
Abstract
Background
The incidence of dementia and cognitive decline is increasing with no therapy or cure. One of the reasons treatment remains elusive is because there are various pathologies that contribute to age-related cognitive decline. Specifically, with Alzheimer’s disease, targeting to reduce amyloid beta plaques and phosphorylated tau aggregates in clinical trials has not yielded results to slow symptomology, suggesting a new approach is needed. Interestingly, exercise has been proposed as a potential therapeutic intervention to improve brain health and reduce the risk for dementia, however the benefits throughout aging are not well understood.
Results
To better understand the effects of exercise, we preformed transcriptional profiling on young (1–2 months) and midlife (12 months) C57BL/6 J (B6) male mice after 12 weeks of voluntary running. Data was compared to age-matched sedentary controls. Interestingly, the midlife running group naturally broke into two cohorts based on distance ran - either running a lot and more intensely (high runners) or running less and less intensely (low runners). Midlife high runners had lower LDL cholesterol as well as lower adiposity (%fat) compared to sedentary, than midlife low runners compared to sedentary suggesting more intense running lowered systemic markers of risk for age-related diseases including dementias. Differential gene analysis of transcriptional profiles generated from the cortex and hippocampus showed thousands of differentially expressed (DE) genes when comparing young runners to sedentary controls. However, only a few hundred genes were DE comparing either midlife high runners or midlife low runners to midlife sedentary controls. This indicates that, in our study, the effects of running are reduced through aging. Gene set enrichment analyses identified enrichment of genes involved in extracellular matrix (ECM), vascular remodeling and angiogenesis in young runners but not midlife runners. These genes are known to be expressed in multiple vascular-related cell types including astrocytes, endothelial cells, pericytes and smooth muscle cells.
Conclusions
Taken together these results suggest running may best serve as a preventative measure to reduce risk for cerebrovascular decline. Ultimately, this work shows that exercise may be more effective to prevent dementia if introduced at younger ages.
Similar content being viewed by others


Background
With an aging population, the impact of age-related cognitive impairment is increasing [1,2,3,4]. Cognitive impairment can involve many symptoms, including memory loss, newfound difficulties with language, and inability to make every-day decisions [5]. Further, the incidence of many dementias for which aging is the major risk factor is increasing. For example, Alzheimer’s Disease (AD), with approximately 5.4 million people affected in the 2016, is expected to increase to over 13 million in the next few years [6]. The majority of clinical trials for AD therapeutics have focused on reducing hallmark pathologies such as amyloid beta accumulation and tau tangles. However, these pathologies do not always correlate to cognitive function, and therefore these therapeutic targets may not rescue the symptoms that ultimately burden the patient [7, 8]. Currently, despite numerous preclinical studies and clinical trials, there are no therapies for cognitive decline.
Non-pharmacological interventions have been proposed as alternatives to pharmacological treatments to prolong brain health – reducing risk for age-related cognitive decline and dementias. Obesity and physical inactivity increase risk for cognitive decline and dementia, suggesting that interventions such as diet and exercise can mitigate risk [9]. Exercise has positive effects, not just improving systemic health, but also cerebral health through increases in cerebral plasticity, neurogenesis, as well as hippocampal and cortical volume [10, 11]. Exercise earlier in life correlated with reduced cognitive impairment with age [12]. The cerebral benefits of exercise may arise through the induction of brain derived neurotropic factor (BDNF), which triggers neuronal proliferation in the dentate gyrus [13]. A recent study has also explored the use of running, as well as an AAV-based gene therapy to increase neuronal proliferation and survival, in a mouse model of AD [14]. Viral-induced neurogenesis alone did not benefit cognition as well as running did, suggesting that running also promotes non-neuronal changes that improve cognition. Therefore, the full spectrum of processes by which running promotes brain health and reduces risk for dementias remains unclear. Furthermore, given that many such studies are performed in young mice, it remains unknown whether the beneficial effects of running persist through multiple life stages.
Here, we chose an unbiased transcriptional profiling approach to better understand the effects of running on overall brain health. To date, an extensive evaluation of the transcriptome of the brain in response to running across ages has not been assessed. RNA sequencing was performed on the cortex and hippocampus from young and middle aged (midlife) C57BL/6 J (B6) male mice that were provided running wheels for 12 weeks. Transcriptional profiles were compared to aged-matched sedentary controls. Within the midlife running cohort, half the mice ran markedly faster and farther (high runners) than the other half (low runners). This provided natural variation in our midlife running dataset and allowed us to also interrogate how the intensity of running impacted systemic health as well as transcriptional profiles in the brain. The young cohort did not exhibit such variation in their voluntary exercise. Transcriptomes of young running mice showed considerably more differentially expressed (DE) genes compared to either low-running or high-running midlife cohorts. Gene set enrichment analyses revealed enrichment of genes in pathways implicated in vascular remodeling in young, but not midlife mice.
