrates that were either homogeneous or heterogeneous but had the same total amount of nutrients.
Results
We found that the morphological traits (plant height, ramet number, spacer diameter and length) and biomass accumulation of the guerrilla clonal plants (T. orientalis) were significantly enhanced by heterogeneity, but those of the phalanx clonal plants (A. calamus, S. tabernaemontani and B. umbellatus) were not. The results showed that the benefits of environmental heterogeneity to clonal plants may be correlated with the type of clonal structure.
Conclusions
Guerrilla clonal plants, which have a dispersed, flexible linear structure, are better suited to habitats with heterogeneous resources. Phalanx clonal plants, which form compact structures, are better suited to habitats with homogeneous resources. Thus, wetland clonal species with the guerrilla clonal structure benefit more from soil nutrient heterogeneity.
Similar content being viewed by others
Background
Wetlands are unique areas that have characteristics of both land and water ecosystems and are among the world’s most productive environments [30]. Clonal growth forms dominate in many major biomes worldwide and are successful in wetland plant communities; thus, clonal plants play important roles in maintaining wetland ecosystems [25, 29, 31]. For example, clonal plants cover 66.69% of wetlands in China [31]. Emergent macrophytes have been shown to play an important role in wetland ecosystems [9, 37]. In aquatic habitats, vegetative propagation predominates among plant taxa; for instance, the majority of wetland species are rhizomatous clonal plants [32]. Clonal plants have special clonal life-history traits: I. trade-offs between clonal growth and reproduction [14]; II. clonal growth forms [34]; III. clonal plasticity [35]; and IV. clonal integration [2, 46]. For example, clonal plants can share resources (nutrients, water, etc.) among individual units through clonal integration, which increases plant survival and growth performance in habitats with different patterns of resource availability. In addition, clonal plants can increase their viability through risk sharing and resource storage [15, 44]. Thus, clonal plants have a strong ability to adapt to environmental pressure and resist disturbance.
Plants in natural habitats often experience heterogeneity in the spatial and temporal distribution of soil nutrients [21, 46]. Wetland plants, especially emergent macrophytes, are sensitive to the distribution of soil pollutants and nutrients during their growth process [7, 45]. In addition, clonal plants often exhibit more sensitive reactions than non-clonal plants; for example, the ramets of clonal plants interconnected by spacers (rhizomes, stolons, etc.) are often located in high-quality patches to efficiently utilize heterogeneously distributed resources [1, 12]. Spatial heterogeneity in soil nutrient availability can affect the growth performance of individual plants and the productivity and structure of plant communities [23, 36, 45]. For example, changing the spatial scale of nutrient heterogeneity can change the relative richness of species grown in mixtures [33, 43]. Thus, the heterogeneous distribution of soil nutrients may affect the fragile ecological stability of wetlands. Investigating the relationship between heterogeneous soil nutrient distribution and clonal wetland plants is highly important for shedding more light on the mechanisms of vegetation restoration.
Clonal plants with different clonal structure types may exhibit different strategies for adapting to their habitats and have different capacities for horizontal spreading [34, 42]. The main types of clonal plant growth are the guerrilla and phalanx types, but there are also many intermediate types [31, 34]. Clonal wetland plants that exhibit the phalanx structure, which consists of highly aggregated ramets connected by few and/or short spacers [24, 39]. In contrast, clonal wetland plants that exhibit the guerrilla growth form have a flexible distribution of ramets connected by many and/or long spacers [24, 39]. Phalanx clonal plants thrive in stable and homogeneous habitats, while guerrilla clonal plants grow in disturbed and heterogeneous habitats [41, 28, 40]. For example, soil nutrient heterogeneity significantly increased the relative yield of the guerrilla clonal plant Bolboschoenus planiculmis and decreased that of the phalanx clonal plant Carex neurocarpa [39]. However, few studies have focused on how clonal growth forms affect the responses of wetland plants to resource heterogeneity.
We designed an experiment to address the growth performance of clonal wetland plants in an environment of soil resource heterogeneity. The following questions were addressed: (1) Does soil nutrient heterogeneity have significant effects on the growth performance of clonal wetland plants? and (2) how do the growth responses of plants with different clonal growth forms to heterogeneous soil nutrients differ?
