
Abstract
Background
Ready-to-eat (RTE) vegetables have become increasingly popular along with the trend of moving towards a healthy lifestyle. However, RTE vegetables are at a higher risk of containing pathogens, maybe owing to lack of rigorous sanitization procedures. To understand the prevalence and potential risk of Listeria monocytogenes in RTE vegetables, we investigated the contamination level and characteristics of L. monocytogenes isolated from fresh vegetables.
Results
Twenty-three (5.49%) of the 419 vegetables samples were positive for L. monocytogenes. Phylogenetic group I.1 (1/2a-3a) and II.2 (1/2b-3b-7) strains were predominant in 30 isolates, which accounted for 33.3 and 50.0%, respectively. Multilocus sequence typing of the 30 isolates grouped them into nine sequence types (STs). The most common STs were ST87 (36.7%) and ST8 (26.7%). Virulence analysis showed that all 30 isolates harbored eight classical virulence genes, 10.0% isolates harbored the llsX gene (ST3 and ST1 strains), and 36.7% carried the ptsA gene and belonged to ST87. Approximately 83.3% isolates carried full-length inlA, whereas five isolates had premature stop codons in inlA, three of which belonged to ST9 and two to ST8. Antibiotic susceptibility showed the isolates were varyingly resistant to 13 antibiotics, 26.7% of the isolates were multi-drug resistant.
Conclusions
The fresh vegetables contain some potential hypervirulent L. monocytogenes (ST1 and ST87) in the Chinese markets. In addition, the high rate of L. monocytogenes isolates was multi-drug resistant. Fresh raw vegetables may be a possible transmission route for L. monocytogenes infection in consumers. Therefore, sanitization of raw fresh vegetables should be strengthened to ensure their microbiological safety when used as RTE vegetables.
Similar content being viewed by others

Background
In recent years, there is an increasing demand for ready-to-eat (RTE) vegetables because people have realized the importance of a healthy diet and healthy lifestyle. However, the increasing consumption of fresh produce, including fruits and vegetables was recognized as a source of microbiological foodborne outbreaks in many parts of the world [1, 2].
Listeria monocytogenes is a common Gram-positive facultative anaerobic bacillus that is recognized as one of four foodborne pathogens by the World Health Organization. It can invasively infect humans and animals, causing severe listeriosis leading to meningoencephalitis, sepsis and fetal infection or miscarriage in pregnant women, with a mortality rate of 20–30% [3]. High-risk groups include pregnant women, infants, immunodeficient patients and the elderly [3]. Fresh vegetables and herbs, such as coriander, lettuce, tomato, cucumber, cabbage, carrot, and shallot, are generally used as raw material in RTE vegetables after minimal processing to preserve nutrients, which is an important source of RTE vegetable pollution. In recent years, reports show an increase in the prevalence of L. monocytogenes in fresh and RTE vegetables. A study revealed that L. monocytogenes was detected in 6.67% fruit and vegetable products from 2011 to 2015 in Jilin Province, China (Yang et al., 2017). Both in Europe and USA, a link between foodborne disease outbreaks and pathogen (such as L. monocytogenes) contaminated green leafy vegetables, such as lettuce and spinach, and their RTE salads has been reported [4]. In recent years, the occurrence of L. monocytogenes in vegetables has been reported in several countries, with the detection rate ranging from 0.6 to 9.1% [5,6,7,8]. China is the largest vegetable producer and consumer in the world, with an output volume of vegetables and processed products of 10.1 million tons in 2015 [9], However, there is no uniform method for risk assessment and management of L. monocytogenes in RTE vegetables [10],which are mostly consumed raw or after minimal processing and pathogen contamination poses a health risk.
It is critical to evaluate the virulence of L. monocytogenes in fresh vegetables. This study aimed to evaluate the qualitative and quantitative contamination level by L. monocytogenes in fresh vegetables from Chinese markets. We determined the phylogenetic group, virulence profiles, and antimicrobial resistance and performed multilocus sequence typing (MLST) to comprehensively study the virulence of L. monocytogenes isolated from fresh vegetables across China.
Results
Qualitative and quantitative analysis of L. monocytogenes in fresh vegetable samples
A total of 23 (5.49%) positive samples were detected by quantitative and qualitative methods in the 419 collected vegetable samples. According to the different types of vegetables collected, the positive rate of lettuce was highest (7.78%), followed by coriander (4.49%), tomato (4.90%) and cucumber (4.88%) (Table 1). Among the 23 positive samples, quantitative detection by MPN method showed that the contamination level of two samples corresponding to lettuce and tomato were up to 110 MPN/g, one tomato sample was 24 MPN/g, and the remaining 20 samples were between 0.3–10 MPN/g. We also isolated 30 strains of L. monocytogenes from the 23 positive samples (Additional file 1: Table S2).