Results
Voluntary running distances at midlife showed a bimodal distribution
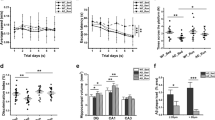
To understand the molecular changes in the brain in response to voluntary running, running wheels were provided to young (1–2 month old, mo) and midlife (12 mo) C57BL/6 J (B6) male mice for 12 weeks. Age-matched sedentary controls had no access to a running wheel (Fig. 1a). To quantify running, wheel rotations per minute were assessed overnight, when mice are most active, for at least five nights during the last week of the experiment. These data were used as an estimate for the overall running performed during the 12 weeks. Although there was some variation between average wheel rotations per night in the young cohort, five of six mice averaged over 10,000 rotations per night. Of the 12 midlife running mice however, half averaged fewer than 10,000 rotations per night deemed ‘Low Runners’ (gray), and half averaged greater than 10,000 rotations per night, deemed ‘High Runners’ (black) (Fig. 1b). Of the minutes active, the high running cohort ran faster (average rotations per minute of active minutes) than the low running cohort (Fig. 1c). Direct comparison of speed and distance between midlife low and high runners revealed significant differences (p < 0.001) (Fig. 1d, Additional file 1: Figure S1A). This was not due to dominance in group housing (Additional file 9: Table S1). High runners ran a similar distance (on average, greater than 10,000 rotations per night) to young runners. Additionally, midlife high runners spent more time running compared to midlife low runners (Additional file 1: Figure S1B). High runners also were quantified to have run at 100 rotations per minute, while the low runners did not show this ability (Additional file 1: Figure S1C). Young sedentary mice gained more weight over the course of the experiment than young running mice (Fig. 1e, f). There was no significant difference in weight from the start of the experiment to the end between midlife running and midlife sedentary mice (Fig. 1f, Additional file 1: Figure S1D). To better understand body composition in the midlife cohort, mice were subjected to NMR a week before harvest to assess fat and lean mass percentages. Midlife high runners showed a significant decrease in fat mass (g) reflected by a corresponding decrease in adiposity (%fat) (Additional file 1: Figure S2A, S2B, S2D). Lean muscle mass was not different among the groups but the midlife high running cohort had a higher percent lean muscle mass compared to low running and sedentary cohorts, presumably due to the lower body weight of the high runners (Additional file 1: Figure S2C, S2E). This indicates that the weight composition was more favorable (high lean muscle percentage, low fat percentage) in the high runners compared to the other midlife mice.

Voluntary Running at young and midlife reveals natural variation in running intensity. a Experimental strategy of running mice. Young mice were given access to voluntary running wheels for 12 weeks starting at 1–2 months of age and ending at 4–5 months of age. Midlife mice were given access to wheels for 12 weeks starting at 12 months of age and ending at 15 months of age. b Average wheel rotations per night showed dichotomous response of midlife running. Midlife mice that ran below 10,000 rotations per night (red line), were deemed ‘Low’ (grey). Midlife mice that ran above 10,000 rotations per night, deemed ‘High’ (black). c Graph of distance vs speed showed differences between low and high running groups; all midlife high runners trended faster and farther than midlife low runners. d Midlife running speed between low and high runner groups showed a significant difference (p < 0.001). e Weight over the course of running experiment. f Change in weight of each cohort from start to end of the experiment (p < 0.001)
To assess whether running during midlife altered metabolic indicators of health in the blood, we measured cholesterol composition, non-fasted glucose, triglycerides, and non-esterified fatty acids (NEFA) levels. Total cholesterol was significantly reduced in young runners compared to age-matched sedentary controls (Fig. 2a). Midlife high runners also showed a significantly lower total cholesterol profile compared to midlife sedentary controls, which could be attributed to the decrease in LDL cholesterol (Fig. 2a-c). HDL cholesterol remained unchanged with running (Fig. 2c). Non-fasted glucose levels were not significantly different, although there was trend towards lower blood glucose in running compared to sedentary mice at both ages (Fig. 2d). Triglycerides and NEFA were reduced by running at a young age, but this effect was not seen in the midlife cohort (Fig. 2e, f). Taken together, there was a significant shift towards a healthier body composition and blood profile in midlife high runners compared to low runners. Further, systemic health benefits such as levels of total cholesterol, including LDL, can be altered at midlife due to more intense running.

Lipid profiling of the blood showed ability to alter cholesterol composition in midlife. a Significant difference in total cholesterol plasma concentration at harvest in young runners and between midlife high runners to midlife sedentary mice (young *p = 0.0124, midlife *p = 0.0282). b No significant difference in High Density Lipoprotein (HDL) plasma concentration between runners and sedentary mice. c Significant difference in Low Density Lipoprotein (LDL) plasma concentration between midlife high runners and midlife sedentary mice (*p = 0.0489). d No change in non-fasted glucose plasma concentration across all cohorts. e Significant decrease in in plasma triglyceride between young run and young sedentary concentration (*p = 0.0151). f Significant reduction in Non-Esterified Fatty Acid (NEFA) plasma concentrations between young run and young sedentary cohorts (*p = 0.0374)
Young running affects transcriptional signatures more robustly than midlife running
To assess transcriptional changes in the brain after running at two different ages, RNA-seq was performed on hippocampus and cortex – vulnerable regions in age-related cognitive decline and dementia (Additional file 1: Figures S3 and S4). Differentially expressed (DE) genes (FDR < 0.05) were identified by comparing (i) young running to young sedentary, (ii) midlife high runners to midlife sedentary, and (iii) midlife low runners compared to midlife sedentary (Additional file 2: Table S2, Additional file 3: Table S3, Additional file 4: Table S4, Additional file 5: Table S5, Additional file 6: Table S6, Additional file 7: Table S7 and Additional file 8: Table S8, Additional file 1: Figure S5, see Methods). In the cortex, there were 1252 DE genes (742 upregulated, 510 downregulated) when comparing young runners to young sedentary mice (Fig. 3a, b). However, there were only 214 DE genes (70 upregulated, 144 downregulated) in the midlife high runners compared to midlife sedentary, and 67 DE genes (20 upregulated and 47 downregulated) comparing midlife low runners to midlife sedentary (Fig. 3b). Hippocampal analysis revealed similar results to the cortex. In young mice there were 2026 DE genes (1498 upregulated, 528 downregulated) in the young runners compared to young sedentary (Fig. 3f, g). In midlife mice, there were 271 DE genes (87 upregulated, 184 downregulated) comparing midlife high runners to midlife sedentary controls, and 172 DE genes (80 upregulated, 92 downregulated) comparing midlife low runners to midlife sedentary controls (Fig. 3g). These results show that transcriptionally, running at a young age has a far greater effect on the number of DE genes than at midlife.

