
Abstract
Background
Trichothiodystrophy nonphotosensitive 1 (TTDN1) is a disease with mental retardation, brittle hair. Some cases of the diseases are caused by mutations of the MPLKIP gene.
Methods
We carefully identified the clinic characteristics, the sulfur level and pattern of the hair shafts of a female patient of with the symptom of hypergonadotropic hypogonadism, and of her parents and brother whose are healthy. We also collected the blood sample of the patient and performed the exon sequencing. One G insertion in MPLKIP was identified after analyzing the obtained exon sequencing profile. The G insertion sites in the patient, her parents and brother, were verified using Sanger sequencing. The G insertion in MPLKIP were compared to the dbSNP.
Results
The female patient of TTDN1 carries a homozygous G insertion (rs747470385) in the MPLKIP gene. The parents and brother of the patient are heterozygous carriers of the same mutation, but are healthy. The hair shafts of the patient had a tiger-tail pattern with relatively low sulfur levels. To the best of our knowledge, this is the first report that autosomal recessive inheritance of the G insertion in the MPLKIP gene results in TTDN1.
Conclusion
Our results indicate that the homozygotic G insertion in MPLKIP results in the TTDN1 with hypergonadotropic hypogonadism, while heterozygous carriers of the same mutation have no symptoms and healthy. These results provide novel insights into the association of mutations in MPLKIP and TTDN1 with hypergonadotropic hypogonadism.
Similar content being viewed by others

Background
Trichothiodystrophy (TTD) is an autosomal recessive disorder of neuroectodermal origin. This condition is characterized by a cross-banding pattern, i.e. tiger-tail, under polarized light and low sulfur content in the hair shafts, which are associated with variable and neuroectodermal symptoms. The condition manifests with brittle hair, intellectual disability, dwarfism, decreased fertility, microcephaly, abnormal facial features, ichthyosis premature aging, nail dystrophies, and a propensity for respiratory infections [1]. TTD is divided into two types: photosensitive (TTD1-3) and nonphotosensitive (TTD4 or TTDN1). Photosensitivity exists in about 50% of TTD patients, which is due to reduced amounts of transcription/repair factor IIH involved in global genome and transcription repair [2, 3]. Mutations in the MPLKIP gene cause some cases of TTDN1.
MPLKIP is located on chromosome 7p14.1 and includes two coding exons that encode a protein called M-phase-specific PLK1-interacting protein, which can interact with PLK1 protein. Based on the function of the PLK1 protein and its location in the nucleus, the MPLKIP protein plays a role in regulating the cell cycle. It is ubiquitously expressed in epidermis, hair follicles, brain, heart, liver, kidney, skeletal muscle, pancreas, lung, and placenta [4–6]. The precise function of MPLKIP has not yet been determined, but clinical symptoms of TTDN1 result from mutation of the gene [4].
Results
Physical characteristics of the patient
A 16 year old girl in China with primary amenorrhea and mental retardation was sent to the endocrinology clinic by her parents. The patient was born full term via spontaneous vaginal delivery. Her birth weight and length are unknown. Her parents are cousins related by blood. When the patient was 7 years old in primary school, she was observed to have significant mental retardation. Patients had no complaints of tetany, ostalgia, hearing loss, or hyposmia. Menarche did not occur until 16 years of age. A small amount of bleeding due to an artificial menstrual cycle induced by local doctors occurred for 5 months. The heights of her father and mother were 171 and 154 cm, respectively. The development and intelligence of her elder brother was normal. There was no history of any endocrine illness in other family members.