Materials and methods
Study area
The present study was conducted in Arongqi County, Inner Mongolia, China (48°10.883′ N, 123° 22.699′ E; altitude: 206 m). The Alun River in Arongqi County is a perennial flowing water body. The average annual temperature is 16.9 °C, the average annual precipitation is 470–570 mm, the average annual evaporation is 1400–1600 mm, and the average annual sunshine duration is 2600–2700 h in region.
Plant materials
Scirpus yagara, Typha orientalis, Phragmites australis, Sparganium stoloniferum, Acorus calamus, Schoenoplectus tabernaemontani and Butomus umbellatus are emergent-rooted wetland plants. These seven species, which commonly co-occur in many freshwater ecosystems and are the dominant species in various wetland habitats, were selected for this study. These plants have obvious clonal growth structures and can therefore be used to effectively compare the response mechanisms of the two clonal structures to heterogeneity.
Guerrilla clonal plants
Scirpus yagara, Typha orientalis, Phragmites australis, Sparganium stoloniferum are perennial, rhizomatous, herbaceous clonal plants that show the guerrilla growth form (Fig. 1). These plants can produce isolated ramets via long spacers, resulting in widely spaced ramets called “spreading ramets” [6, 24, 41].

Plant Materials. Guerrilla clonal plants: a Typha orientalis; b Phragmites australis; c Sparganium stoloniferum; d Scirpus yagara. Phalanx clonal plants: e Acorus calamus; f Schoenoplectus tabernaemontani; g Butomus umbellatus
Phalanx clonal plants
Acorus calamus, Schoenoplectus tabernaemontani and Butomus umbellatus are perennial, rhizomatous, herbaceous clonal plants that show the phalanx growth form (Fig. 1). These plants grow few short spacers, resulting in closely packed ramets called “clumping ramets” [4, 6, 20].
Experimental design
On April 12, 2016, ramets from each of the seven species were collected from the riparian zone of the Alun River. All of the collected ramets were precultivated in plastic buckets (70 cm long × 50 cm wide x 47 cm deep) with 20 cm of Alun River sediment (soil: mean ± SE, 0.29 ± 0.03 mg.g−1 N; 0.53 ± 0.02 mg.g−1 P; 31.64 ± 1.12 mg.g−1 organic material content) and 5 cm of Alun River water (water: 0.86 ± 0.14 mg.L−1N; 0.16 ± 0.04 mg.L−1 P) for approximately 60 days in the greenhouse. After culturing, 26 morphologically identical rooted ramets from each species (height: approximately 30 cm for S. tabernaemontani, T. orientalis, P. australis and A. calamus; approximately 20 cm for S. stoloniferum, B. yagara, and B. umbellatus) were selected. Ten ramets of each species were randomly selected to measure their initial dry biomass.
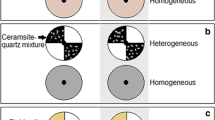
This experiment was set up in buckets (70 cm diameter x 70 cm height) with two substrate types: I. the heterogeneous soil treatment (HE), in which the buckets were divided into two areas (Fig. 2); one area was filled with river clay (soil: mean ± SE, 0.32 ± 0.02 mg.g−1 N; 0.55 ± 0.02 mg.g−1 P; 32.57 ± 1.21 mg.g−1 organic material content), and the other area was filled with pure sand; II. The homogeneous soil treatment (HO) was a mixture of the same total amount of river clay and sand per bucket. The total amount of soil nutrients was the same in all treatments. On July 2, 2016, 8 ramets of each species were planted at the intersection of the clay and sand areas of the heterogeneous buckets to ensure equal access to high-nutrient and low-nutrient patches. The remaining 8 ramets of each species were planted at the centre of the homogeneous soil buckets. One plant of each species was planted in each bucket. Each treatment was replicated eight times, and 112 buckets were used in total. Each treatment was watered with purified water every 2–3d to minimize the limitation of their growth due to water availability. All plant materials were harvested on September 22, 2016, the total number of ramets per plant was recorded, and the plant height, spacer length and diameter were measured. Each plant was then divided into its aboveground (leaves and stems above the soil surface) and belowground parts (roots and rhizomes), dried at 70 °C for 5d and weighed. The belowground/aboveground biomass ratio was calculated as follows:

Schematic representation of soil substrate types. The light grey area in He was filled with lake sediment, and the dotted area was filled with sand. The shaded area in Ho represents an even mixture of the same amount of lake sediment and sand
Statistical analysis
When necessary, the data were transformed and normalized. Thus, all data on plant traits met the assumptions of normality and homogeneity of variance prior to analysis. One-way ANOVA was applied to test the effects of the soil treatment on plant traits. Growth traits were analysed using a three-way nested ANOVA with soil nutrient heterogeneity (homogeneous vs. heterogeneous), growth form (phalanx vs. guerrilla), and species nested within growth forms as factors. All data analyses were performed using SPSS 22.0 (SPSS, Chicago, IL, United States).