RNA-seq analysis identified ECM-related enrichment in both the cortex and hippocampus of young runners. a Cortical region (purple) used for RNA-seq. b Number of DE genes (FDR < 0.05) found in the cortex of the young runners compared to young sedentary (‘Young Run’), midlife low runners compared to midlife sedentary (‘Low Run’), and midlife high runners compared to midlife sedentary (‘High Run’) (FDR < 0.05). c IPA canonical pathway analysis showed enriched ‘Hepatic Fibrosis’ and ‘GP6 Signaling Pathway’ in the cortex of young run compared to young sedentary, however not significant in midlife comparisons. d KEGG pathway enrichment analysis in the cortex for young run compared to young sedentary (FDR < 0.05). e GO term enrichment analysis in the cortex for young run compared to young sedentary (FDR < 0.05). f Hippocampal region (green) used for RNA-seq tissue submission. g Number of DE genes (FDR < 0.05) found in the hippocampus of the young runners compared to young sedentary (‘Young Run’), midlife low runners compared to midlife sedentary (‘Low Run’), and midlife high runners compared to midlife sedentary (‘High Run’) (FDR < 0.05). h IPA canonical pathway analysis showed enriched ‘Hepatic Fibrosis’ and ‘GP6 Signaling Pathway’ in the hippocampus of young run compared to young sedentary, however not significant in midlife comparisons. i KEGG pathway enrichment analysis in the hippocampus for young run compared to young sedentary (FDR < 0.05). j GO term enrichment analysis in the hippocampus for young run compared to young sedentary (FDR < 0.05)
Extracellular matrix-related genes are altered in young but not midlife runners
Next, we sought to identify which pathways and genes were significantly altered by running at a young age, but showed no difference at midlife. We first analyzed transcriptional data through Ingenuity Pathway Analysis (IPA) canonical pathway analysis. DE genes comparing young runners to young sedentary controls were enriched for canonical pathway terms ‘Hepatic Fibrosis’, ‘GP6 Signaling’, and ‘Circadian Rhythm Signaling’ in the cortex and hippocampus. (Fig. 3c, h). These pathways were not enriched in the midlife data. ‘Hepatic Fibrosis’ and ‘GP6 Signaling Pathway’ terms contain extracellular matrix (ECM)-related genes, such as collagens and laminins. KEGG pathway analysis further identified ‘ECM-receptor Interaction’ and ‘Focal Adhesion’ in the cortex and hippocampus, which substantiated the significant changes to the ECM (Fig. 3d, i). There were no KEGG terms enriched in either the cortex or hippocampus for the midlife low runners. Only the KEGG pathway ‘Malaria’ was enriched in the cortex of midlife high runners. Genes DE in the ‘Malaria’ pathway are primarily hemoglobin-related genes suggesting a potential change in oxygenation in high runners. Only the KEGG pathway ‘ECM-receptor Interaction’ was enriched in the hippocampus of midlife high runners. Of the ten DE genes in ‘ECM-receptor Interaction’ were seven collagen or laminin genes (Col24a1 (FDR = 2.39e-6, FC = − 2.17), Col6a3 (FDR = 0.026,FC = − 1.38), Col6a2 (FDR = 0.0032,FC = -1.45), Col6a5 (FDR = 0.00029,FC = − 1.72), Col6a1 (FDR = 0.00025,FC = − 1.52), Col5a1 (FDR = 9.03e-8,FC = − 1.75), and Lamc2 (FDR = 6.85e-8,FC = − 2.07)) which were all downregulated. This suggests that more intense running at midlife can influence ECM-related genes, but potentially not in a positive way.
GO terms are a way of categorizing DE genes into functional biological groups. GO term analysis of the DE genes from the cortex and hippocampus data from young runners showed enrichment for ‘ECM Organization’ and ‘Cell Adhesion’ (Fig. 3e, j). Interestingly, vascular remodeling-related terms were significantly enriched in the cortex of young runners, including terms such as ‘blood vessel remodeling’, ‘ECM Organization’, ‘Angiogenesis’, and ‘Cell Adhesion’ (Fig. 3e). None of these terms were enriched in the midlife data, irrespective of distance ran. However, one GO term, ‘Cellular Oxidant Detoxification’, which comprised mainly hemoglobin component genes, was enriched in both low and high runners in the hippocampus but only in the high runners of the cortex at midlife. These data suggest there is an overall change in ECM composition or organization due to running at a young age that is not reflected to the same degree at midlife. (Fig. 3).
Running upregulates genes related to vascular remodeling in young but not midlife mice
IPA revealed ‘Hepatic Fibrosis’ and ‘GP6 Signaling’ as significantly enriched canonical pathways in both the cortex and the hippocampus (Fig. 3). These pathways contain many collagens and laminins that comprise the basement membrane component of the blood brain barrier that is important for maintaining brain health (Fig. 4a). The majority of genes in ‘GP6 Signaling’ pathway were upregulated in the young runners in both the cortex and hippocampus but were not DE in either midlife high or low runners (Fig. 4b-f).

