
Abstract
Background
It is unclear whether the offspring of subclinical hypothyroidism (SCH) pregnant rats still have abnormal cardiac development, and whether early intervention with L-T4 can improve the abnormality of these offspring. Therefore, the aim of this study was to investigate the effect of early L-T4 intervention on the heart development of offspring of SCH pregnant rats and its possible molecular mechanism.
Methods
Eighty female Wistar rats were randomly divided into Sham group (placebo control), SCH group, LT4-E10 group (L-T4 treatment started on the 10th day of gestation), and LT4-E13 group (L-T4 treatment started on the 13th day of gestation). Each group was further divided into E16 (16th day of gestation), E18 (18th day of gestation), P5 (5th day postnatal day), and P10 (10th day postnatal day) subgroups. The levels of serum TT4 and TSH, the ratio of heart weight to body weight of offspring rats, the expression of metabolic enzymes, and the histopathology of cardiomyocytes were determined. To elucidate the effects of L-T4 on cardiac development of offspring of SCH pregnant rats, the expression levels of GATA4, Nkx2–5 and proteins involved in BMP4/Smad4 signaling pathway were detected by immunohistochemistry, real time quantitative polymerase chain reaction and Western blotting to elucidate the molecular mechanism of L-T4 regulating the heart development of the offspring of SCH pregnant rats.
Results
Compared with Sham group, serum TSH was significantly increased in SCH pregnant rats. Moreover, early L-T4 intervention significantly reduced the levels of serum TSH. Compared with the offspring in the SCH group, early L-T4 intervention significantly increased the heart weight, heart weight to body weight ratio, the activities of succinate dehydrogenase (SDH), Na+/K+-ATPase and Ca2+-ATPase, but reduced myocardial cell shrinkage and nuclear staining, hyperemia/congestion and vacuolar degeneration. In addition, early L-T4 intervention not only significantly increased the mRNA and protein expression of Gata4 and Nkx2–5, but also increased the protein expression involved in BMP4/Smad4 signal pathway in myocardium of the offspring of SCH pregnant rats.
Conclusions
Early L-T4 intervention can regulate the cardiac development of the offspring of SCH pregnant rats by activating BMP4/Smad4 signaling pathway and increasing the expression of Gata4 and Nkx2–5 proteins.
Similar content being viewed by others

Background
Subclinical hypothyroidism (SCH) is one of the most common thyroid diseases during pregnancy, affecting 2 to 3% of the global population. SCH was defined as increased serum thyroid-stimulating hormone (TSH) during pregnancy, but the level of free thyroxine (FT4) is normal [1]. Pregnancy has a great influence on the thyroid function of pregnant women. During pregnancy, human chorionic gonadotropin (HCG) weakly binds and stimulates TSH receptor and stimulates thyroid to produce more thyroxine. Meanwhile, the volume of thyroid increases by 10 to 40%, and the demand for thyroxine and iodine also increases by 50%, which will inevitably lead to hypothyroidism in the third trimester of pregnancy. Patients diagnosed with SCH before pregnancy have more severe symptoms of thyroid hormone deficiency during pregnancy [2]. It is known that poor control of SCH during pregnancy is related to pregnancy complications and growth retardation of offspring. In addition, women with hypothyroidism have an increased risk of hypertension and fetal death. These data indicate that there is a certain relationship between pregnant women with SCH and the risk of fetal disease [3].
Since the cardiovascular system is rich in thyroid receptor, it is relatively sensitive to the change of TSH levels [4]. Previous studies have showed that TSH levels above 7.0 mIU/L led to abnormalities in lipid metabolism, oxidative stress and endothelial function, thereby increasing the risk of atherosclerosis and congestive heart failure. In addition, patients with SCH under 65 years of age were associated with increased risk of coronary heart disease, heart failure, and cerebrovascular diseases. Patients with TSH levels above 10 mIU/L have a higher risk of heart failure with lower ejection fraction compared to participants with normal thyroid function [5]. The left ventricular diastolic dysfunction caused by SCH may be related to endothelial dysfunction, arterial stiffness and inflammation [6]. Another clinical study indicated that patients with SCH often had a slower heart rate and shorter corrected QT, which may be closely related to cardiac systolic dysfunction [7]. Longer P-wave duration, longer PR interval and decreased voltage were observed in patients with SCH. In addition, age and gender were known to be closely related to the occurrence of SCH [8]. However, it is still unclear the effect of pregnant women with SCH on fetal heart development and its possible molecular mechanism.