Upon physical examination, her blood pressure was 100/66 mmHg, temperature 36 ∘C, pulse rate 82 beats/min, and respiratory rate 20 breaths/min (see Table 1). The patient had a height of 150 cm (<3rd centile on the 2009 Chinese Academy of Pediatrics growth chart) with a weight of 39 kg (Table 1). Arm span was 145 cm with an upper/lower segment ratio of 0.9:1 (Table 1). There was a lack of secondary sexual characteristics Tanner BII and PH I (Table 1). Bilateral cataracts were found (Table 1). The oral cavity had no enamel hypoplasia. The neck was not short with excess skin and no goiter. There were no notable special facial features except sparse eyebrows and brittle hair. Abnormal forearm carrying angles were not found. The fifth middle phalanxes on their both hands were short. Chvostek’s sign, Trousseau’s sign, and signs of neuromuscular irritability were not present. Her skin was normal with no UV light sensitivity or ichthyosis issues.
Testing revealed a follicle-stimulating hormone (FSH) of 127.4 mIU/ml (normal range: 3.5-12.5 mIU/ml), progesterone (PROG): 0.52 nmol/l (0.6-4.7 nmol/l), luteal hormone (LH): 65.25 mIU/ml (2.4-12.6 mIU/ml), estradiol (E2) 18.35 pmol/l (45.4-854 pmol/l), prolactin (PRL): 17.3 ng/ml (3.4-24.1 ng/ml) and testosterone (TEST): 0.18 nmol/ml (0.16-1.33 nmol/ml) (see Table 1). Serum 25 hydroxy vitamin D was 19.68 ng/ml (>30 ng/ml). Both arginine and levodopa stimulation tests for growth hormone were carried out, revealing normal growth hormone responses to stimulation. Liver, renal, thyroid, coagulation, fasting plasma glucose, blood fat, and dynamic cortisol level tests were normal. The serum alkaline phosphatase, calcium, phosphorus, 24 hr urinary calcium, and parathormone were within reference ranges. However, the 24 hr urinary phosphorus was 5.99 mmol/d (12.9-42 mmol/d).
MRI of the hypophysis was normal. X-rays of the wrist and hand suggested that skeletal age was around 12 years, which did not correspond to chronological age. Double energy x-ray bone density suggested osteoporosis (Z: -4.5). Wechsler intelligence scale revealed her intelligence quotient was lower than 40 points. Electrocardiogram showed sinus rhythm, and first degree a-v block and ultrasonic cardiogram were normal. B ultrasonic examination showed that Liver, bile, pancreas, spleen and kidney were normal and both the uterus (43 mm ×28 mm ×15 mm) and ovaries (right 16 mm ×5 mm, left 14 mm ×4.5 mm) were smaller and the endometrium was thin. Her hair shafts were examined by polarized light microscopy and had a characteristic tiger-tail pattern (Fig. 1a), which was not noticed in her mother’s hair (Fig. 1b). The sulfur level of the hair measured by atomic emission spectrometry was 1.905 mg/kg significantly lower than both parents and a healthy control (patient: 1.905 mg/kg compared to mother: 4.177 mg/kg, father: 4.811 mg/kg, and a normal control: 4.62 mg/kg) (P = 0.02, t-test).

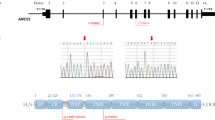
The brittle hair of the patient and G insertion identified in the patient. a The typical alternating dark and light “tiger tail” banding was seen by polarized light microscopy in hair of the patient. b The typical alternating dark and light “tiger tail” banding was not seen by polarized light microscopy in hair of the patient’s mother. c to f The Sanger sequencing of the mutated region in MPLKIP for the patient, her mother, her father and her elder brother, respectively. g The position of the G insertion in the MPLKIP gene. Query is the sequence obtained in Sanger sequencing. Ref. is the sequence of MPLKIP mRNA (NM_138701.3). h The amino acid sequences of wild type MPLKIP gene (top) and the mutated MPLKIP gene with the identified G insertion. Only the first 58 amino acids of the wild type MPLKIP protein are shown. i The G insertion was examined in UCSC Genome Browser to be compared to the dbSNP (v150). The G insertion in MPLKIP is rs747470385 (-/G), as indicated by the blue rectangle
Characterizing the genomic mutations of the patient
Chromosomal karyotyping revealed the patient had a normal female karyotype (46, XX). In order to acquire a molecular diagnosis, exon sequencing was performed for the patient. After analyzing the exon sequencing profile, we found a G insertion in the first exon of the MPLKIP gene. We then confirmed the G insertion with Sanger sequencing, for the patient, and her father, mother, and elder brother (see green boxes in Fig. 1c to f). From Fig. 1c, it could be seen that the Sanger sequencing of the patient is clear and has no minor peaks in the region. However, in the Sanger sequencing results of her mother, father and brother, two nucleotides after the G insertion sites (in the shaded regions of Fig. 1d to f, respectively) have minor or almost equal peaks, suggesting heterozygous genotypes. To summarize, the patient is homozygous for the G insertion, while her father, mother, and brother are heterozygous carriers (see Fig. 1d to f).