General upregulation of Collagens and Laminins due to young running with an attenuated response at midlife. a Number of significant collagen, laminin or other genes in young ‘Hepatic Fibrosis’ and ‘GP6 Signaling’ pathways in the cortex and hippocampus. b Fold changes of genes enriching for ‘GP6 Signaling’ in the cortex. c Fold changes of genes enriching for ‘GP6 Signaling’ in the hippocampus. d Heatmap of Col4s and laminins in the cortex and hippocampus that are significant in young run compared to young sedentary, while not significant in midlife comparisons. e Examples of significant cortical genes found in (d) showing attenuated responses in midlife cohort contrasts compared to young. f Examples of significant hippocampal genes found in (d) showing various responses in midlife cohort contrasts compared to young
GO term analysis identified enrichment of ‘ECM organization’, ‘Blood Vessel Remodeling’ and ‘Angiogenesis’ in the cortex (Fig. 5a). These terms were significantly enriched in young mice but not in midlife mice. Of these vascular terms, ‘Angiogenesis’ contained 32 genes (26 upregulated, 6 downregulated) (Fig. 5a, b). At least six of the upregulated DE genes are implicated in Vegf-induced angiogenesis (Mmp14 (FDR = 1.23e-8, FC = 1.75), Vegfa (FDR = 0.038, FC = 1.20), Kdr (FDR = .0011, FC = 1.41), Flt1 (FDR = .031, FC = 1.21), Dll4 (FDR = 0.037, FC = 1.59), Notch1 (FDR = 0.002, FC = 1.29)) (Fig. 5c, d) [15]. However, of the 26 DE genes that were significantly upregulated in young runners compared to young sedentary, only two genes (Col8a1, Hif3a) were significantly DE in midlife high runners when compared to midlife sedentary controls (Fig. 5c-d).

Angiogenesis genes are significantly enriched in young runners but not midlife runners. a GO terms enriched for vascular remodeling in the cortex show general upregulation. b Fold changes for DE genes enriched in the Angiogenesis term in the cortex. c Heatmap comparison between angiogenesis gene fold changes in the cortex in the young contrast (all significant) and midlife contrasts. d Examples of significant cortical angiogenesis genes found in (c) showing various responses in midlife cohort contrasts compared to young
Running activates genes in multiple cerebrovascular-related cell types
Finally, we determined whether specific cell types were more dramatically affected by young but not midlife running. This might provide insight into which cell type(s) are no longer responding to running in midlife mice. A sample of basement membrane and angiogenesis genes DE in young, but not midlife, running datasets were cross referenced to two cell-type specific datasets – the Brain RNA seq and a single cell RNA-seq dataset focused on cerebrovascular associated cells [16, 17]. First, we evaluated Col4 and laminin genes that were DE in young runners. As expected, the majority of the genes were expressed by cerebrovascular-related cells such as astrocytes and endothelial cells and not other cells in brain such as neurons, oligodendrocytes and microglia (Additional file 10: Table S9). However, many genes were expressed by different subsets of vascular-related cells such as Col4a1 (endothelial cells, pericytes, vascular smooth muscle cells and fibroblasts) Lamb2 (astrocytes, endothelial cells, pericytes, vascular smooth muscle cells and fibroblasts) and Lama5 (astrocytes, endothelial cells and vascular smooth muscle cells) (Additional file 1: Figure S6). Second, we assessed cell type specific expression of angiogenesis genes. Upstream components of the angiogenesis pathway were mainly expressed by astrocytes, including Mmp14 and Vegfa (Fig. 6a, b). Downstream components of the angiogenesis pathway were primarily expressed by endothelial cells (Kdr, Flt1, Dll4, Notch1). Therefore, ECM- and angiogenesis-related genes affected by running are expressed in multiple cell types relevant to the cerebrovasculature. The results are summarized in Fig. 7.

Angiogenesis genes are primarily expressed in astrocytes and endothelial cells. a. Expression of Mmp14 is primarily produced in pericytes and astrocytes. b Expression of Vegfa is primarily produced by astrocytes. c Expression of Flt1 is primarily produced in endothelial cells. d Expression of Dll4 is primarily produced in endothelial cells. Cell type specific expression reproduced from Zheng et al. (left), and Vanlandewijck and He et al. (right)

Running young is more effective than running at midlife. a Depiction of transcriptional changes occurring in a young mouse due to running, including upregulation of collagens and laminins from astrocytes, pericytes and endothelial cells, as well as induction of the VEGFA-KDR-DLL4 angiogenesis pathway. b Depiction of molecular changes occurring in midlife mice due to running, including downregulation of collagens and laminins, and a trend for induction of early angiogenesis genes from astrocytes (MMP14, VEGFA) but an attenuated downstream angiogenesis response (FLT-1, DLL4, NOTCH1) in endothelial cells
Discussion
Here, we assessed B6 male mice at two ages, young and midlife, to better understand the effects of running on the aging brain. Our data show early indication that intervention of running may not be as beneficial to the cerebrovasculature in midlife as it is at a young age. While systemic health (lipid profile, body composition) is improved by running at midlife, transcriptional profiling of the hippocampus and cortex showed fewer genes impacted during midlife, specifically in the cerebrovasculature. This suggests there may be a reduced responsiveness to running by specific cerebrovascular cell types in these brain regions with age. Therapies or interventions during midlife may require different approaches than preventative measures in young, more responsive individuals. In this study, half the midlife cohort (high runners) ran as much as the young cohort, whereas the other half ran far less (low runners, Fig. 1). Interestingly, this did not drastically affect the transcriptional profiles (Fig. 3). We would have anticipated that running further and faster would have been more beneficial at midlife, however, our results show a mitigated transcriptional response in all midlife runners compared to young runners.