Although it is well known that women with hypothyroidism should increase the dose of levothyroxine during pregnancy, the exact timing and dose are not clear. A previous study found that women with hypothyroidism took administrated 150 μg/day of levothyroxine, a manufactured form of the thyroid hormone thyroxine (T4), during13 weeks of pregnancy, which can lead to an increase in TSH or a decrease in FT4 [9]. It has been confirmed that the demand for exogenous L-T4 dose of most pregnant women increase from 25 to 30% during the first 4 to 6 weeks of pregnancy and gradually increased within 16 to 20 weeks of pregnancy, and then stabilize until delivery [10]. It was reported that compared with healthy people, the left ventricular systolic and diastolic function of SCH patients was slightly affected. Moreover, L-T4 treatment reversed the change of left ventricular systolic and diastolic function of SCH patients, but has no significant effect on systolic pressure [11]. A similar study also found that for children with hypothyroidism and ventricular diastolic dysfunction, L-T4 treatment can significantly improve left ventricular dysfunction [12]. However, the effects of L-T4 on the fetal heart development of pregnant women with SCH and its possible mechanism are still unknown. Therefore, the aim of this preclinical study was to investigate the effect of L-T4 treatment on the fetal heart development in SCH pregnant rats and to explore its possible molecular mechanism.
Methods
Animals and groups
Eighty female Wistar rats, aged 6–8 weeks and weighing 180–200 g, were purchased from the Benxi Changsheng Biotechnology Co., Ltd. The animals were kept in the Experimental Animal Department of the Northern Theater General Hospital. The ambient temperature was maintained at 21–22 °C with 50–60% relative humidity. All rats were maintained under a 12:12-h light/dark cycle, with access to food and water ad libitum. Eighty rats were randomly divided into Sham group (placebo control), SCH group, LT4-E10 group (L-T4 treatment started on the 10th day of gestation), and LT4-E13 group (L-T4 treatment started on the 13th day of gestation). Each group was further divided into E16 (16th day of gestation), E18 (18th day of gestation), P5 (5th day postnatal day), and P10 (5th day postnatal day) subgroups. The animal experiment procedure was approved by the Animal Care and Use Committee of the General Hospital of Northern Theater Command, which complied with the national guidelines on the protection and use of laboratory animals.
Establishment of the SCH rat model
Establishment of the SCH rat model refers to the previously literature [13].The rats were injected with 3% pentobarbital sodium (0.1 mL/100 g) and underwent thyroidectomy, while Sham group rats underwent sham thyroid surgery. The rats were fed normally for 4 w after operation, then the blood was collected from the retroorbital venous plexus, and serum TSH and TT4 were detected. When serum TSH levels were higher than that in Sham group, the TT4 levels were lower than that in Sham group, confirming the successful establishment of the SCH rat model. Four weeks after surgery, rats in the SCH group were injected subcutaneously with L-thyroxine (L-T4, Sigma, USA) 1.0 μg/100 g/day on the neck. Sham group rats were injected subcutaneously with physiological saline (50 μL/100 g/day) on the neck. Calcium lactate (0.1% w/v) was added to the drinking water for all rats after surgery. Nine days later, all rats were mated with normal male rats (male: female = 1: 2). The pregnant rats were then kept in single cages until delivery. The day of vaginal plus was confirmed by microscopic observation and designated as E0. Serum and tissue samples were collected at E16, E18, P5 and P10. At the end of the experiment, all rats were anesthetized with pentobarbital (50 mg/kg, intraperitoneal) and euthanized by thoracotomy and hearts removal.
Measurement of TT4 and TSH
Blood samples obtained from the rats were immediately centrifuged at 13,000 g for 15 min and stored at − 80 °C for measurement of serum TT4 (#CEA452Ge, Youersheng, China) and TSH (#CEA463Ra, Youersheng, China) using a supersensitive chemiluminescence immunoassay.
ATPase activity analysis
The ATPase activity was detected using an ultramicro Ca2+-ATPase kit and Na+/K+-ATPase kit (#A070, Nanjing Jiancheng Bioengineering Institute, China) according to the manufacturer’s instructions. The succinate dehydrogenase (SDH) activity was measured by kit(A022, Nanjing Jiancheng Bioengineering Institute, China). Protein content was measured with a Coomassie blue protein assay kit (#WLA004a, wanleibio, China). ATPase activity was expressed as mol Pi liberated per mg protein per hour (mol Pi/(mg prot·hr)).
RNA isolation and quantitative real-time PCR
RNA was extracted from heart tissues from each group using TRIzol (Life Tech). First strand of cDNA was synthesized using total RNA and RT-PCR was carried out by TaqMan expression assays, and β-actin (#WL01845, wanleibio, China) was used as a internal reference. The sequences for Nkx2–5, Gata4, BMP4, Smad4 and GAPDH were performed through the ABI PRISM system. Primer sequences are shown: Bmp4 F 5′-ATCGTTACC TCAAGGGAGTGGA-3′; Bmp4 R 5′-ATCGTTACCTCAAGGGAGTGGA-3′; Samd4 F 5′-CGTTCACGAGGCATTTAC-3′; Samd4 R 5′-GGGAGGGAGTTGGACTG-3′; Nkx2–5 F 5′-TGGACAAAGCCGAGACAGAC-3′; Nkx2–5 R 5′-TCAGCGGGCGACAGGTA-3′; Gata4 F 5′-AAACGGAAGCCCAAGAAT-3′; Gata4 R 5′-GCTGCTGTGCCCATA GTGAG-3′; β-actin F 5′-GGAGATTACTGCCCTGGCTCCTAGC-3′; β-actin R 5′-GGCCGGACTCATCGTACTCCTGCTT. Reactions were performed in a total volume of 20 μL and gene expression was determined by SYBR Premix Ex Taq TM II (TaKaRa Biotechnology Co., Ltd.) in accordance with the manufacturer’s instructions. Reactions began with a 10 s hot activation of Taq polymerase at 95 °C, followed by 40–45 cycles of amplification in three steps (denaturation at 95 °C for 5 s, 30 s annealing at 60 °C and 30 s extension at 72 °C). The mRNA expression was measured as a ratio to β-actin.