Phylogenetic group analysis
Multiple PCR was used to analyze the phylogenetic group of 30 isolates obtained from 23 L. monocytogenes-positive samples (Additional file 1: Figure S1). Isolates belonging to different phylogenetic group from the same sample were included in this study. Isolates were divided into four phylogenetic groups, among which phylogenetic group I.1 (1/2a-3a) accounted for 33.3% (10/30), phylogenetic group I.2 (1/2c-3c) accounted for 13.3% (4/30), phylogenetic group II.1 (4b-4d-4e) accounted for 3.3% (1/30), and phylogenetic group II.2 (1/2b-3b-7) was predominant at 50.0% (15/30) (Fig. 1). L. monocytogenes of two different phylogenetic groups were isolated from three lettuce samples, one cucumber sample and one coriander sample, suggesting that there were different serotypes of L. monocytogenes contaminating the same vegetable sample.

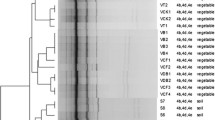
Characteristics of 30 L. monocytogenes isolated from fresh vegetables. Of the 10 virulence genes (prfA, mpl, plcA, inlB, plcA, hly, iap, actA, llsX and ptsA), black squares indicate the presence of the corresponding gene; white squares represent lack of corresponding gene. (a) PMSC, premature stop codons in inlA; black squares indicate the presence of full-length inlA. (b) K (k), kanamycin; CN (cn), gentamycin; S (s), streptomycin; LEV (lev), levofloxacin; CIP (cip), ciprofloxacin; SAM (sam), sulbactam/ampicillin; E (e), erythromycin; RD (rd), rifampin; DA (da), clindamycin; TE (te), tetracycline; DO (do), doxycycline; VA (va), vancomycin; C (c), chloramphenicol; − indicates no resistance. Antibiotic abbreviations in uppercase indicateresistance, while those in lowercase indicate intermediate resistance
Multilocus sequence typing analysis
According to the sequencing results of the seven housekeeping genes using MLST, the sequence types (STs) and clonal complexes (CCs) of the isolates were obtained. The 30 isolates were classified into nine STs belonging to nine CCs (Fig. 1). ST87/CC87 and ST8/CC8 were predominant and accounted for 36.7% (11/30) and 26.7% (8/30), respectively. CC9 accounted for 13.3% (4/30), CC3 for 6.7% (2/30), and CC1, CC5, CC7, CC59 and CC155 were 3.3% (1/30) each, respectively.
Virulence profiles analysis
In this study, a total of 10 virulence genes (prfA, mpl, plcA, inlB, plcA, hly, iap, actA, llsX and ptsA) of 30 isolates were detected by PCR. The results showed that except for the llsX and ptsA, all isolates carried the other eight virulence genes. Only three isolates (413-2LM, 1965-1LM and 2013-1LM) carried the llsX gene, which belonged to ST1 and ST3. A total of 11 (36.7%) isolates harbored the ptsA gene and all belonged to ST87 (Additional file 1: Figure S2). Analysis of inlA DNA sequencing showed that five isolates harbored PMSCs in the inlA gene. Four isolates (2114-1LM, 2114-3LM, 2363-1LM and 2363-3LM) had a deletion of an adenine at position 1637 of inlA [11], and one isolate (3015-1LM) had a deletion of an adenine at position 12 [12], These frame shift mutations lead to the creation of a nonsense codon, TAA, at position 1729 and 25, respectively. Moreover, three isolates (2114-1LM, 2363-1LM, and 2363-3LM) belonged to ST9, and the other two (3015-1LM and 2114-3LM) belonged to ST8.
Antimicrobial resistance analysis
According to CLSI guidelines and MIC method, the antimicrobial resistance analysis of 16 antibiotic agents against the L. monocytogenes isolates is shown in Table 2 and Fig. 1. Among the 30 L. monocytogenes isolates, all strains were susceptible to three antibiotics, penicillin, ampicillin, and sulfamethoxazole/trimethoprim. In contrast, resistance to clindamycin was the most common resistance and was identified in 63.3% isolates. On the other hand, only four (13.3%) isolates (133-1LM, 413-2LM, 413-4LM, and 1741-1LM) were susceptible to all the 16 antibiotics. Other isolates showed varying degrees of resistance and/or intermediate resistance to individual antibiotics. It is worth mentioning that 26.7% isolates were defined as multi-drug resistant, of which 1331-1LM and 1331-2LM were isolated from the same tomato sample and showed resistant/intermediate resistant to 12 and 9 antibiotics, respectively.