Previous exercise studies in mouse models have primarily focused on the benefits of exercise in young rodents. Findings from these experiments are promising, showing that running can reduce amyloid plaque development, induce neurogenesis, reduce infarct volume, and improve cognition [14, 18,19,20]. However, the brain during development and early adult life is considered more plastic than in later life, which indicates that therapies found to be beneficial in young, may not properly translate to aged trials [2, 21].
We took an unbiased approach to identify transcriptional changes in two vulnerable brain regions (cortex and hippocampus) as a result of running. We show that many of the transcriptional changes as a result of running were to genes relevant to the cerebrovasculature. Particularly, we highlight genes involved in basement membrane composition (particularly collagens and laminins) that are upregulated in young mice due to running that were not affected by running at midlife. Similarly, genes in the angiogenesis pathway (e.g. Mmp14, Vegfa, Kdr, Flt1, Dll4, and Notch1) showed induction by running in young but not midlife mice [15]. These transcriptional data suggest that the benefits of exercise to the cerebrovasculature declines with age, increasing the need for a greater understanding of midlife exercise as an intervention to improve cerebrovascular health [22].
Previous studies have focused on the neuroprotective benefits of exercise. In Choi et al., of the 5XFAD mice (a model relevant to AD) that were exercised, half were categorized as showing increased neurogenesis while the other half did not show this effect, potentially due to the amount run [14]. Additionally, it was shown that viral-induced neurogenesis was not enough to rescue cognitive decline, while exercise-induced neurogenesis was able to sufficiently improve cognitive results [14]. This finding indicates that running is providing additional benefits to substantiate neurogenesis mediated improvements in cognition. Based on the results of our study, we predict that in Choi et al. there were running-induced improvements to the cerebrovasculature which would provide better clearance of amyloid and better maintenance of neuronal health resulting in improved cognition [14]. It is known that the development of the neuronal and vasculature systems is strongly linked, with growth occurring simultaneously because neurons require vascular support for oxygen and nutrients. This further supports the need for cerebrovascular improvement to accompany adult therapy-induced neurogenesis [23]. If neurogenesis and angiogenesis become uncoupled, this may cause stress to neurons, reduced cerebral blood flow, and a disruption to neurovascular coupling. Therefore, when considering midlife interventions to cognitive decline and dementia it is important to consider both neuronal and cerebrovascular health.
Our study assessed both distance and intensity of running. Although most previous studies tend to only report time allotted for running, a few studies included more detailed breakdown of running data. For instance, in one study, young (3mo) and aged (19mo) male mice were analyzed for total distance over a day and had no significant difference with 3.0 ± 0.12 miles/day for young, and 2.4 ± 0.06 miles/day for old [11]. The mice used in this study had an average of 1.32 ± 0.69 miles/12 h for young mice, 0.28 ± 0.13 miles/12 h for midlife low runners, and 2.00 ± 0.50 miles/12 h for midlife high runners. Results from this study found that there was a distinct loss of vascular plasticity in older mice, substantiating the findings from our study [11]. Another study in young C57BL/6 J female mice explored differential gene expression in the striatum after 16 days of running in conjunction with voluntary ethanol consumption. The female running mice revealed 1305 DE genes due to wheel running, compared to our 1252 in DE genes in the young running cortex, and 2026 DE genes in the young running hippocampus [24]. Running distances cannot be compared as Darlington et al. used a vertical, external to the cage, larger diameter, running wheel design, dissimilar to the one used in our study [24].
One limitation of our study is that we used healthy wild-type male mice to assess the effects of exercise at two different ages. In our study, the effects of exercise were greater in young compared to midlife male mice. Future studies are necessary to understand the sex-specific effects of running, and how these differences might play a role in exercise-induced transcriptional differences. Additionally, this study did not extend past transcriptional profiling. Further assessment of vascular density and functional testing of responsiveness through cerebral blood flow (CBF) measurements would aid in our understanding of how running improves cerebrovascular health through aging. Moreover, C57BL/6 J do not show cognitive decline at these early and midlife ages, so cognition was not assessed. Determining the relationship between cerebrovascular health and cognition are still required. It is also not clear how running impacts cerebrovascular health in humans and animal models predisposed to vascular damage and dementia through genetic or environmental risk factors.
Due to the increasing recognition of a vascular contribution to dementia symptomology, it would be pertinent to know if running alleviates these effects of vascular risk in genetically predisposed mice at different ages. For example, the ε4 allele of apolipoprotein E (APOEε4), the greatest risk factor for late-onset AD and Vascular Dementia, is associated with early cerebrovascular decline in both mice and humans [25, 26]. This is proposed to be due to a lack of binding of APOEε4 to the low-density lipoprotein receptor related protein 1 (LRP1) leading to an increase in MMP9 and a breakdown of basement membrane proteins and endothelial cell tight junctions [25]. Therefore, when evaluating running as a potential intervention, it is imperative to understand whether benefits will be seen across multiple dementia risk genotypes. Although these experiments are challenging to perform in human populations, they can be readily performed in mouse models.
Due to the heterogeneity of dementia pathology, it is possible that the typical approach to treating dementias, is too narrow in scope, only able to alleviate the burden of a small subset of dementia cases. Only recently has the American Heart Association acknowledged the prevalence of Vascular Contributions to Cognitive Impairment and Dementia (VCID), explaining that cerebral infarcts are frequent and common with age, including in patients with diagnosed with AD [27]. We hypothesize that exercise is key in identifying new pathways, specifically those related to cerebrovascular health, that may help a broader population of dementia patients. In summary, our research and others show that exercise benefits cerebral health by improving multiple systems including the cerebrovasculature. However, the benefits of exercise appear to decline with age, further supporting that combinatorial approaches are required to prevent cognitive decline and dementias.