We carefully examined the G insertion in MPLKIP and found that the G insertion locates immediately after the 177th nucleotide in MPLKIP transcript (Fig. 1g). The normal MPLKIP protein consists of 179 amino acids. The G insertion leads to a significantly short amino acid sequence with only 58 residuals, and the sequence changes after the 32nd amino acid (see Fig. 1h). Thus, the homozygous G insertion may lead to total deletion of functional MPLKIP protein in the patient. We compared the identified G insertion to the latest dbSNP (v150) using UCSC Genome Browser and found that the G insertion is a reported SNP, rs747470385 (-/G) (Fig. 1i).
The protein-protein interaction network of MPLKIP
We investigated the functional roles of MPLKIP by examining its protein-protein interaction network as well. As shown in Fig. 2a, MPLKIP serves as a bridge between the RNA Polymerase II complex (left part) and many other co-factors. Among proteins interacting with MPLKIP, SBDS encodes a highly conserved protein that plays an essential role in ribosome biogenesis. SBDS protein interacts with elongation factor-like GTPase 1 to disassociate eukaryotic initiation factor 6 from the late cytoplasmic pre-60S ribosomal subunit allowing assembly of the 80S subunit. Mutations within SBDS are associated with the autosomal recessive disorder Shwachman-Bodian-Diamond syndrome (or Shwachman-Diamond syndrome), which is a rare congenital disorder characterized by exocrine pancreatic insufficiency, bone marrow dysfunction, skeletal abnormalities and short stature. FAM120C encodes a potential transmembrane protein and lies in a region where mutations and deletions have been associated with intellectual disability and autism. Some of the symptoms caused by mutations in SBDS and FAM120C, such as short stature and intellectual disability, are similar to shown in our patient.

The protein-protein interaction network of MPLKIP. a The proteins interacting with MPLKIP examined using STRING [22]. An edge between two proteins (shown as nodes) means these two proteins are interacting proteins. Purple and azure edges stand for experimentally determined interactions and interactions from curated databases, respectively. Green, red, and blue edges stand for predicted interactions from gene neighborhood, gene fusions and gene co-occurrence, respectively. Light green and black edges stand for other interactions from text mining and co-expression, respectively. b Some of the enriched GO terms for genes in Part (a). c The enriched KEGG pathways for genes in Part (a)
We then performed Gene Onotology (GO) and KEGG pathway enrichment analysis for the interacting proteins of MPLKIP in Fig. 2a using KOBAS2 [7]. As shown in Fig. 2b, these genes are significantly enriched in many RNA Polymerase II related transcription GO terms, such as Transcription elongation from RNA polymerase II promoter, DNA-templated transcription, elongation, and Transcription initiation from RNA polymerase II promoter, DNA-directed RNA polymerase II, holoenzyme, DNA-directed RNA polymerase complex, Nucleic acid binding, and RNA polymerase II carboxy-terminal domain kinase activity. In Fig. 2c, it is shown that these genes are enriched in transcription related pathways too, such as Basal transcription factors and Nucleotide excision repair. Mutations in genes on the nucleotide excision repair pathway are associated with diseases, such as xeroderma pigmentosum (XP), Cockayne syndrome (CS) and trichothiodystrophy (TTD). In summary, these results suggest that the loss of MPLKIP caused by the G insertion will disrupt the interactions between many other co-factors and Pol II complex, and affect the Pol II involved transcription activities, which contributed to symptoms witnessed in the patient.