Discussion
There is no doubt that L. monocytogenes is a critical foodborne pathogen worldwide. At present, there are very little data on the identification of L. monocytogenes contamination in fresh vegetables in China, especially for quantitative analysis, which limits risk assessment and the development of relevant safety standards for fresh vegetables used in RTE vegetables. In this study, 419 commercially available fresh vegetables in 43 representative cities/regions in China were investigated, of which 23 (5.49%) were positive for L. monocytogenes (Table 1). The highest contamination rate (7.78%) was found in lettuce sample and the MPN value of one positive sample exceeded 100 MPN/g. This could be due to the relatively large leaf surface of lettuce that is close to the soil surface and therefore easily contaminated by soil and environmental water. Wang et al. analyzed 153 RTE vegetable samples from six districts in Zigong City, Sichuan Province of China, and found a L. monocytogenes positive rate of 6.5% [13], which was consistent with our results. Kuanet al. reported that L. monocytogenes was more frequently observed in organic (9.1%) than in conventional (2.7%) vegetables in Malaysia [6]. These studies have reported different degrees of L. monocytogenes contamination in RTE vegetables. To date, National Health Commission of P. R. China has formulated a zero-tolerance policy for L. monocytogenes in cooked meat and ready-to-eat fresh meat. European Union has formulated a < 100 CFU/g policy for ready-to-eat foods during their shelf-life. However, there is no specific quantitative rule for the presence of L. monocytogenes in ready-to-eat vegetables. Although the prevalence of L. monocytogenes on fresh vegetables is low, RTE vegetables are generally not treated at high temperature, and if the raw material is contaminated, it poses a major potential transmission route of L. monocytogenes infection. It is necessary to perform the risk assessment of L. monocytogenes in vegetables for providing basic data to formulate the quantitative rule.
L. monocytogenes was divided into 13 serotypes, which were further divided into five phylogenetic groups by multiplex PCR [14]. In this study, L. monocytogenes phylogenetic group I.1 (33.3%) and II.2 (50.0%) were predominant. Previous studies have shown that phylogenetic group I.1 (1/2a-3a) of the food isolate was mainly serotype 1/2a, and phylogenetic group II.2 (1/2b-3b-7) was primarily serotype 1/2b [14]. This indicated that phylogenetic groups I.1 and II.2 of the isolates predominate in fresh vegetables, which is consistent with previous studies [15]. Of the 13 serotypes, more than 95% of disease and food contamination were associated with 4 serotypes (1/2a, 1/2b, 1/2c, and 4b), and approximately 50% of clinical cases were of serotype 4b, followed by 1/2a (27%). The vast majority of L. monocytogenes serotypes isolated from fresh vegetables were consistent with the serotypes of clinical pathogenic and food contaminating strains, suggesting that contamination of fresh vegetables used to make RTE vegetables may be a transmission route for L. monocytogenes infection.
Previous studies have shown that L. monocytogenes was a highly heterogeneous species with regards to pathogenicity, which consisted of different virulence clones, and clones CC1, CC2, CC4 and CC6 were strongly associated with a clinical origin. Especially, CC4 strain carried LIPI-4, a locus involves in neural and placental infection, is considered as high pathogenicity [16]. We analyzed the MLST data of 30 strains that were collected from fresh vegetables in China and observed that the frequency distribution of these clones was highly uneven (Fig. 1). The most prevalent clones were CC87 (36.7%), CC8 (26.7%), CC9, (13.3%), CC3 (6.7%), and CC1, CC5, CC7, CC59, and CC155 at 3.3%. In western countries, ST121, ST9, and ST8 are predominant in food items and food associated environments [16]. Unlike western countries, CC87 is predominant in vegetable samples in this study, which is also persistent in pre-packed smoked salmon in Singapore, Asia [17]. The PMSCs in inlA are mainly found in ST9 in this study, which was consistent with the results of ST121, indicating that the attenuated virulence of predominant L. monocytogenes STs may occur in food items and food associated environments. In addition, Wang et al. analyzed the CCs of 33 L. monocytogenes clinical isolates from different regions of China in the past decade, and showed that CC87 (24.2%) and CC8 (9.09%) strains were predominant [13]. These results demonstrate that L. monocytogenes isolated from fresh vegetables might have potentially hypervirulent. To