Results
Soil nutrient heterogeneity had significant effects on the ramet number and belowground/aboveground biomass ratio, while clonal growth form did not significantly affect aboveground biomass or spacer length (Table 1). Soil nutrient heterogeneity x growth form had significant effects on the biomass and morphological characteristics, except spacer diameter, of the seven species (Table 1).
Guerrilla clonal plants
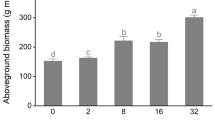
The heterogeneous soil treatment had a positive impact on the growth performance of guerrilla clonal plants. For example, the heterogeneous soil treatment significantly increased the biomass and morphological characteristics, except the belowground/aboveground biomass ratio, of T. orientalis (Figs. 3 and 4). Significantly higher biomass and spacer length were observed in P. australis in the heterogeneous soil treatment (Figs. 3 and 4). Additionally, the heterogeneous soil treatment significantly affected biomass accumulation and allocation, and increased some morphological traits (plant height, ramet number, spacer length) in guerrilla clonal plants. However, for S. yagara, a significantly higher ramet number and spacer diameter were observed in the heterogeneous soil treatment (Fig. 4).

Effects of substrate heterogeneity on biomass accumulation and distribution in seven clonal plants. Values are a aboveground biomass, b belowground biomass, c total biomass, and d belowground/aboveground biomass ratio. Values are means ± SEs. The bars with different lowercase letters are significantly different

Effects of substrate heterogeneity on the morphological traits of seven clonal plants. Values are a plant height, b ramet number, c spacer diameter, and d spacer length. Values are means ± SEs. The bars with different lowercase letters are significantly different
Phalanx clonal plants
The heterogeneous soil treatment had a negative impact on the growth performance of the phalanx clonal plants. For example, significantly lower biomass of A. calamus was observed in the heterogeneous soil treatment, but no effect was observed on its morphological characteristics (Figs. 3 and 4). The heterogeneous soil treatment significantly increased the belowground biomass, total biomass, belowground/aboveground biomass ratio and plant height of S. tabernaemontani (Figs. 3 and 4). The heterogeneous soil treatment significantly decreased the growth traits, except the spacer diameter, of B. umbellatus (Figs. 3 and 4).
Discussion
A few experimental results have shown that resource heterogeneity can increase plant performance, as measured by the accumulation and allocation of biomass. This has been observed at the levels of individual plants, populations and whole communities [3, 16 and 22]. However, other experiments have demonstrated that the positive responses of clonal plants to resource heterogeneity may not always be adaptive or may be temporary [10, 17, 27 and 45]. In this study, the performance of clonal wetland plants in heterogeneous environments was correlated with the type of clonal growth forms. For example, the heterogeneous distribution of nutrients in the soil substrate significantly increased the growth performance of the guerrilla clonal plants (T. orientalis; Figs. 3 and 4), but the homogeneous distribution of nutrients in the soil substrate significantly increased the growth performance of the phalanx clonal plants (A. calamus, S. tabernaemontani and B. umbellatus; Figs. 3 and 4). This may be because guerrilla clonal plants can spread rapidly and produce offspring vegetatively by forming flexible, spreading offspring ramets, which have access to dispersed resources. Phalanx clonal plants usually spread slowly and form aggregated clones, which have access to centralized resources [8, 29, 31]. In addition, some species exhibit a trade-off between the two growth forms in different habits and successional stages. For instance, Leymus secalinus can respond and adapt to small-scale heterogeneity in its resource supply by altering the plasticity of its spacer morphology [41]. Therefore, guerrilla clonal plants may be better suited to a heterogeneous distribution of soil nutrients, and phalanx clonal plants show significantly enhanced growth performance in homogeneous soil nutrient conditions.