Treatments and consequences of the patients
Due to gonadal failure, the patient was initially given 0.5 mg estradiol valerate once a day. Low-dose oestrogen monotherapy enabled normal breast and uterine development. She had a vitamin D deficiency (serum 25 hydroxy vitamin D was 19.68 ng/ml) with osteoporosis, although osteoporosis in this patient was probably one of the phenotypes of hypogonadism; therefore, oral calcitriol and calcium carbonate were given at doses of 0.25 μg and 600 mg, respectively, once daily. Although she had a short stature, her parents did not want to treat the problem, so no treatments were given to correct her height.
The patient followed up with us after 3 months. Her breast developed to Tanner BIII and B ultrasonic examination showed both her uterus (61 mm ×42 mm ×26 mm) and ovaries (right one was 25 mm ×16 mm, while the left one was not clear) were bigger than before and the thickness of endometrium went from thin to 3 mm with estradiol increased to 330.6 pmol/L (normal range 45.4-854 pmol/L). Her bone mineral density did not increase with initial estradiol valerate treatment and a short treatment course with calcitriol. The patient was kept on oestrogen and vitamin D supplements and was advised to follow-up again after 3 months.
Discussion
The patient had a number of symptoms associated with endocrine disorders, including hypergonadotropic hypogonadism, short stature, and osteoporosis. Meanwhile, she presented with distinct characteristics of mental retardation, brittle hair with a tiger-tail pattern and low sulfur level, and cataract without photosensitivity. Many cases reported that TTDN1 manifested decreased fertility but hypergonadotropic hypogonadism confirmed ovarian lesions were the reason of gonad disorder in this paper.
The relationship between TTDN1 and MPLKIP was reported in 2005 [4]. Only 14 cases were reported in [8–12] after the study of Nakabayashi et al. [4]. As summarized in Table 2, there are different special phenotypic manifestations in addition to common symptoms that derive from various genetic mutations of MPLKIP. The homozygous splice mutation c.339 + 1G > A within MPLKIP results in mitral regurgitation [8]. Heller et al. [10] reported that mutations at two different loci cause autism spectrum disorder (see Table 2). Mutation c. 277delT results in seizure disorders [10], and mutation c.505dupA within the MPLKIP gene has been shown to cause severe renal failure [9]. Swagemakers et al. [13] reported that Pollitt syndrome patients, with mental and physical retardation and trichorrhexis nodosa, carry C326delA. Botta et al. [11] reported several cases of axial hypotonia and reduced motor coordination due to deletions at four different loci, and several cases with failure to thrive, very poor motor performances and speech due to deletions at two different loci (see Table 1). Nakabayashi et al. [4] and Przedborski et al. [14] reported that 187_188delGG causes ataxia.
Although the disease with low incidences displays many of the characteristics of endocrine disease, we hope it can be easily recognized by endocrinologists. This is the first report of autosomal recessive inheritance of the G insertion (rs747470385) and provides novel insights into endocrine symptoms associated with hypergonadotropic hypogonadism. Although the relationship between hypogonadism and the MPLKIP gene mutation is unclear, the gene plays a key role in regulating in cytokinesis and mitosis that may be a possible reason to explain the relationship.
TTDN1 should be considered in patients with hypogonadism. Hair shafts should be examined by polarized light microscopy and sulfur levels in the hair can be measured as a simple and convenient method for screening for TTDN1.