elucidate the status of virulence genes in the isolated strains, we performed full-length sequencing of inlA and PCR amplification of ten other virulence genes. The results suggest that only five isolates harbored PMSCs in the inlA gene and all isolates harbored eight virulence genes (prfA, mpl, plcA, inlB, plcA, hlyA, iap and actA). Particularly, the llsX gene (encoding Listeriolysin, LLS, a hemolytic and cytotoxic factor) belonging to LIPI-3, has been greatly associated with a subset of lineage I in human listeriosis [18]. LLS acts to target the host gut microbiota, responsible for the majority of listeriosis outbreaks [19]. Four isolates (413-2LM, 1965-1LM and 2013-1LM) carried the llsX gene, and belonged to CC1 and CC3, indicating the presence of Listeria pathogenicity islands 3 (LIPI-3) in the genome, which enhances hemolytic and cytotoxic activity of L. monocytogenes [18]. Recent study reported that phosphotransferase system (PTS) of the cellobiose family mediates L. monocytogenes neural and placental tropisms, which consists of six genes (including the ptsA gene) [16]. A total of 11 isolates analyzed in this study carried LIPI-4 and belonged to ST87. Wang et al. reported that ST87 is the predominant clinical isolate of listeriosis in China, which is different from human infection cases in the United States and Europe [20]. In conclusion, although the L. monocytogenes contamination rate in fresh vegetables was low, their isolates contained phenotypes, which frequently results in listeriosis for people, including ST87, ST8 and potential high-virulence strains carrying LIPI-3. Furthermore, 83.3% isolates carried the full-length inlA, suggesting that fresh vegetables could become the transmission route of L. monocytogenes infection.
At present, the drug resistance of food-borne pathogens is a growing concern that threatens public health. Antibiotics commonly administered to treat listeriosis include penicillin, amoxicillin, ampicillin, meropenem, vancomycin, gentamicin, rifampin, cotrimoxazole, levofloxacin, and linezolid, with the first three used with the highest frequency [21]. However, L. monocytogenes isolates with varying levels of resistance to antibiotics has been reported. Escolar et al. found Listeria spp. isolated from RTE products of animal origin to have clindamycin (100%), ciprofloxacin (52%), penicillin (32%) and ampicillin (20%) resistance in Spain [22]. None of isolates was found to be resistant to ampicillin and penicillin in this study, suggesting that the first-choice drugs are still effective for listeriosis treatment. The results of this study suggest the presence of a high level of resistance to clindamycin (63.3%, see Table 2), which is used in hospital treatments [23]. On the other hand, 26.7% isolates were found to be multi-drug resistant in our study, with two strains (1331-1LM and 1331-2LM) isolated from the same tomato sample showing 110 MPN/g and being tolerant to 12 and 9 antibiotics, respectively. These results prompt the necessity of novel alternatives for antimicrobial to disrupt the emerging multidrug resistance of L. monocytogenes. The acquisition of multi-drug resistance is possibly related to the widespread use of antibiotics. At the same time, the bacteria can develop resistance mechanisms or acquire resistance by transmission of genetic material from other bacterial species [22]. Therefore, it is necessary and significant to further study the possible mechanism of resistance, especially the antimicrobial resistance mechanisms of L. monocytogenes in fresh vegetables.
Conclusion
In conclusion, 5.49% of fresh vegetables collected from the 43 representative cities/regions markets in China were positive for L. monocytogenes. Phylogenetic group I.1 (33.3%) and II.2 (50.0%) of L. monocytogenes were dominant in fresh vegetable sources. The strains of ST87 (36.7%) and ST8 (26.7%) were predominantly identified by MLST analysis. The virulence genes prfA, mpl, plcA, inlB, plcA, hlyA, iap and actA were carried by all strains, in contrast 10.0% carried the llsX gene and 36.7% carried the ptsA gene. Moreover, 83.3% of the isolates were full-length for inlA, indicating that most isolates were capable of invading the host cells. Except for sulfamethoxazole, ampicillin, and penicillin, different degrees of resistance to the other 13 antibiotics was observed, and multi-drug resistant strains accounted for 26.7%. The results suggest that fresh raw vegetables may be a possible transmission vehicle for L. monocytogenes infection in consumers, and should be treated with rigorous sanitization treatments when used for RTE vegetable to ensure microbiological safety.