Conclusions
This study identifies genes and pathways induced by voluntary running in young mice. These genes include those involved in cerebrovascular remodeling such as basement membrane production and angiogenesis. In addition, these data propose a mechanism that running at middle life, independent of running intensity, is less effective at activating genes relevant to cerebrovascular remodeling. These data suggest renewed efforts are needed to identify those at risk of cerebrovascular decline as early as possible, and that preventative strategies that involve exercise to enhance cerebrovascular health maybe more effective in younger compared to older individuals.
Methods
Mouse strains
All experiments involving mice were conducted with approval and accordance described in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. All experiments were approved by the Animal Care and Use Committee at The Jackson Laboratory. All mice used in this study were male C57BL/6 J (B6, stock number JR00664, The Jackson Laboratory). Mice were kept in a 12/12-h light/dark cycle and fed ad libitum 6% kcal fat standard mouse chow. A total of 48 mice were used in this study.
Exercise by voluntary running
Group housed mice (two-three per pen) were provided access to low profile saucer wheels (Innovive Inc) 24 h a day for 12 weeks. Sedentary mice did not have access to running wheels. In the first experiment, young mice were housed from wean (1-2mo) until the end of the experiment (4-5mo). Two cohorts of mice were used to evaluate running at the young timepoint; the first group (n = 6 per activity) had weights, running tracked, and blood lipid profiling, while the second group (n = 5 per activity) had RNA-seq run on the cortex and hippocampus. In the second experiment (n = 12 for running and sedentary), mice were aged without wheel access, and at 12mo were provided running wheels. For both experiments, in the final week, mice were individually housed and given a trackable low-profile running wheel (Med Associates Inc.) to estimate the running behavior for the previous 11 weeks of the experiment. Running wheel rotations were measured in 1-min bins to allow for distance traveled (sum of rotations) calculated per mouse each night. Average rotations were calculated per mouse for each tracked night. Average speed while active was calculated by isolating the minute intervals where activity was measured, and averaging the number of rotations for the minutes active. Percent of time at each speed was calculated by totaling the number of minute bins that mice ran between 0, 1–30 rotations, 31–70 rotations, 71–100 rotations and 100+ rotations and dividing by the total amount of minutes tracked.
Nuclear magnetic resonance (NMR) imaging
The midlife running cohort was subjected to NMR imaging 1 week before harvest. Weight was taken and mice were briefly placed into a Plexiglas tube 2.5 in. by 8 in. which was then subjected to NMR (EchoMRI, Houston, TX). Magnetic field was measured by a 5-gauss magnet. Measurements included weight, lean muscle mass, fat mass, and water composition. Adiposity was calculated by (fat/body weight) × 100. Percentage lean muscle mass was calculated by (lean muscle mass/body weight) × 100.
Harvesting, tissue preparation and blood chemistry
All mice were euthanized by intraperitoneal injection of a lethal dose of Ketamine (100 mg/ml)/Xylazine (20 mg/ml) and blood was collected at harvest through approved cardiac puncture protocols. Mice were perfused intracardially with 1X PBS. Brains were carefully dissected and hemisected sagittally. Hippocampus (Hippo) and cortex (Ctx), were then carefully separated and snap frozen in solid CO2 for RNA-sequencing. Blood was also collected in K2 EDTA (1.0 mg) microtainer tubes (BD) at harvest (non-fasted) and kept at room temperature for at least 30 min to prevent clotting and then centrifuged at 22 °C for 15 min at 5000 rpm. Plasma was carefully collected and aliquoted. Plasma was characterized on the Beckman Coulter AU680 chemistry analyzer.
RNA and protein extraction, library construction and RNA sequencing
RNA sequencing (RNA-seq) was performed by The Jackson Laboratory Genome Technologies Core. RNA extraction involved homogenization with TRIzol (Invitrogen) as previously described [28]. RNA was isolated and purified using the QIAGEN miRNeasy mini extraction kit (QIAGEN) in accordance with manufacturer’s instructions. RNA quality was measured via the Bioanalyzer 2100 (Agilent Technologies) and poly(A) RNA-seq sequencing libraries were compiled by TruSeq RNA Sample preparation kit v2 (Illumina). Quantification was performed using qPCR (Kapa Biosystems). RNA-seq was performed on the HiSeq 4000 platform (Illumina) for 2x100bp reads for a total of 45 million reads according to the manufacturer’s instructions.
RNA-seq quality control and gene set enrichment
Quality control for each sample was completed using NGSQCToolkit v2.3 which removed adaptors and trimmed low quality bases (Phred< 30) [29]. To quantify gene expression of the trimmed reads, we used RSEM v1.2.12 which uses Bowtie2 v2.2.0 for alignment of these reads [30]. We used mouse genome mm-10 based upon the B6 reference genome. Differential gene expression (DGE) analysis was completed on the hippocampus and cortex separately, using EdgeR 3.20.9 [31]. A second quality control step included filtering out genes with less than at least 1 read per million for more than one sample. Normalization of trimmed mean of M values (TMM) was performed and quasi-likelihood F-test was used to attain DGE. Differentially Expressed genes (DE genes) were identified by comparing (i) young running to young sedentary, (ii) midlife low running to midlife sedentary, and (iii) midlife high running to midlife sedentary for each tissue (cortex or hippocampus). Genes were considered DE if the False Discovery Rate was less than 0.05 (FDR < 0.05). Given the young and midlife running were run at separate times it was not possible to directly compare the young data to the midlife data.