The wild-type MPLKIP gene is translated into a protein with 179 residues. Insertion of G at base 177 strongly shifts translation. Some termination codons occur in the mutant, implying the mutant protein may significantly differ from the wild-type protein. There are only short polypeptides of ≤58 residues if translated (see Fig. 1h). The homologous sequence of the mutant was not found in the Uniprot database [15]. For the short polypeptides, their secondary structures [16] and disorder sequence [17] predictions show the majority of the polypeptides are coils. It implies the polypeptides could be very flexible without any stable 3D structures. Therefore, the mutant proteins tend to degrade quickly. Therefore, all clinical symptoms in the patient may result from loss of function of the original protein. To summarize, the homozygous G insertion (rs747470385) in her MPLKIP gene caused trichothiodystrophy nonphotosensitive 1 (TTDN1) with hypergonadotropic hypogonadism.
Our results suggest the homozygous G insertion (rs747470385) could potentially be used as a genetic method to screen patients with hypergonadotropic hypogonadism for their diagnosis as TTDN1.
Conclusions
The contributions of this work are three folds. First, our results indicate that the homozygotic G insertion (rs747470385) cause TTDN1. Second, the homozygotic G insertion could potentially be used as a diagnostic marker of TTDN1. Third, our results indicate that the heterozygous rs747470385 does not lead to TTDN1. However, the marriage between heterozygous rs747470385 carriers should be prevented to minimize the risk of sick offspring.
Methods
Physical and clinical examinations were performed for the patient, her parents and her elder brother in the First People’s Hospital of Yunnan Province (The Kunhua Affiliated Hospital of Kunming University of Science and Technology, abbreviated as FPHY). The sulfur level of the hair was examined using inductively coupled plasma-atomic emission spectrometry at Kunming CTER Trace Element Measurement Research Center, Kunming, Yunnan, China.
The blood sample of the patient was collected in FPHY. The DNA sample was extracted and used for DNA subtotal exon sequencing at the RunningGene Inc, Beijing, China. The obtained exon sequencing profile had been deposited into the NCBI SRA database and available with the accession number SRR7427760. The obtained sequencing profile was aligned to human genome (hg38) with Bowtie2 [18]. The mutations were analyzed using SAMTools [19] and BCFTools [20] from the alignment results of Bowtie2.
The G insertion in the patient, her mother, father, and elder brother, were verified using Sanger sequencing with the forward primer of CTGGAGTAGGAGCCAGGGT and reverse primer of GGAGGGCCGGTTGATACAG. After examining results of Sanger sequencing, a homozygous G insertion was found in the first exon of the MPLKIP gene of the patient. Heterozygous G insertions at the same positions were found in the parents and brother of the patient. The obtained G insertion was compared to the latest dbSNP using the UCSC Genome Browser [21].
The protein-protein interaction networks of MPLKIP was examined by using STRING [22]. A minimum required interaction score of 0.4 and the second shell with no more than 20 interactors were used when searching the protein “MPLKIP” in the species of “Homo sapiens” in STRING. The enriched Gene Ontology terms and KEGG pathways of proteins interacting with MPLKIP were analyzed by using KOBAS2 [7].
Abbreviations
- BMD:
-
Bone Mineral Density
- CS:
-
Cockayne syndrome
- E2:
-
Estradiol
- FSH:
-
Follicle-Stimulating Hormone
- GH:
-
Growth Hormone
- GO:
-
Gene Onotology
- LH:
-
Luteal Hormone
- MRI:
-
Magnetic Resonance Imaging
- PRL:
-
Prolactin
- PROG:
-
progesterone
- TEST:
-
Testosterone
- TTD:
-
trichothiodystrophy
- TTDN1:
-
trichothiodystrophy nonphotosensitive 1
- XP:
-
xeroderma pigmentosum
References
Faghri S, Tamura D, Kraemer KH, DiGiovanna JJ. Trichothiodystrophy: a systematic review of 112 published cases characterises a wide spectrum of clinical manifestations. J Med Genet. 2008; 45(10):609–21.
Botta E, Nardo T, Lehmann AR, Egly J-M, Pedrini AM, Stefanini M. Reduced level of the repair/transcription factor tfiih in trichothiodystrophy. Hum Mol Genet. 2002; 11(23):2919–28.