Methods
Sample collection
A total of 419 fresh vegetables, which form the main raw ingredients in RTE vegetables, were randomly purchased from 43 representative cities or regions of China (Additional file 1: Table S1). The collected samples included coriander (n = 89), lettuce (n = 90), tomato (n = 102), cucumber (n = 123), and other vegetables (n = 15). Sampling was carried out over a 5-year period (July 2011 to July 2016). After collection, all samples were immediately placed in sterile bags, kept in an insulated box with ice packs, and transported to the laboratory for analysis within 2 h after reaching the laboratory.
Qualitative and quantitative analysis
Qualitative analysis was carried out according to the food microbiological test of L. monocytogenes in GB4789.40–2010 (National Food Safety Standards of China) with slight modification. In brief, samples were cut and mixed on a sterile workbench, and 25 g of each sample was homogenized in 225 mL Listeria enrichment broth 1 (LB1; Guangdong Huankai Co. Ltd., Guangzhou, China). Homogenates were cultured at 30 °C for 24 h, after which 0.1 mL LB1 enriched cultures were transferred to 10 mL of LB2 and incubated at 30 °C for 24 h. After incubation, the collected samples were streaked on Chromagar Listeria plates (Guangdong Huankai Co. Ltd., China) with an inoculation loop and incubated at 37 °C for 48 h. Three to five colonies typically blue in color with a white halo were selected for identification of L. monocytogenes using the Microgen ID Listeria identification system (Microgen, Camberley, United Kingdom) according to manufacturer’s instructions. The L. monocytogenes isolates were stored with 20% glycerol at − 40 °C for further use.
Quantitative detection of L. monocytogenes was performed using the operational flow of the most probable number (MPN) method [24]. 25 g of each sample was added to 225 mL of Frasher enrichment broth (Guangdong Huankai Co. Ltd., China) in a homogenization bag. The sample was shaken evenly, before transferring 10 mL (1 g), 1 mL (0.1 g), or 0.1 mL (0.01 g) of the homogenates to sterile tubes. To the latter two tubes, 10 mL Fraser broth was also added, and each dilution was prepared in triplicate [25]. After incubating at 30 °C for 48 h, the darkened Fraser tubes were selected and streaked onto Listeria chromogenic agar plates. After further purification, the isolates were identified using the Microgen ID Listeria identification system and multiplex PCR, and the MPN was calculated by referring to the MPN table.
Phylogenetic group analysis
The L. monocytogenes isolates of fresh vegetables were identified for phylogenetic groups by multiplex PCR according to the method of Doumith et al. The multiplex PCR method can classify 13 serotypes of L. monocytogenes into five phylogenetic groups, which are designated as I.1 (1/2a-3a), I.2 (1/2c-3c), II.1 (4b-4d-4e), II.2 (1/2b-3b-7) and III (4a-4c). The PCR products were separated by electrophoresis on a 1.5% agarose gel (containing 0.005% Goldview stain) at 100 V for 40 min, photographed using Tanon 2500 UV camera system. Images were saved in TIFF format for further analysis.
Multilocus sequence typing (MLST) analysis
According to the analytical method established by Ragon et al. [26], the specific primers of seven housekeeping genes (abcZ, bglA, cat, dapE, dat, ldh and lhkA) were used for MLST using PCR amplification on isolated strains. The ldh primers were ldhF: 5′–GACAGAACAATTGGGGATGCAATG–3′ and ldhR: 5′–AACGCCGTAGAATGTAGCGCCT–3′ [27]. The all annealing temperatures are 52 °C (45 °C for bglA and 58 °C for ldh). The amplified products were subjected to bidirectional sequencing, and MLST analysis was performed using the Listeria MLST database (http://bigsdb.pasteur.fr/listeria/listeria.html) curated by the Pasteur Institute of France. Based on MLST’s seven housekeeping genes (abcZ, bglA, cat, dapE, dat, ldh and lhkA) linkage sequences, the Neighbor-Joining phylogenetic tree was constructed using MEGA 7.0.
Virulence profiles analysis
PCR amplification was performed to detect potential virulence-related genes of 30 L. monocytogenes isolates from fresh vegetable sources. The 10 pairs of primers and reaction conditions are shown respectively in Table 3. Multiple naturally occurring mutations leading to a premature stop codon (PMSC) in inlA have been reported worldwide, and these mutations were causally associated with attenuated virulence [28, 29]. Therefore, the inlA gene was investigated by amplicon sequencing to determine the presence of PMSCs. The full-length inlA gene (2403 bp) was amplified from 30 isolates using external primers, and internal primers were used for sequencing [30]. The inlA sequences were assembled with MEGA software (version7.0.26). By comparing to the complete inlA sequence of the L. monocytogenes EGDe reference strain, sites of PMSC mutations in inlA were determined [31].