Immunohistochemistry staining
Immunohistochemistry staining was performed to localize Nkx2–5 (1:500, #A12688, wanleibio, China), Gata4 (1:1000, #WL01293, wanleibio, China), BMP4 (1:500, #WL02806, wanleibio, China) and Smad4 (1:300, #WL02049, wanleibio, China). Heart tissues were routinely embedded in paraffin, and sectioned at a 3–5 μ m thickness. All sections were dewaxed, and incubated with 0.3% H2O2 for 10 min. The antigen was repaired by heating 0.03 M citrate buffer (pH 6.0) for 40 min at 95 °C, and then incubated with 5% goat serum albumin for 20 min to block the non-specific binding sites. All sections were incubated with primary antibody overnight at 4 °C in a wet box and stained with sheep anti-rabbit IgG-HRP secondary antibodies (Abcam, UK). The sections were stained using 3,3-diaminobenzidine (DAB) (Sigma, USA) and counterstained with hematoxylin solution. Tissue sections without primary antibodies were the negative controls. Finally, samples were observed and photographed under a microscope.
Western blotting
Western blotting was performed as described previously [14]. Heart samples were lysed in complete RIPA buffer (10 mM Tris-HCl pH 7.4, 150 mM NaCl, 1% NP40, 0.1% sodium dodecyl sulfate, 1 mM phenylmethylsulfonyl fluoride and 1× protease inhibitor cocktail [Roche]) and homogenized using a Sonic Dismembrator 100. The concentration of tissue protein was detected using a BCA protein concentration test kit(#WLA004, wanleibio, China), and equal amounts of soluble protein were separated on 10% polyacrylamide gels, transferred onto a nitrocellulose membrane, and followed by routine western blot analysis. Primary antibody: Nkx2–5 (1:500, #A12688, wanleibio, China), Gata4 (1:1000, #WL01293, wanleibio, China), BMP4 (1:500, #WL02806, wanleibio, China) and Smad4 (1:300, #WL02049, wanleibio, China). Proteins were visualized using a ClarityTM Western ECL Substrate (#WLA003; wanleibio, China) and a Tanon 5200 Full automatic chemiluminescence image analysis system (Tanon Science and Technology Co., Ltd., Shanghai, China).
Statistical analysis
Statistical analysis was performed using SPSS 20.0 statistical software (IBM Corp., Armonk, NY, USA). Variables are expressed as the means ± standard error of the mean (SEM). Data were analyzed using t-tests and one-way ANOVA. Statistical significance was reached at a two-sided p < 0.05.
Results
Effects of L-T4 on serum TSH and TT4 expression in the offspring of SCH pregnant rats
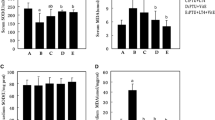
In order to evaluate the thyroid function of SCH pregnant rats, the expression levels of serum TT4 and TSH of all pregnant rats were measured. The serum TSH level in SCH group was significantly higher than that in the Sham group (Fig. 1a, p < 0.05). There was no significant difference in TT4 expression level between the two groups, confirming the successful establishment of the SCH rat model (Fig. 1b). Interestingly, in the LT4-E10 and LT4-E13 groups, L-T4 treatment significantly reduced serum TSH expression level in SCH pregnant rats (Fig. 1a, compared to SCH group, p < 0.05). Moreover, there was no significant difference in the TT4 expression level between SCH, LT4-E10 and LT4-E13 groups. The results indicated that L-T4 treatment would affect the serum TSH expression levels in SCH pregnant rats, but did not change the expression level of TT4.

Effects of L-T4 treatment on the expression levels of serum TSH and TT4 in the offspring of SCH pregnant rats. a TSH expression level of pregnant rats in the Sham, SCH, LT4-E10 (L-T4 treatment started on the 10th day of gestation), and LT4-E13 (L-T4 treatment started on the 13th day of gestation) groups. There were 20 rats in each group. b TT4 expression level of pregnant rats in each group. *p < 0.05 vs Sham group; #p < 0.05 vs SCH group
L-T4 treatment promoted cardiac development in the offspring of SCH pregnant rats
To explore the effect of L-T4 on the heart development of the offspring of SCH pregnant rats, the heart weight and the ratio of heart weight to body (heart/body) weight were examined. The results showed that the heart weight and the heart/body weight of the offspring in the SCH group were significantly lower than that in the Sham group. Furthermore, in the LT4-E10 and LT4-E13 groups, L-T4 treatment significantly increased the heart weight and heart/body weight of the offspring of SCH pregnant rats (Fig. 2, compared to SCH group, p < 0.05). The results suggested that early administration of L-T4 can promote the heart development of the offspring of SCH pregnant rats.