Owing to their strong horizontal expansion ability and morphological plasticity, guerrilla clonal plants respond to nutrient conditions by concentrating their foraging organs (such as rhizomes, stolons, and corms) where nutrient levels are relatively high [12]. For example, T. orientalis produced more ramets and larger spacer diameters and lengths in the heterogeneous soil treatment (Fig. 4). In addition, guerrilla clonal plants can alter the branching angle and distribution of ramets as well as their biomass allocation ratio to obtain available resources [18, 38]. These specific clonal growth organs are characterized by different functional traits (such as dispersal, resource acquisition, storage, shoot cycling and protection) [32] and can adapt to diverse environments. Thus, clonal plants, especially guerrilla clonal plants, are widely used in the ecological remediation of various habitats.
Clonal plants often appear as pioneer species in the initial stages of community succession [26]. In the process of vegetation restoration, clonal plants play a dominant role in changing the vegetation community environment and maintaining community ecological function [11]. For example, Psammochloa villosa (a guerrilla clonal plant) can improve vegetation coverage and sand fixation in mobile dune patches [13]. Particle size is an important feature of soil, and it can greatly affect the growth of clonal plants [19]. In this experiment, the heterogeneous distribution of soil nutrients and the particle size may have inhibited the foraging behaviour of the phalanx clonal plants. This may have occurred because the mechanical resistance of the soil particles decelerated root growth and expansion [5].
Resources (light, water, nutrients, etc.) and environmental conditions (disturbances, geography, herbivory, etc.) exhibit spatial and temporal heterogeneity, which is ubiquitous within natural habitats [21, 45]. In this study, T. orientalis showed high ecological adaptability to heterogeneous resource habitats. In addition, the guerrilla clonal plants (T. orientalis, P. australis and S. stoloniferum) accumulated more belowground biomass in the heterogeneous soil, in which larger foraging organ masses may explore and occupy resource-rich patches (Fig. 1). These identical individuals can share resources and stress through the physical connections of their spacers [2, 42]. This is why the guerrilla growth is very common in early successional stages and disturbed habitats.
Conclusions
In conclusion, the heterogeneity of soil nutrients promotes the growth of guerrilla clonal plants, especially that of T. orientalis. The flexible clonal structure of guerrilla clonal plants can be used to effectively utilize resources. Future studies should focus on how various ecological factors, such as temperature and competition, affect growth responses to resource heterogeneity.
Availability of data and materials
The data are available from the corresponding author upon reasonable request and with permission from Wuhan University.
Abbreviations
- T. orientalis :
-
Typha orientalis
- A. calamus :
-
Acorus calamus
- S. tabernaemontani :
-
Schoenoplectus tabernaemontani
- B. umbellatus :
-
Butomus umbellatus
- S. yagara :
-
Scirpus yagara
- P. australis :
-
Phragmites australis
- S. stoloniferum :
-
Sparganium stoloniferum
- HE:
-
Heterogeneous soil treatment
- HO:
-
Homogeneous soil treatment
References
Alpert, P., 1997. Division of labour in clonal plants. The ecology and evolution of clonal plants, 137-154.
Alpert P. Clonal integration in Fragaria chiloensis differs between populations: ramets from grassland are selfish. Oecologia. 1999;120:69–76.
Bartels SF, Chen HY. Is understory plant species diversity driven by resource quantity or resource heterogeneity? Ecology. 2010;91:1931–8.
Bernard JM. Life history and vegetative reproduction in Carex. Can J Bot. 1990;68:1441–8.
Bengough AG, McKenzie BM, Hallett PD, Valentine TA. Root elongation, water stress, and mechanical impedance: a review of limiting stresses and beneficial root tip traits. J Exp Bot. 2011;62:59–68.
Board E. Flora of China, vol. 23. Bejing: Sci Press; 2010.
Bonanno G, Vymazal J, Cirelli GL. Translocation, accumulation and bioindication of trace elements in wetland plants. Sci Total Environ. 2018;631:252–61.
Cowie, N.R., Watkinson, A.R., Sutherland, W.J., 1995. Modelling the growth dynamics of the clonal herb”Anemone nemorosa” L. in an ancient coppice wood. Abstracta Botanica, 35-49.