Antimicrobial resistance analysis
According to the criteria of Staphylococcus or Enterococcus susceptibility test of the Clinical and Laboratory Standards Institute (CLSI) guidelines (M100-27th ed.) [32], all strains were analyzed by the disk diffusion method. A total of 14 antibiotic agents (classified in 10 categories) were tested at specific concentrations per disk. Details are provided in Table 2. Staphylococcus aureus ATCC 25923 and Escherichia coli ATCC 25922 were used as quality control strains. Particularly, ampicillin and penicillin were tested by minimal inhibitory concentration (MIC) method. Isolates showing growth in wells with a concentration more than or equal to 2 μg/mL were considered resistant strains according to Standards Institute guidelines [33]. Isolates which resistant to three or more types of antibiotic resistance were defined as multidrug-resistant strains [34].
Abbreviations
- CCs:
-
Clonal complexes
- CLSI:
-
Clinical and Laboratory Standards Institute
- L. monocytogenes :
-
Listeria monocytogenes
- LB1:
-
Listeria enrichment broth 1
- LB2:
-
Listeria enrichment broth 2
- MIC:
-
Minimal inhibitory concentration
- MLST:
-
Multilocus sequence typing
- MPN:
-
Most probable number
- PMSC:
-
Premature stop codon
- RTE:
-
Ready-to-eat
- STs:
-
Sequence types
References
Alegbeleye OO, Singleton I, Sant'Ana AS. Sources and contamination routes of microbial pathogens to fresh produce during field cultivation: a review. Food Microbiol. 2018;73:177–208.
Wadamori Y, Gooneratne R, Hussain MA. Outbreaks and factors influencing microbiological contamination of fresh produce. Journal of the Science of Food & Agriculture. 2017;97(5):1396–403.
Radoshevich L, Cossart P. Listeria monocytogenes: towards a complete picture of its physiology and pathogenesis. Nat Rev Microbiol. 2018;16(1):32–46.
Taban BM, Halkman AK. Do leafy green vegetables and their ready-to-eat [RTE] salads carry a risk of foodborne pathogens? Anaerobe. 2011;17(6):286–7.
Mritunjay SK, Kumar V. A study on prevalence of microbial contamination on the surface of raw salad vegetables. Biotech. 2017;7(1):13.
Kuan CH, Rukayadi Y, Ahmad SH, Che WJWMR, Thung TY, Chang WS, Loo YY, Tan CW, Ramzi OB. Comparison of the microbiological quality and safety between conventional and organic vegetables sold in Malaysia. Front Microbiol. 2017;8:1433.
Tango CN, Wei S, Khan I, Hussain MS, Kounkeu PN, Park JH, Kim SH, Oh DH. Microbiological quality and safety of fresh fruits and vegetables at retail levels in Korea. J Food Sci. 2018;83(1):386–92.
Ajayeoba TA, Atanda OO, Obadina AO, Bankole MO, Adelowo OO. The incidence and distribution ofListeria monocytogenesin ready-to-eat vegetables in South-Western Nigeria. Food Science & Nutrition. 2016;4(1):59–66.
ZHANG X: Development status and countermeasures of vegetable and processed export industry in China. China Vegetables 2018.Article in Chinese.
Zheng L: Quantitative risk assessment and Management of Listeria monocytogenes in ready-to-eat salads. University of Shanghai for science and technology 2012.Article in Chinese.
Jonquières R, Bierne H, Mengaud J, Cossart P. The inlA gene of Listeria monocytogenes LO28 harbors a nonsense mutation resulting in release of Internalin. Infection & Immunity. 1998;66(7):3420–2.
Felício MTS, Hogg T, Gibbs P, Teixeira P, Wiedmann M. Recurrent and sporadic Listeria monocytogenesContamination in Alheiras represents considerable diversity, including virulence-attenuated isolates. Appl Environ Microbiol. 2007;73(12):3887–95.
Wang H, Luo L, Zhang Z, Deng J, Wang Y, Miao Y, Zhang L, Chen X, Liu X, Sun S. Prevalence and molecular characteristics of Listeria monocytogenes in cooked products and its comparison with isolates from listeriosis cases. Frontiers of medicine. 2018;12(1):104–12.
Doumith M, Buchrieser C, Glaser P, Jacquet C, Martin P. Differentiation of the major Listeria monocytogenes serovars by multiplex PCR. J Clin Microbiol. 2004;42(8):3819–22.