L-T4 promotes cardiac development in the offspring of SCH pregnant rats. a Body weight of the offspring of SCH pregnant rats. b The ratio of heart weight to body weight (Heart/Body Weight) of the offspring of SCH pregnant rats. Results are expressed as the mean ± SEM (n = 5 per group). *p < 0.05 vs Sham group; #p < 0.05 vs SCH group
L-T4 treatment improved the metabolic function of cardiomyocytes in the offspring of SCH pregnant rats
To clarify the effects of L-T4 on the metabolic ability of cardiomyocytes of the offspring of SCH pregnant rats, the activities of SDH, Na+/K+-ATPase and Ca2+-ATPase were detected. The results showed that compared to Sham group, the activities of SDH, Na+/K+-ATPase and Ca2+-ATPase of the offspring were significantly decreased in the SCH group at E16, E18, P5 and P10. In the LT4-E10 and LT4-E13 groups, L-T4 treatment significantly increased the activities of SDH, Na+/K + -ATP and Ca2 + −ATP (Fig. 3, compared to SCH group, p < 0.05). The results suggested that L-T4 treatment regulated the cardiac development of the offspring of SCH pregnant rats by increasing the metabolic function of the cardiomyocytes.

L-T4 treatment improved the metabolic function of cardiomyocytes in the offspring of SCH pregnant rats. (a) Na+/K+-ATPase activity (b) Ca2+-ATPase activity (c) SDH activity. All experiments were repeated at least three times. Data were expressed as the mean ± SEM (n = 5 per group). *p < 0.05 vs Sham group; #p < 0.05 vs SCH group
L-T4 treatment attenuated cardiac tissue injury of the offspring of SCH pregnant rats
The results of histopathological staining showed that the cardiomyocytes of the offspring in the Sham group were orderly arranged, and the structure of cardiomyocytes was clear and intact (Fig. 4). However, the cardiomyocytes of the offspring in the SCH group showed wrinkled, hyperchromatic and loosely arranged structures, with obvious hyperemia, congestion and vacuolar degeneration cell shrinkage, hyperchromatic nuclei, local hyperemia/congestion and vacuolar degeneration. Moreover, in the LT4-E10 and LT4-E13 groups, the degree of cardiomyocyte degeneration, inter-tissue congestion and vacuolar degeneration gradually improved after L-T4 treatment. The results suggested that L-T4 treatment during pregnancy can improve the pathological changes of the heart tissue of the offspring of SCH pregnant rats.

L-T4 attenuated cardiac tissue injury of the offspring of SCH pregnant rats. Representative histopathological images of each group (HE staining, scale bar = 50 μm)
L-T4 promoted the expression of cardiac development proteins in the offspring of SCH pregnant rats
Real-time quantitative PCR, immunohistochemistry and Western blotting were used to detect the expression of mRNAs and proteins related to cardiac development and differentiation of the offspring of SCH pregnant rats. The results showed that the expression levels of Nkx2–5 and Gata4 mRNA of the offspring in the SCH group was significantly lower than that in the Sham group. Furthermore, in the LT4-E10 and LT4-E13 groups, L-T4 treatment significantly increased the expression levels of Nkx2–5 and Gata4 mRNA in the myocardium of the offspring (Fig. 5a-b, compared to SCH group, p < 0.05). Western blotting analysis further confirmed the results that the expression levels of Gata4 and Nkx2–5 protein in the myocardium of the offspring in the SCH group was significantly lower than that in the Sham group, while L-T4 treatment further significantly increased the expression levels of Gata4 and Nkx2–5 protein in the myocardium of the offspring in the LT4-E10 and LT4-E13 groups (Fig. 5c-d, compared to SCH group, p < 0.05). Similarly, the results of immunohistochemical staining showed that Gata4 and Nkx2–5 proteins were mainly expressed in the nucleus of heart cells. Compared to Sham group, the number of Gata4- and Nkx2–5-positive cells of the offspring in the SCH group were significantly reduced. Moreover, L-T4 treatment significantly increased the number of Gata4- and Nkx2–5-positive cells of the offspring in the LT4-E10 and LT4-E13 groups (Fig. 5e-f, compared to SCH group, p < 0.05). The results suggested that L-T4 treatment can regulate the cardiac development of the offspring of SCH pregnant rats by increasing the expression of Gata4 and Nkx2–5 proteins.