Vermeulen W, Bergmann E, Auriol J, Rademakers S, Frit P, Appeldoorn E, Hoeijmakers JH, Egly J-M. Sublimiting concentration of tfiih transcription/dna repair factor causes ttd-a trichothiodystrophy disorder. Nat Genet. 2000; 26(3):307.
Nakabayashi K, Amann D, Ren Y, Saarialho-Kere U, Avidan N, Gentles S, MacDonald JR, Puffenberger EG, Christiano AM, Martinez-Mir A, Salas-Alanis JC, Rizzo R, Vamos E, Raams A, Les C, Seboun E, Jaspers NGJ, Beckmann JS, Jackson CE, Scherer SW. Identification of c7orf11 (ttdn1) gene mutations and genetic heterogeneity in nonphotosensitive trichothiodystrophy. Am J Hum Genet. 2005; 76(3):510–6.
Nakabayashi K, Fernandez BA, Teshima I, Shuman C, Proud VK, Curry CJ, Chitayat D, Grebe T, Ming J, Oshimura M, et al.Molecular genetic studies of human chromosome 7 in russell–silver syndrome. Genomics. 2002; 79(2):186–96.
Winkles JA, Alberts GF. Differential regulation of polo-like kinase 1, 2, 3, and 4 gene expression in mammalian cells and tissues. Oncogene. 2005; 24(2):260.
Xie C, Mao X, Huang J, Ding Y, Wu J, Dong S, Kong L, Gao G, Li C-Y, Wei L. Kobas 2.0: a web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011; 39(suppl_2):316–22.
Shah K, Ali RH, Ansar M, Lee K, Chishti MS, Abbe I, Li B, Smith JD, Nickerson DA, Shendure J, et al.Mitral regurgitation as a phenotypic manifestation of nonphotosensitive trichothiodystrophy due to a splice variant in mplkip. BMC Med Gen. 2016; 17(1):13.
Pode-Shakked B, Marek-Yagel D, Greenberger S, Pode-Shakked N, Pras E, Barzilai A, Yassin S, Sidi Y, Anikster Y. A novel mutation in the c7orf11 gene causes nonphotosensitive trichothiodystrophy in a multiplex highly consanguineous kindred. Eur J Med Gen. 2015; 58(12):685–8.
Heller ER, Khan SG, Kuschal C, Tamura D, DiGiovanna JJ, Kraemer KH. Mutations in the ttdn1 gene are associated with a distinct trichothiodystrophy phenotype. J Investig Dermatol. 2015; 135(3):734–41.
Botta E, Offman J, Nardo T, Ricotti R, Zambruno G, Sansone D, Balestri P, Raams A, Kleijer WJ, Jaspers NG, Sarasin A, Lehmann AR, Stefanini M. Mutations in the c7orf11 (ttdn1) gene in six nonphotosensitive trichothiodystrophy patients: no obvious genotype–phenotype relationships. Hum Mutat. 2007; 28(1):92–6.
La Serna-Infantes J, Pastor MC, Trubnykova M, Velásquez FC, Sotomayor FV, Barriga HA. Novel contiguous gene deletion in peruvian girl with trichothiodystrophy type 4 and glutaric aciduria type 3. Eur J Med Gen. 2018; 61(7):388–92.
Swagemakers SM, Jaspers NG, Raams A, Heijsman D, Vermeulen W, Troelstra C, Kremer A, Lincoln SE, Tearle R, Hoeijmakers JH, van der Spek PJ. Pollitt syndrome patients carry mutation in ttdn1. Meta gene. 2014; 2:616–8.
Przedborski S, Ferster A, Goldman S, Wolter R, Song M, Tonnesen T, Pollitt RJ, Vamos E. Trichothiodystrophy, mental retardation, short stature, ataxia, and gonadal dysfunction in three moroccan siblings. Am J Med Genet. 1990; 35:566–73.
Shen Y, Maupetit J, Derreumaux P, Tufféry P. Improved pep-fold approach for peptide and miniprotein structure prediction. J Chem Theory Comput. 2014; 10(10):4745–58.