Ingenuity Pathway Analysis (IPA) was used to identify enriched canonical pathways for each DE gene list. Additionally, Database for Annotation, Visualization and Integrated Discovery (DAVID, v6.8) was used to identify enrichment of Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways and Gene Ontology (GO) terms, with the background gene sets being all trimmed normalized genes for each comparison. Enriched KEGG pathways and GO terms with FDR < 0.05 were considered significant. Cancer-related pathways were excluded from visualization.
Statistical analyses
Details of statistical analyses of RNA-seq data are provided above. All other statistical analyses were performed in GraphPad Prism v7.0a. Body composition and lipid profiling results between the young running and young sedentary cohorts utilized an unpaired t-test. Body composition and lipid profiling results between midlife sedentary, low runners, and high runners were compared with a one-way ANOVA.
Availability of data and materials
All RNA-seq data is available in GeoArchive under the accession number GSE132930. The submission process is in progress. ID numbers for accessing the data will be provided prior to acceptance of this manuscript and this statement will be amended accordingly.
Abbreviations
- AD:
-
Alzheimer’s Disease
- ANOVA:
-
Analysis of Variance
- B6:
-
C57BL/6 J
- BDNF:
-
Brain derived neurotrophic factor
- Ctx:
-
Cortex
- DAVID:
-
Database for Annotation, Visualization, and Integrated Discovery
- DE:
-
Differentially expressed
- DGE:
-
Differential gene expression
- ECM:
-
Extracellular matrix
- FDR:
-
False Discovery Rate
- GO:
-
Gene Ontology
- HDL:
-
High density lipoprotein
- Hippo:
-
Hippocampus
- IPA:
-
Ingenuity Pathway Analysis
- KEGG:
-
Kyoto Encyclopedia of Genes and Genomes
- LDL:
-
Low density lipoprotein
- logCPM:
-
Log (counts per million)
- mo:
-
Months
- NEFA:
-
Non-esterified fatty acids
- NMR:
-
Nuclear magnetic resonance
- RNA-seq:
-
RNA sequencing
- VCID:
-
Vascular Contributions to Cognitive Impairment and Dementia
References
Riedel BC, Thompson PM, Brinton RD. Age, APOE and sex: triad of risk of Alzheimer’s disease. J Steroid Biochem Mol Biol. 2016;160:134–47.
Chieffi S, Messina G, Villano I, Messina A, Valenzano A, Moscatelli F, et al. Neuroprotective effects of physical activity: evidence from human and animal studies. Front Neurol. 2017;8:188.
Stimpson NJ, Davison G, Javadi AH. Joggin’ the noggin: towards a physiological understanding of exercise-induced cognitive benefits. Neurosci Biobehav Rev. 2018;88:177–86.
Beard JR, Officer A, de Carvalho IA, Sadana R, Pot AM, Michel JP, et al. The world report on ageing and health: a policy framework for healthy ageing. Lancet. 2016;387(10033):2145–54.
Langa KM, Levine DA. The diagnosis and management of mild cognitive impairment: a clinical review. JAMA. 2014;312(23):2551–61.
Alzheimer’s A. 2016 Alzheimer’s disease facts and figures. Alzheimers Dement. 2016;12(4):459–509.
Coulthard EJ, Love S. A broader view of dementia: multiple co-pathologies are the norm. Brain. 2018;141(7):1894–7.
Kling MA, Trojanowski JQ, Wolk DA, Lee VM, Arnold SE. Vascular disease and dementias: paradigm shifts to drive research in new directions. Alzheimers Dement. 2013;9(1):76–92.
Norton S, Matthews FE, Barnes DE, Yaffe K, Brayne C. Potential for primary prevention of Alzheimer’s disease: an analysis of population-based data. Lancet Neurol. 2014;13(8):788–94.
van Praag H, Schinder AF, Christie BR, Toni N, Palmer TD, Gage FH. Functional neurogenesis in the adult hippocampus. Nature. 2002;415(6875):1030–4.
van Praag H, Shubert T, Zhao C, Gage FH. Exercise enhances learning and hippocampal neurogenesis in aged mice. J Neurosci. 2005;25(38):8680–5.
Middleton LE, Barnes DE, Lui LY, Yaffe K. Physical activity over the life course and its association with cognitive performance and impairment in old age. J Am Geriatr Soc. 2010;58(7):1322–6.
Greenwood BN, Strong PV, Foley TE, Fleshner M. A behavioral analysis of the impact of voluntary physical activity on hippocampus-dependent contextual conditioning. Hippocampus. 2009;19(10):988–1001.
Choi SH, Bylykbashi E, Chatila ZK, Lee SW, Pulli B, Clemenson GD, et al. Combined adult neurogenesis and BDNF mimic exercise effects on cognition in an Alzheimer’s mouse model. Science. 2018;361(6406):eaan8821.
Mancuso MR, Kuhnert F, Kuo CJ. Developmental angiogenesis of the central nervous system. Lymphat Res Biol. 2008;6(3–4):173–80.
Zhang Y, Chen K, Sloan SA, Bennett ML, Scholze AR, O'Keeffe S, et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J Neurosci. 2014;34(36):11929–47.
Vanlandewijck M, He L, Mae MA, Andrae J, Ando K, Del Gaudio F, et al. A molecular atlas of cell types and zonation in the brain vasculature. Nature. 2018;554(7693):475–80.
Khodadadi D, Gharakhanlou R, Naghdi N, Salimi M, Azimi M, Shahed A, et al. Treadmill exercise ameliorates spatial learning and memory deficits through improving the clearance of peripheral and central amyloid-beta levels. Neurochem Res. 2018;43(8):1561–74.
Tang Y, Zhang Y, Zheng M, Chen J, Chen H, Liu N. Effects of treadmill exercise on cerebral angiogenesis and MT1-MMP expression after cerebral ischemia in rats. Brain Behav. 2018;8(8):e01079.