L-T4 treatment promoted the expression of cardiac development proteins in the offspring of SCH pregnant rats. a-b Expression levels of Gata4 and Nkx2–5 mRNA in the offspring of SCH pregnant rats; c-d Representative Western blot images and quantitative analysis of Gata4 and Nkx2–5 expression levels; e-f Representative immunohistochemical images of Gata4 and Nkx2–5 expression and quantitative analysis of Gata4- and Nkx2–5-positive cells in each group (Scale bar = 50 μm). All experiments were repeated at least three times. Data were expressed as the mean ± SEM (n = 5 per group). *p < 0.05 vs Sham group; #p < 0.05 vs SCH group
L-T4 treatment increased the expression of BMP4/Smad4 proteins in the offspring of SCH pregnant rats
Real-time quantitative PCR, immunohistochemistry and Western blotting were used to detect the BMP4 and Smad4 mRNA and protein levels of the offspring’s myocardium in SCH pregnant rats. The results showed that the mRNA levels of BMP4 and Smad4 in myocardial tissues of the offspring in the SCH group were significantly lower than that in the Sham group. Furthermore, L-T4 treatment significantly increased the expression of BMP4 and Smad4 mRNA in myocardial tissues of the offspring in the LT4-E10 and LT4-E13 groups (Fig. 6a-b, compared to SCH group, p < 0.05). The results of Western blotting analysis showed that the levels of BMP4 and Smad4 proteins in myocardial tissues of the offspring in the SCH groups were significantly lower than that in the Sham group, while L-T4 treatment significantly increased the expression of BMP4 and Smad4 proteins in both LT4-E10 and LT4-E13 groups (Fig. 6c-d, p < 0.05). Immunohistochemical staining showed that BMP4 and Smad4 were mainly located in the cytoplasm and nucleus of myocardial cells. The number of BMP4- and Smad4-positive myocardial cells in the offspring in SCH group was significantly lower than that in the Sham group. Similarly, L-T4 treatment significantly increased the number of BMP4- and Smad4-positive myocardial cells in the LT4-E10 and LT4-E13 groups (Fig. 6e-f, compared to SCH group, p < 0.05). The results suggest that L-T4 regulates the cardiac development of the offspring in SCH pregnant rats by activating BMP4/ Smad4 signal pathway.

L-T4 treatment increased the expression of BMP4/Smad4 protein in the offspring of SCH pregnant rats. a-b Expression level of BMP4 and Smad4 mRNA in the offspring of SCH pregnant rats; c-d Representative Western blot images and quantitative analysis of BMP4 and Smad4 intensity in each group; e-f) Representative immunohistochemical images of BMP4 and Smad4 expression and quantitative analysis of BMP4- and Smad4- positive cells in each group (Scale bar = 50 μm). All experiments were repeated at least three times. Data were expressed as the mean ± SEM (n = 5 per group). *p < 0.05 vs Sham group; #p < 0.05 vs SCH group
Discussion
Some studies have shown that the offspring of SCH pregnant rats were often accompanied by neurodevelopmental abnormalities, and early intervention with L-T4 can alleviate the neurodevelopmental abnormalities. However, it is not clear whether the offspring of SCH pregnant rats have cardiac developmental abnormalities, and whether L-T4 early intervention can improve the abnormalities of the offspring in SCH pregnant rats. The results of this study revealed that L-T4 treatment significantly decreased the serum TSH expression level in SCH pregnant rats, increased the heart weight and heart/body weight ratio of the offspring in SCH pregnant rats, improved the metabolic function of myocardial cells, and alleviated the pathological changes of myocardial tissues. In addition, L-T4 significantly increased the mRNA and proteins expression of Gata4, Nkx2–5, BMP4, and Smad4 of the offspring in SCH pregnant rats. The results suggest that L-T4 early intervention regulates the cardiac development of the offspring in SCH pregnant rats by activating BMP4/Smad4 signaling pathway, and then increasing the expression of Gata4 and Nkx2–5 proteins.