Orsi RH, den Bakker HC, Wiedmann M. Listeria monocytogenes lineages: genomics, evolution, ecology, and phenotypic characteristics. Int J Med Microbiol. 2011;301(2):79–96.
Maury MM, Tsai YH, Charlier C, Touchon M, Chenalfrancisque V, Leclercq A, Criscuolo A, Gaultier C, Roussel S, Brisabois A. Uncovering Listeria monocytogenes hypervirulence by harnessing its biodiversity. Nat Genet. 2016;48(3):308–13.
Chau ML, Aung KT, Hapuarachchi HC, Lee PS, Lim PY, Kang JS, Ng Y, Yap HM, Yuk HG, Gutiérrez RA, Ng LC. Microbial survey of ready-to-eat salad ingredients sold at retail reveals the occurrence and the persistence of Listeria monocytogenes sequence types 2 and 87 in pre-packed smoked salmon. BMC Microbiol. 2017;17(1):46.
Cotter PD, Draper LA, Lawton EM, Daly KM, Groeger DS, Casey PG, Ross RP, Hill C. Listeriolysin S, a novel peptide Haemolysin associated with a subset of lineage I Listeria monocytogenes. PLoS Pathog. 2008;4(9):e1000144.
Quereda JJ, Nahori MA, Meza-Torres J, Sachse M, Titos-JimeÂnez P, Gomez-Laguna J, Dussurget O, Cossart P, Pizarro-Cerdá J. Listeriolysin S is a Streptolysin S-like virulence factor that targets exclusively prokaryotic cells in vivo. MBio. 2017;8(2). pii: e00259-17.
Wang Y, Jiao Y, Lan R, Xu X, Liu G, Wang X, Zhang L, Pang H, Jin D, Dai H. Characterization of Listeria monocytogenes isolated from human Listeriosis cases in China. Emerging Microbes & Infections. 2015;4(8):e50.
Pagliano P, Arslan F, Ascione T. Epidemiology and treatment of the commonest form of listeriosis: meningitis and bacteraemia. Infez Med. 2017;25(3):210–6.
Escolar C, Gómez D, Rota García MD, Conchello P, Herrera A. Antimicrobial resistance profiles of Listeria monocytogenes and Listeria innocua isolated from ready-to-eat products of animal origin in Spain. Foodborne Pathogens & Disease. 2017;14(6):357–63.
Moreno LZ, Paixão R, Gobbi DD, Raimundo DC, Ferreira TP, Moreno AM, Hofer E, Reis CM, Matté GR, Matté MH. Characterization of antibiotic resistance in Listeria spp. isolated from slaughterhouse environments, pork and human infections. Journal of Infection in Developing Countries. 2014;8(4):416–23.
Gombas DE, Chen Y, Clavero RS, Scott VN. Survey of Listeria monocytogenes in ready-to-eat foods. J Food Prot. 2003;66(4):559–69.
Chen M: Study on the genetic diversity and quorum sensing of foodborne Listeria monocytogenessouth China University of Technology; 2014.Article in Chinese.
Ragon M, Wirth T, Hollandt F, Lavenir R, Lecuit M, Le MA, Brisse S. A New Perspective on Listeria monocytogenesEvolution. PLoS Pathog. 2008;4(9):e1000146.
Cheng J, Chen M, Chen Y, Zhang J, Wu Q. Exploring potential mechanism of quinolone resistance of foodborne Listeria monocytogenes isolates. J Food Sci Technol. 2018;36(04):32–40 Article in Chinese.
Nightingale KK, Ivy RA, Ho AJ, Fortes ED, Njaa BL, Peters RM, Wiedmann M. inlA premature stop codons are common among Listeria monocytogenes isolates from foods and yield virulence-attenuated strains that confer protection against fully virulent strains. Applied & Environmental Microbiology. 2008;74(21):6570–83.
Roldgaard BB, Andersen JB, Hansen TB, Christensen BB, Licht TR. Comparison of three Listeria monocytogenes strains in a Guinea-pig model simulating food-borne exposure. FEMS Microbiol Lett. 2010;291(1):88–94.
Wu S, Wu Q, Zhang J, Chen M, Guo W. Analysis of multilocus sequence typing and virulence characterization of Listeria monocytogenes isolates from Chinese retail ready-to-eat food. Front Microbiol. 2016;7:168.
Gelbíčová T, Koláčková I, Pantůček R, Karpíšková R. A novel mutation leading to a premature stop codon in inlA of Listeria monocytogenes isolated from neonatal listeriosis. New Microbiol. 2015;38(2):293–6.