Buchan DW, Minneci F, Nugent TC, Bryson K, Jones DT. Scalable web services for the psipred protein analysis workbench. Nucleic Acids Res. 2013; 41(W1):349–57.
Linding R, Jensen LJ, Diella F, Bork P, Gibson TJ, Russell RB. Protein disorder prediction: implications for structural proteomics. Structure. 2003; 11(11):1453–9.
Langmead B, Salzberg SL. Fast gapped-read alignment with bowtie 2. Nat Methods. 2012; 9(4):357–9.
Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, Marth G, Abecasis G, Durbin R. 1000 Genome Project Data Processing Subgroup: The sequence alignment/map format and samtools. Bioinformatics. 2009; 25(16):2078–9.
Danecek P, McCarthy SA, Consortium H, Durbin R. A method for checking genomic integrity in cultured cell lines from snp genotyping data. PLoS ONE. 2016; 11(5):1–13.
Rosenbloom KR, Armstrong J, Barber GP, Casper J, Clawson H, Diekhans M, Dreszer TR, Fujita PA, Guruvadoo L, Haeussler M, Harte RA, Heitner S, Hickey G, Hinrichs AS, Hubley R, Karolchik D, Learned K, Lee BT, Li CH, Miga KH, Nguyen N, Paten B, Raney BJ, Smit AFA, Speir ML, Zweig AS, Haussler D, Kuhn RM, Kent WJ. The UCSC Genome Browser database: 2015 update. Nucleic Acids Res. 2015; 43(D1):670–81.
Szklarczyk D, Morris JH, Cook H, Kuhn M, Wyder S, Simonovic M, Santos A, Doncheva NT, Roth A, Bork P, et al.The string database in 2017: quality-controlled protein–protein association networks, made broadly accessible. Nucleic Acids Res. 2016; 45(Database issue):D362–8.
Acknowledgements
Not applicable.
Funding
This work was funded by the two grants of National Natural Science Foundations of China (number 81660146) and a Health System Academic Leader Project of Yunnan province (D-201661) to YZhou; and by two grants of National Natural Science Foundations of China (numbers 31460295 and 31760314) to YZheng; and by the grant of National Natural Science Foundations of China (number 31401130) to YC. Publication of this article was sponsored by National Natural Science Foundations of China (number 81660146) grant.
Availability of data and materials
The one obtained exon sequencing profile had been deposited into the NCBI SRA database and available with the accession number SRR7427760.
About this supplement
This article has been published as part of BMC Medical Genetics Volume 19 Supplement 1, 2018: Proceedings of the 29th International Conference on Genome Informatics (GIW 2018): medical genetics. The full contents of the supplement are available online at https://bmcmedgenet.biomedcentral.com/articles/supplements/volume-19-supplement-1.
Author information
Authors and Affiliations
Contributions
YKZ, ZL, and YZ conceived and designed the research. YKZ, XY, YC, ZL, and HS performed the physical and clinical examinations for the patient. YKZ collected the blood samples for the patient, her mother, father and brother. YKZ, YC and ZL read the related literature. YZ and LL designed the computational pipeline for analyzing exon sequencing profile and Sanger sequencing results. ZL, YC, LL, and YZ analyzed the exon sequencing profile and Sanger sequencing results. YKZ and YZ wrote the manuscript. All authors have read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The study was approved by the Medicine Ethics Committee of The First People’s Hospital of Yunnan Province that is composed of 17 members, including experts of laws, scientists and clinicians with relevant expertise. The patient, her parents and brother, agreed on the research and the publication of the current work.
Consent for publication
The patient, her parents and brother, agreed on the research and the publication of the current work.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Zhou, YK., Yang, XC., Cao, Y. et al. A homozygous G insertion in MPLKIP leads to TTDN1 with the hypergonadotropic hypogonadism symptom. BMC Med Genet 19 (Suppl 1), 214 (2018). https://doi.org/10.1186/s12881-018-0723-5
Published:
DOI: https://doi.org/10.1186/s12881-018-0723-5