When pregnant women have hypothyroidism, abnormal thyroid hormone levels can severely affect the development of neuromotor, auditory, cardiovascular, and respiratory systems [15]. Thyroid hormones can promote the transformation of fetal cardiomyocytes from proliferation to hypertrophy and differentiation during full-term and early pregnancy [16]. Since the thyroid function of pregnant women affects the growth and maturation of fetal organs, the birth weight of the baby can indirectly reflect the level of thyroid function of pregnant women [17]. When the concentration of fetal thyroid hormone is kept within a relatively narrow range, it will not affect the normal development of the heart [18].Similarly, the concentration of T4 in umbilical cord blood was positively correlated with the birth weight and height of the baby, because thyroid hormones can provide nutrition and oxygen signals to the fetus in the uterus to regulate fetal growth [19]. Therefore, fetal thyroid hormone is essential for the increase of fetal weight and differentiation of specific cell types at the critical stage of development. Moreover, SCH caused growth retardation of fetal rats in the uterus, and even led to abortion or premature delivery [13] and permanent neurological defects of the offspring [20, 21]. In this study, we found that L-T4 early intervention not only significantly decreased the serum TSH levels in the SCH pregnant rats, but also increased the heart weight and heart/body weight ratio of the offspring, improved the metabolic function of myocardial cells of the offspring, and alleviated the pathological changes of myocardial tissues of the offspring. It was reported that thyroid hormone deficiency affected the expression profiles of myomiR network in the heart of fetal rats and the expression of downstream targeting genes, which in turn led to increased expression of b-MHC and related cardiac dysfunction in adulthood [22]. An infant with ectopic thyroid tissue was found to have sinus bradycardia associated with congenital hypothyroidism [23]. Another study found that SCH was associated with decreased cardiac output in patients with impaired vascular smooth muscle relaxation and reduced nitric oxide level. These changes were related to the decreased expression of sarcoplasmic reticulum Ca2+-ATPase and the inhibition of expression of ATP enzyme phosphoprotein [24]. Low circulating levels of thyroid hormone inhibited the activity of sarcoplasmic reticulum calcium ATP enzyme, which controlled the contraction and relaxation cycle through ATPase, leading to myocardial stiffness and eventually left ventricular diastolic dysfunction [25]. In addition, L-T4 treatment significantly improved low density lipoprotein, total cholesterol, hypertension, diastolic dysfunction and delayed arteriosclerosis in patients with SCH [26]. These results suggest that L-T4 treatment can improve the cardiac development of the offspring in SCH pregnant rats, which was related to increasing the heart weight, improving the metabolic function of the cardiomyocytes and alleviating the pathological changes of cardiomyocytes.
During the early development of embryonic heart, many genes such as bone morphogenetic proteins (BMPs), fibroblast growth factor and activin/nodule are involved in heart development and differentiation [27, 28]. BMP signaling pathway played a central role in the induction of mesoderm and heart development. The BMP ligand binds to type II receptor and then activates type I receptor to phosphorylate Smad1, Smad5 or Smad8 signal transduction pathway regulated by BMP receptor [29]. When BMP is released from the receptor complex, the phosphorylated R-Smad binds to the common Smad4 to form a trimeric complex composed of two R-Smad and Smad4, and then induces the transcription of downstream genes [30]. BMP4 knockout Fetal mice died within 9.5 days after birth, and most of them had little or no mesoderm differentiation [31]. BMP4 can induce the expression of Nkx2–5 and Gata4 in cardiac progenitor cells, which is necessary for cardiac development and differentiation [32]. Gata4 is one of the key transcription factors in fetal heart development. Abnormal expression of Gata4 can cause various fetal heart malformations, such as septal defect, tetralogy of Fallot, myocardial trabecular dysplasia, and valve malformation [33,34,35]. In this study, our results showed that the mRNA and protein expression levels of Gata4, Nkx2–5, BMP4, and Smad4 of the offspring were significantly decreased in SCH pregnant rats. Remarkably, L-T4 early intervention not only significantly increased the mRNA and protein levels of Gata4 and Nkx2–5, but also activated the BMP4/Smad4 signaling pathway in the myocardium of the offspring of SCH pregnant rats. It was reported that exposure of pregnant rats to di (2-ethylhexyl) phthalate caused cardiac malformation in the offspring, which may be related to the inhibition of cardiac Gata4/mef2c/ch expression [36]. Similarly, the potential toxicity of PM2.5 to fetal heart tissue was related to the down-regulation of transcriptional factors Gata4 and Nkx2–5, which were functionally associated with to embryonic heart development and abnormalities of fetal heart structure and function [37]. Moreover, exposure of pregnant rats to PM2.5 aggravated cardiovascular dysplasia caused by homocysteine in their offspring [38]. These results indicate that early treatment of L-T4 can significantly improve the myocardial development of the offspring of SCH pregnant rats, which may be related to the activation of BMP4/Smad4 and the regulation of Gata4 and Nkx2–5 protein expression.
Conclusion
In conclusion, early administration of L-T4 can regulate the cardiac development of the offspring of SCH pregnant rats by activating BMP4/Smad4 signal pathway, and then increasing the expression of Gata4 and Nkx2–5.
Availability of data and materials
The datasets generated and analysed during the current study are not publicly available due the principle of confidentiality of funding, but are available from the corresponding author on reasonable request.
Abbreviations
- SCH:
-
Subclinical hypothyroidism
- PPARγ:
-
Peroxisome proliferator-activated receptor gamma
- LT4-E10 group:
-
L-T4 treatment started on the 10th day of gestation
- SDH:
-
Succinate dehydrogenase
- T4:
-
Thyroid hormone thyroxine
- TSH:
-
Thyroid-stimulating hormone
- BMPs:
-
Bone morphogenetic proteins
References
Wu T, Flowers JW, Tudiver F, Wilson JL, Punyasavatsut N. Subclinical thyroid disorders and cognitive performance among adolescents in the United States. BMC Pediatr. 2006;6:12.
Teng W, Shan Z, Patil-Sisodia K, Cooper DS. Hypothyroidism in pregnancy. Lancet Diabetes Endocrinol. 2013;1(3):228–37.