Clinical and Laboratory Standard Institute: Performance Standards for Antimicrobial Susceptibility Testing: 27th Informational Supplement (M100-27th ed.). Wayne, PA: Clinical and Laboratory Standards Institute. January, 2017.
Clinical and Laboratory Standard Institute: Methods for antimicrobial dilution and disk susceptibility testing for infrequently isolated or fastidious bacteria; approved guideline-second edition. M45-A2, vol. 30, no. 18. Wayne, PA: Clinical and Laboratory Standard Institute. 2014.
Magiorakos AP, Srinivasan A, Carey RB, Carmeli Y, Falagas ME, Giske CG, Harbarth S, Hindler JF, Kahlmeter G, Olssonliljequist B. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–81.
Notermans SH, Dufrenne J, Leimeisterwächter M, Domann E, Chakraborty T. Phosphatidylinositol-specific phospholipase C activity as a marker to distinguish between pathogenic and nonpathogenic Listeria species. Applied & Environmental Microbiology. 1991;57(9):2666–70.
Xu X, Wu Q, Zhang J, Deng M, Zhou Y. Studies on specific detection of Listeria monocytogenes in foods by duplex PCR. Chinese Journal of Health Laboratory Technology. 2009;19(6):1199–201 Article in Chinese.
Chen M, Wu Q, Zhang J, Guo W, Wu S, Yang X. Prevalence and contamination patterns of Listeria monocytogenes in Flammulina velutipes plants. Foodborne Pathogens & Disease. 2014;11(8):620–7.
Hudson JA, Lake RJ, Savill MG, Scholes P, Mccormick RE. Rapid detection of Listeria monocytogenes in ham samples using immunomagnetic separation followed by polymerase chain reaction. J Appl Microbiol. 2001;90(4):614–21.
Furrer B, Candrian U, Hoefelein C, Luethy J. Detection and identification of Listeria monocytogenes in cooked sausage products and in milk by in vitro amplification of haemolysin gene fragments. J Appl Microbiol. 1991;70(5):372–9.
Suárez M, Gonzálezzorn B, Vega Y, Chicocalero I, Vázquezboland JA. A role for ActA in epithelial cell invasion by Listeria monocytogenes. Cell Microbiol. 2001;3(12):853–64.
Clayton EM, Hill C, Cotter PD, Ross RP. Real-time PCR assay to differentiate Listeriolysin S-positive and -negative strains of Listeria monocytogenes. Appl Environ Microbiol. 2011;77(1):163–71.
Acknowledgements
We like to thank the team of curators of the Institute Pasteur MLST databases for curating the data and making them publicly available at http://bigsdb.pasteur.fr/.
Funding
We would like to acknowledge the financial support from the National Natural Science Foundation of China (31701718, 31501580), Natural Science Foundation of Guangdong Province, China (2017A030313173), Pearl River S&T Nova Program of Guangzhou (201710010018), GDAS’ Special Project of Science and Technology Development (2017GDASCX-0201), and the Science and Technology Planning Project of Guangdong Province (2016A0101–05012). The funding bodies had no role in the design of the study, in the collection, analysis, interpretation of data, or in the writing of the manuscript.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its supplementary information files.
Author information
Authors and Affiliations
Contributions
QW, YC, and MC conceived and designed the experiments. YC, JC, LF, MC and TL performed the experiments. YZ, HZ, RP, XW and QY conducted bioinformatics analyses. YC, MC, QW, and JZ drafted the manuscript. QW, YD, JW, JB and JZ reviewed the final manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional file
Additional file 1:
Table S1. The detail detection results of Listeria monocytogenes in 419 vegetable samples. Table S2. Listeria monocytogenes strains isolated from fresh vegetables Figure S1. Serogroup analysis of Listeria monocytogenes strains isolated from fresh vegetable samples by multiplex PCR. The strain no. 1–30 correspond to Table S2. Figure S2. The presence of virulence-related genes in Listeria monocytogenes isolated from fresh vegetable samples. A, prfA; B, mpl; C, plcA; D, inlB; E, plcA; F, hly; G, iap; H, actA; I, llsX; J, ptsA.*:The strain no. 1–30 correspond to Additional file 1: Table S2. (ZIP 8979 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Chen, M., Chen, Y., Wu, Q. et al. Genetic characteristics and virulence of Listeria monocytogenes isolated from fresh vegetables in China. BMC Microbiol 19, 119 (2019). https://doi.org/10.1186/s12866-019-1488-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12866-019-1488-5