Otsuka S, Sakakima H, Sumizono M, Takada S, Terashi T, Yoshida Y. The neuroprotective effects of preconditioning exercise on brain damage and neurotrophic factors after focal brain ischemia in rats. Behav Brain Res. 2016;303:9–18.
Lin TW, Tsai SF, Kuo YM. Physical exercise enhances neuroplasticity and delays Alzheimer’s disease. Brain Plast. 2018;4(1):95–110.
Bailey DM, Calverley TA, Corkill R. Drugs for dementia: exercise is medicine. BMJ. 2019;364:k5438.
Walchli T, Wacker A, Frei K, Regli L, Schwab ME, Hoerstrup SP, et al. Wiring the vascular network with neural cues: a CNS perspective. Neuron. 2015;87(2):271–96.
Darlington TM, McCarthy RD, Cox RJ, Miyamoto-Ditmon J, Gallego X, Ehringer MA. Voluntary wheel running reduces voluntary consumption of ethanol in mice: identification of candidate genes through striatal gene expression profiling. Genes Brain Behav. 2016;15(5):474–90.
Bell RD, Winkler EA, Singh I, Sagare AP, Deane R, Wu Z, et al. Apolipoprotein E controls cerebrovascular integrity via cyclophilin A. Nature. 2012;485(7399):512–6.
Iturria-Medina Y, Sotero RC, Toussaint PJ, Mateos-Perez JM, Evans AC, Alzheimer’s Disease Neuroimaging I. Early role of vascular dysregulation on late-onset Alzheimer’s disease based on multifactorial data-driven analysis. Nat Commun. 2016;7:11934.
Gorelick PB, Scuteri A, Black SE, Decarli C, Greenberg SM, Iadecola C, et al. Vascular contributions to cognitive impairment and dementia: a statement for healthcare professionals from the american heart association/american stroke association. Stroke. 2011;42(9):2672–713.
Soto I, Graham LC, Richter HJ, Simeone SN, Radell JE, Grabowska W, et al. APOE stabilization by exercise prevents aging neurovascular dysfunction and complement induction. PLoS Biol. 2015;13(10):e1002279.
Patel RK, Jain M. NGS QC toolkit: a toolkit for quality control of next generation sequencing data. PLoS One. 2012;7(2):e30619.
Langmead B, Trapnell C, Pop M, Salzberg SL. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009;10(3):R25.
Lun AT, Chen Y, Smyth GK. It’s DE-licious: a recipe for differential expression analyses of RNA-seq experiments using quasi-likelihood methods in edgeR. Methods Mol Biol. 2016;1418:391–416.
Acknowledgments
The authors wish to thank Todd Hoffert from Clinical Assessment Services for blood chemistry, Heidi Munger and the Genome Technologies group for RNA-sequencing, and Tim Stearns and Vivek Philip from Computational Sciences.
Funding
This work was supported by T32HD007065 to Kate Foley. Also, the authors are especially grateful to Tucker Taft and his wife Phyllis R. Yale, and the estate of Bennett Bradford and his daughter, Deborah Landry. Their generous and thoughtful support of Alzheimer’s research at The Jackson Laboratory supported this study. These funding sources supported study design, data collection and interpretation, and writing of the manuscript.
Author information
Authors and Affiliations
Contributions
KEF, LCG and GRH conceived and designed the project. KEF and LCG performed experiments. KEF and HSY performed bioinformatic analysis. KEF and GRH wrote this manuscript. All authors approved this manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
No human subjects or data was used in this study. All experiments involving mice were approved by the Animal Care and Use Committee at The Jackson Laboratory in accordance with guidelines set out in The Eighth Edition of the Guide for the Care and Use of Laboratory Animals. All euthanasia used methods were approved by the American Veterinary Medical Association.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1: Figure S1.
Midlife runners show variation in running speed and distance. Figure S2. Midlife High runners show reduced adiposity due to running. Figure S3. CPM scatterplot of young runners compared to young sedentary in the cortex and hippocampus. Figure S4. CPM scatterplot of midlife runners compared to midlife sedentary in the cortex and hippocampus. Figure S5. Fewer DE genes in the cortex and hippocampus of midlife run mice. Figure S6. Col4a1, Lamb2 and Lama5 are expressed in multiple cerebrovascular-related cells.
Additional file 2: Table S2.
All genes detected by RNA-seq, used as “background” gene list for gene set enrichment analyses.
Additional file 3: Table S3.
Differentially expressed genes in the cortex comparing MidlifeRunHigh to MidlifeSed.
Additional file 4: Table S4.
Differentially expressed genes in the hippocampus comparing MidlifeRunHigh to MidlifeSed.
Additional file 5: Table S5.
Differentially expressed genes in the cortex comparing MidlifeRunLow to MidlifeSed.
Additional file 6: Table S6.
Differentially expressed genes in the hippocampus comparing MidlifeRunLow to MidlifeSed.
Additional file 7: Table S7.
Differentially expressed genes in the cortex comparing YoungRun to YoungSed.
Additional file 8: Table S8.
Differentially expressed genes in the hippocampus comparing YoungRun to YoungSed.
Additional file 9: Table S1.
Housing of Midlife Running mice.
Additional file 10: Table S9.
Expression of Laminin and Collagen IV genes in both Zheng et al., and Vanlandewijck and He et al.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Foley, K.E., Yang, H.S., Graham, L.C. et al. Transcriptional profiling predicts running promotes cerebrovascular remodeling in young but not midlife mice. BMC Genomics 20, 860 (2019). https://doi.org/10.1186/s12864-019-6230-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-019-6230-z