Chan S, Boelaert K. Optimal management of hypothyroidism, hypothyroxinaemia and euthyroid TPO antibody positivity preconception and in pregnancy. Clin Endocrinol. 2015;82(3):313–26.
Santos OC, Silva NA, Vaisman M, Turano MD, Dytz MG, Huber GA, Braulio VB, Teixeira PF. Evaluation of epicardial fat tissue thickness as a marker of cardiovascular risk in patients with subclinical hypothyroidism. J Endocrinol Investig. 2015;38(4):421–7.
Biondi B, Cappola AR, Cooper DS. Subclinical hypothyroidism: a review. JAMA. 2019;322(2):153–60.
Bielecka-Dabrowa A, Godoy B, Suzuki T, Banach M, von Haehling S. Subclinical hypothyroidism and the development of heart failure: an overview of risk and effects on cardiac function. Clin Res Cardiol. 2019;108(3):225–33.
Cappola AR, Desai AS, Medici M, Cooper LS, Egan D, Sopko G, Fishman GI, Goldman S, Cooper DS, Mora S. Thyroid and cardiovascular disease research agenda for enhancing knowledge, prevention, and treatment. Circulation. 2019;13:10.
Tayal B, Graff C, Selmer C, Kragholm KH, Kihlstrom M, Nielsen JB, Olsen AS, Pietersen AH, Holst AG, Sogaard P. Thyroid dysfunction and electrocardiographic changes in subjects without arrhythmias: a cross-sectional study of primary healthcare subjects from Copenhagen. BMJ Open. 2019;9(6):e023854.
Alexander EK, Marqusee E, Lawrence J, Jarolim P, Fischer GA, Larsen PR. Timing and magnitude of increases in levothyroxine requirements during pregnancy in women with hypothyroidism. N Engl J Med. 2004;351(3):241–9.
Cerbone M, Capalbo D, Wasniewska M, Mattace Raso G, Alfano S, Meli R, De Luca F, Salerno M. Cardiovascular risk factors in children with long-standing untreated idiopathic subclinical hypothyroidism. J Clin Endocrinol Metab. 2014;99(8):2697–703.
Nakova VV, Krstevska B, Kostovska ES, Vaskova O, Ismail LG. The effect of levothyroxine treatment on left ventricular function in subclinical hypothyroidism. Arch Endocrinol Metab. 2018;62(4):392–8.
Arslan D, Buyukinan M, Uysal C, Deniz CD. Evaluation of cardiovascular risk by growth-differentiation factor-15 and tissue Doppler imaging in children with subclinical hypothyroidism. Endocrine. 2019;65(3):601–7.
Shan L, Zhou Y, Peng S, Wang X, Shan Z, Teng W. Implantation failure in rats with subclinical hypothyroidism is associated with LIF/STAT3 signaling. Endocr Connect. 2019;8(6):718–27.
Ansari MY, Ahmad N, Haqqi TM. Butein activates autophagy through AMPK/TSC2/ULK1/mTOR pathway to inhibit IL-6 expression in IL-1beta stimulated human chondrocytes. Cell Physiol Biochem. 2018;49(3):932–46.
Forhead AJ, Fowden AL. Thyroid hormones in fetal growth and prepartum maturation. J Endocrinol. 2014;221(3):R87–R103.
Chattergoon NN. Thyroid hormone signaling and consequences for cardiac development. J Endocrinol. 2019;242(1):T145–60.
Thornburg K, Jonker S, O'Tierney P, Chattergoon N, Louey S, Faber J, Giraud G. Regulation of the cardiomyocyte population in the developing heart. Prog Biophys Mol Biol. 2011;106(1):289–99.
Miller TA. Growth in congenital heart disease: outcome or predictor? J Am Heart Assoc. 2018;7(17):e010262.
Ribeiro LGR, Silva JF, Ocarino NM, Souza CA, Melo EG, Serakides R. Excess maternal thyroxine alters the proliferative activity and angiogenic profile of growth cartilage of rats at birth and weaning. Cartilage. 2018;9(1):89–103.
Zhang F, Chen J, Lin X, Peng S, Yu X, Shan Z, Teng W. Subclinical hypothyroidism in pregnant rats impaired learning and memory of their offspring by promoting the p75(NTR) signal pathway. Endocr Connect. 2018;7(5):688–97.
Ge JF, Xu YY, Qin G, Cheng JQ, Chen FH. Resveratrol ameliorates the anxiety- and depression-like behavior of subclinical hypothyroidism rat: possible involvement of the HPT Axis, HPA axis, and Wnt/beta-Catenin pathway. Front Endocrinol (Lausanne). 2016;7:44.
Yousefzadeh N, Jeddi S, Ghiasi R, Alipour MR. Effect of fetal hypothyroidism on MyomiR network and its target gene expression profiles in heart of offspring rats. Mol Cell Biochem. 2017;436(1–2):179–87.