Crowe, A.S., Rochfort, Q., Exall, K., Marsalek, J., 2007. Controlling urban stormwater pollution by constructed wetlands: A Canadian perspective. International Journal of Water 3.
Day K, Hutchings M, John E. The effects of spatial pattern of nutrient supply on the early stages of growth in plant populations. J Ecol. 2003;91:305–15.
de Kroon, H., van Groenendael, J., 1997. The ecology and evolution of clonal plants Backhuys Publ. Leiden.
de Kroon H, Visser EJ, Huber H, Mommer L, Hutchings MJ. Environment, 2009.A modular concept of plant foraging behaviour: the interplay between local responses and systemic control. Plant Cell. 2009;32:704–12.
Dong M, Alaten B. Clonal plasticity in response to rhizome severing and heterogeneous resource supply in the rhizomatous grass Psammochloa villosa in an Inner Mongolian dune, China. Plant Ecol. 1999;141:53–8.
Eckert, C.G., 2002. The loss of sex in clonal plants, Ecology and evolutionary biology of clonal plants. Springer, pp. 279-298.
Eriksson O. Patterns of ramet survivorship in clonal fragments of the stoloniferous plant Potentilla anserina. Ecology. 1988;69:736–40.
García-Palacios P, Maestre FT, Gallardo A. Soil nutrient heterogeneity modulates ecosystem responses to changes in the identity and richness of plant functional groups. J Ecol. 2011;99:551–62.
Hagiwara Y, Kachi N, Suzuki J-I. Effects of temporal heterogeneity of water supply on the growth of Perilla frutescens depend on plant density. Ann Bot. 2010;106:173–81.
Henry HA, Aarssen LW. Inter-and intraspecific relationships between shade tolerance and shade avoidance in temperate trees. Oikos. 2001;93:477–87.
Huang L, Dong BC, Xue W, Peng YK, Zhang MX, Yu FH. Soil particle heterogeneity affects the growth of a rhizomatous wetland plant. PLoS ONE. 2013;8(7):e69836.
Humphrey LD, Pyke DA. Demographic and growth responses of a guerrilla and a phalanx perennial grass in competitive mixtures. J Ecol. 1998;86:854–65.
Hutchings MJ, John EA. The effects of environmental heterogeneity on root growth and root/shoot partitioning. Ann Bot. 2004;94:1–8.
Hutchings MJ, Wijesinghe DK. Performance of a clonal species in patchy environments: effects of environmental context on yield at local and whole-plant scales. Evol Ecol. 2008;22:313–24.
Moore JE, Franklin SB. Water stress interacts with early arrival to influence interspecific and intraspecific priority competition: a test using a greenhouse study. J Veg Sci. 2012;23:647–56.
Navas, M.L., Garnier, E., 1990. Demography and growth forms of the clonal perennial Rubia peregrina in Mediterranean vineyard and unmanaged habitats. The Journal of Ecology, 691-712.
Oborny, B., Bartha, S., 1995. Clonality in plant communities-an overview. Abstracta Botanica, 115-127.
Prach K, Pyšek P. Clonal plants—what is their role in succession? Folia Geobotanica. 1994;29:307–20.
Roiloa S, Retuerto R. Small-scale heterogeneity in soil quality influences photosynthetic efficiency and habitat selection in a clonal plant. Ann Bot. 2006;98:1043–52.
Saiz H, Bittebiere AK, Benot ML, Jung V, Mony C. Understanding clonal plant competition for space over time: a fine-scale spatial approach based on experimental communities. J Veg Sci. 2016;27:759–70.
Schmid B. Some ecological and evolutionary consequences of modular organization and clonal growth in plants. Evolutionary Trends Plants. 1990;4:25–34.
Seelig B. Water quality and wetland function in the Northern Prairie Pothole Region. Fargo: North Dakota State University; 2006.
Song M, Dong M. Clonal plants and plant species diversity in wetland ecosystems in China. J Veg Sci. 2002;13:237–44.
Sosnova M, van Diggelen R, Klimešova J. Distribution of clonal growth forms in wetlands. Aquat Bot. 2010;92(1):33–9.
van der Waal C, de Kroon H, Heitkönig IM, Skidmore AK, van Langevelde F, de Boer WF, Slotow R, Grant RC, Peel MP, Kohi EM. Scale of nutrient patchiness mediates resource partitioning between trees and grasses in a semi-arid savanna. J Ecol. 2011;99:1124–33.
Van Groenendael, J., 1997. Clonal plant architecture: a comparative analysis of form and function, The ecology and evolution of clonal plants. Backhuys Publishers Leiden, pp. 1-29.
Van Kleunen M, Fischer M. Adaptive evolution of plastic foraging responses in a clonal plant. Ecology. 2001;82:3309–19.
Wang YJ, Shi XP, Meng XF, Wu XJ, Luo FL, Yu FH. Effects of spatial patch arrangement and scale of covarying resources on growth and intraspecific competition of a clonal plant. Front Plant Sci. 2016;7:753.
Weiss PT, Asce M, Gulliver JS, Asce F, Erickson AJ, Asce SM. Cost and pollutant removal of storm-water treatment practices. J Water Resources Planning Management. 2007;133:218–29.
Xiao K, Yu D, Wang J. Habitat selection in spatially heterogeneous environments: a test of foraging behaviour in the clonal submerged macrophyte Vallisneria spiralis. Freshw Biol. 2006;51:1552–9.
Xue W, Huang L, Yu FH, Bezemer TM. Intraspecific aggregation and soil heterogeneity: competitive interactions of two clonal plants with contrasting spatial architecture. Plant Soil. 2018;425:231–40.
Xue,W., Huang, L., Yu, F. H., 2020. Importance of starting points in heterogeneous environments: interactions between two clonal plants with contrasting spatial architectures. J Plant Ecol. rtaa018.
Ye XH, Yu FH, Dong M. A trade-off between guerrilla and phalanx growth forms in Leymus secalinus under different nutrient supplies. Ann Bot. 2006;98:187–91.
Yu FH, Dong M, Krüsi B. Clonal integration helps Psammochloa villosa survive sand burial in an inland dune. New Phytol. 2004;162:697–704.
Yu FH, Wang N, Alpert P, He WM, Dong M. Physiological integration in an introduced, invasive plant increases its spread into experimental communities and modifies their structure. Am J Bot. 2009;96:1983–9.
Yu, H., Shen, N., Guan, X., Yu, S., Yu, D., Liu, C., 2019a. Influence of Soil Nutrient Heterogeneity and Competition on Sprouting and Ramets Growth of Alternanthera philoxeroides. CLEAN–Soil, Air, Water 47, 1800182.
Yu H, Shen N, Yu D, Liu C. Effects of temporal heterogeneity of water supply and spatial heterogeneity of soils nutrients on the growth and intraspecific competition of Bolboschoenus yagara depend on plant density. Front Plant Sci. 2018;9:1987.
Yu H, Shen N, Yu D, Liu C. Clonal integration increases growth performance and expansion of Eichhornia crassipes in littoral zones: a simulation study. Environ Exp Bot. 2019;159:13–22.
Acknowledgements
We thank Xiaoliang Zhang, Fei Ma, Jingwen Hu, Siqi Yu and Lifu Cui for assistance with the research.
Funding
The authors gratefully acknowledge funding support from the Special Foundation of National Science and Technology Basic Research [Grant Number 2013FY112300], Major Science and Technology Program for Water Pollution Control and Treatment [Grant Number 2015ZX07503-005], the National Natural Science Foundation of China [Grant Number 31,900,281] and the China Postdoctoral Science Foundation [Grant Number 2019M650634].
Author information
Authors and Affiliations
Contributions
HWY Methodology, writing-original draft, data curation, Formal analysis. LGW Investigation, Conceptualization. CHL Investigation, Software. DY Software, Funding acquisition, Supervision. JHQ Conceptualization, Funding acquisition, Supervision, Writing–review and editing. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Both of the plant species were common and naturally distributed in Inner Mongolia, China. No specific permissions were required for these locations. This study did not involve endangered or protected species.
Consent for publication
This manuscript has not been published or presented elsewhere in part or in entirety and is not under consideration by another journal.
Competing interests
We have read and understood your journal’s policies, and we believe that neither the manuscript nor the study violates any of these policies. There are no conflicts of interest to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Yu, H., Wang, L., Liu, C. et al. Effects of a spatially heterogeneous nutrient distribution on the growth of clonal wetland plants. BMC Ecol 20, 59 (2020). https://doi.org/10.1186/s12898-020-00327-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12898-020-00327-1