Nakanomori A, Nagano N, Seimiya A, Okahashi A, Morioka I. Fetal sinus bradycardia is associated with congenital hypothyroidism: an infant with ectopic thyroid tissue. Tohoku J Exp Med. 2019;248(4):307–11.
Udovcic M, Pena RH, Patham B, Tabatabai L, Kansara A. Hypothyroidism and the heart. Methodist Debakey Cardiovasc J. 2017;13(2):55–9.
Rodondi N, Bauer DC, Cappola AR, Cornuz J, Robbins J, Fried LP, Ladenson PW, Vittinghoff E, Gottdiener JS, Newman AB. Subclinical thyroid dysfunction, cardiac function, and the risk of heart failure. J Am Coll Cardiol. 2008;52(14):1152–9.
Peeters RP. Subclinical hypothyroidism. N Engl J Med. 2017;376(26):2556–65.
Liu W, Deng Z, Zeng Z, Fan J, Feng Y, Wang X, Cao D, Zhang B, Yang L, Liu B. Highly expressed BMP9/GDF2 in postnatal mouse liver and lungs may account for its pleiotropic effects on stem cell differentiation, angiogenesis, tumor growth and metabolism. Genes Dis. 2020;7(2):235–44.
Zhou C, Yan Q, Zou QY, Zhong XQ, Tyler CT, Magness RR, Bird IM, Zheng J. Sexual dimorphisms of preeclampsia-dysregulated transcriptomic profiles and cell function in fetal endothelial cells. Hypertension. 2019;74(1):154–63.
Chang H, Huylebroeck D, Verschueren K, Guo Q, Matzuk MM, Zwijsen A. Smad5 knockout mice die at mid-gestation due to multiple embryonic and extraembryonic defects. Development. 1999;126(8):1631–42.
Garside VC, Chang AC, Karsan A, Hoodless PA. Co-ordinating Notch, BMP, and TGF-β signaling during heart valve development. Cell Mol Life Sci. 2013;70(16):2899–917.
Wang J, Greene SB, Martin JF. BMP signaling in congenital heart disease: new developments and future directions. Birth Defects Res A Clin Mol Teratol. 2011;91(6):441–8.
Takei S, Ichikawa H, Johkura K, Mogi A, No H, Yoshie S, Tomotsune D, Sasaki K. Bone morphogenetic protein-4 promotes induction of cardiomyocytes from human embryonic stem cells in serum-based embryoid body development. Am J Physiol Heart Circ Physiol. 2009;296(6):H1793–803.
Kardon G, Ackerman KG, McCulley DJ, Shen Y, Wynn J, Shang L, Bogenschutz E, Sun X, Chung WK. Congenital diaphragmatic hernias: from genes to mechanisms to therapies. Dis Model Mech. 2017;10(8):955–70.
Yuan SM. Fetal arrhythmias: genetic background and clinical implications. Pediatr Cardiol. 2019;40(2):247–56.
He A, Gu F, Hu Y, Ma Q, Ye LY, Akiyama JA, Visel A, Pennacchio LA, Pu WT. Dynamic GATA4 enhancers shape the chromatin landscape central to heart development and disease. Nat Commun. 2014;5:4907.
Tang C, Deng Y, Duan H, Zhang Y, Li Y, Qiu D, Zhou K, Hua Y, Wang C. The effect of maternal exposure to di-(2-ethylhexyl)-phthalate on fetal cardiac development in mice. J Appl Toxicol. 2018;38(6):834–42.
Carter DR, Buckle AD, Tanaka K, Perdomo J, Chong BH. Art27 interacts with GATA4, FOG2 and NKX2.5 and is a novel co-repressor of cardiac genes. PLoS One. 2014;9(4):e95253.
Chen H, Chen X, Hong X, Liu C, Huang H, Wang Q, Chen S, Chen H, Yang K, Sun Q. Maternal exposure to ambient PM2.5 exaggerates fetal cardiovascular maldevelopment induced by homocysteine in rats. Environ Toxicol. 2017;32(3):877–89.
Acknowledgments
Not applicable.
Funding
The study was supported by the PLA foundation of Key issues of logistical support department of the Central Military Commission (BWS16J010). The funding sources had no involvement in study design; data collection, analysis and interpretation; or report writing.
Author information
Authors and Affiliations
Contributions
DX and JY designed experiments, and wrote the manuscript. JLS revised and polished the manuscript. DX completed animal experiment, pathological experiment, molecular biology experiment and protein experiment. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study was approved by the Animal Care and Use Committee of the Northern Theater General Hospital.
Consent for publication
Not applicable.
Competing interests
The authors declare that there are no conflicts of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Xue, D., Sun, J.L. & Yang, J. Early L-T4 intervention improves fetal heart development in pregnant rats with subclinical hypothyroidism rats by activating BMP4/Smad4 signaling pathway. BMC Cardiovasc Disord 20, 369 (2020). https://doi.org/10.1186/s12872-020-01646-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12872-020-01646-3