
Abstract
Background
Self-incompatibility (SI) is a major barrier that obstructs the breeding process in most horticultural plants including tea plants (Camellia sinensis). The aim of this study was to elucidate the molecular mechanism of SI in tea plants through a high throughput transcriptome analysis.
Results
In this study, the transcriptomes of self- and cross-pollinated pistils of two tea cultivars ‘Fudingdabai’ and ‘Yulv’ were compared to elucidate the SI mechanism of tea plants. In addition, the ion components and pollen tube growth in self- and cross-pollinated pistils were investigated. Our results revealed that both cultivars had similar pollen activities and cross-pollination could promote the pollen tube growth. In tea pistils, the highest ion content was potassium (K+), followed by calcium (Ca2+), magnesium (Mg2+) and phosphorus (P5+). Ca2+ content increased after self-pollination but decreased after cross-pollination, while K+ showed reverse trend with Ca2+. A total of 990 and 3 common differentially expressed genes (DEGs) were identified in un-pollinated vs. pollinated pistils and self- vs. cross-pollinated groups after 48 h, respectively. Function annotation indicated that three genes encoding UDP-glycosyltransferase 74B1 (UGT74B1), Mitochondrial calcium uniporter protein 2 (MCU2) and G-type lectin S-receptor-like serine/threonine-protein kinase (G-type RLK) might play important roles during SI process in tea plants.
Conclusion
Ca2+ and K+ are important signal for SI in tea plants, and three genes including UGT74B1, MCU2 and G-type RLK play essential roles during SI signal transduction.
Similar content being viewed by others


Background
Self-incompatibility (SI) is a common phenomenon in plant reproduction system, which prevents self-fertilization in flowering plants. There are two classical known mechanisms for SI, namely, homomorphic gametophytic self-incompatibility (GSI) and homomorphic sporophytic self-incompatibility (SSI). In GSI system, the pollen incompatibility (haploid male gametophyte) is controlled by the S allele, pollen and pistils bearing the same S allele trigger an incompatible reaction [1]. While in SSI system, incompatibility is determined by both S alleles of the (diploid-sporophyte) pollen parents [2].
GSI has been found in many plant species, such as Solanaceae [3, 4] and Rosaceae [5,6,7], while SSI is typically found in Brassicaceae [8]. Both GSI and SSI have male or female determinate conditions which are regulated by different prominent genes [9]. In GSI system, S locus-encoded F-box (SLF/SFB) proteins control the pollen recognition of S-RNase based SI [10,11,12]. In SSI systems, S-locus receptor kinase (SRK) gene and S-locus cysteine-rich protein (SCR)/S-locus protein-11 (SP11) function as a receptor-ligand pair to recognize self-pollens at the surface of stigma epidermal papilla cells [13]. The SRK is a membrane-spanning receptor protein in stigma containing an extracellular domain (S-domain) for recognition of SP11, a transmembrane domain, and an intracellular serine/threonine kinase domain [14]. S-locus glycoprotein gene (SLG) and SRK exhibit series characteristics which are associated with the female determinant of SSI in Brassica [15]. The S domain of SRK is highly similar to the SLG, which is the first S-locus gene to be identified and a soluble glycoprotein secreted to the stigma surface [16, 17]. Besides, the pollen coat protein SCR/ SP11 controls pollen determinant of SSI in Brassica [8, 18], and many SP11, SRK, and SLG alleles were inherited together to term different S haplotypes.
Self-incompatibility mechanism remains unclear in tea plant. Previous studies suggested that tea plant SI might be in late-acting self-incompatibility system (LSI), in that self-pollinated pollen tubes elongated through the style but failed in fertilization [19, 20]. This has made it almost impossible to obtain fruits in self-pollinated tea plants (Camellia sinensis); thus, breeding process in tea plant is not encouraged. LSI is a novel SI system in plants, but the molecular mechanism of this system is still unclear. Recently, Zhang et al. [21] found that tea plant SI might be categorized to GSI through transcriptome analysis. Therefore, the SI mechanism in tea plants is still controversial and needs further exploration.
To understand the mechanism of SI in tea plant, the ion components and pollen tube growth in self- and cross-pollinated pistils were investigated. Furthermore, the transcriptome of self- and cross-pollinated pistils of two tea cultivars ‘Fudingdabai’ and ‘Yulv’ was compared to figure out the DEGs which may be involved in SI of tea plant. ‘Fudingdabai’ is a national superior clone and cultivated widely in China because of its good quality, high yield, and excellent stress resistance, while ‘Yulv’ is a high-quality cultivar selected from the hybrid offsprings of ‘Yabukita’. Both cultivars are self-incompatible and show high fruiting rates after cross-pollination. This study will provide reference for understanding SI mechanism of tea plant.
Methods
Plant materials and treatments
Two ten-year-old tea cultivars, namely, C. sinensis cv. Fudingdabai and C. sinensis cv. Yulv, cultivated in tea germplasm repository of Tea Research Institute of Fujian Academy of Agricultural Sciences were used in this study. Flowers from both of the two tea cultivars have three petals and trifid stigmas. The stigmas of ‘Fudingdabai’ divided at the base but ‘Yulv’ at the upper part (Fig. 1).

Morphology of flowers from ‘Fudingdabai’ and ‘Yulv’ cultivars
Flower buds of the two tea cultivars were harvested at 4:00 pm for pollens collection. Besides, the remaining flower buds of two cultivars were emasculated and used for artificial pollination next morning. A total of four pollination combinations were conducted: ‘Fudingdabai’ (♂) × ‘Fudingdabai’ (♀), ‘Yulv’ (♂) × ‘Yulv’ (♀), ‘Fudingdabai’ (♂) × ‘Yulv’ (♀), and Yulv’ (♂) × ‘Fudingdabai’ (♀), as shown in Fig. 2. The un-pollinated and pollinated pistils at 8, 24, 48 and 72 h were picked from each combination and frozen quickly in liquid nitrogen and stored at − 80 °C for RNA extraction. Three biological replicates were conducted with at least five pistils for each replicate.

The flow diagram of the experiment design. Red and green indicate pollination combination of the two tea cultivars in this study. The color dots mean pollens from the corresponding cultivars
Pollen culture in vitro
Pollen culture medium was prepared with the following substances: 0.59 g MES, 0.02 g H3BO3, 0.05 g Ca(NO3)·4H2O, 5 g sucrose and 5 g polyethylene glycol 4000 (PEG 4000), diluted with distilled water to 100 mL. Pollens were cultured in the medium in the dark and observed by Olympus light microscope (Olympus, Tokyo, Japan). Pollen germination rate and mean pollen tube length at 1, 2, and 4 h were calculated based on eight visual fields.
Fluorescence activity of pollen tube
Fresh un-pollinated and pollinated styles were fixed in FAA fixative buffer (5 mL formalin, 6 mL acetic acid, and 89 mL 50% ethanol) for 24 h [19]. The styles were washed with deionized water and then softened by 2 M NaOH overnight. The softened styles were stained by 0.1% aniline blue solution dissolved with 0.15 M K2HPO4. Finally, the styles were observed under Leica DM6B fluorescence microscope (Leica, Bannockburn, USA) after 15 min staining. At least five styles were observed for each sample.
Ion components of pistils
Un-pollinated and pollinated styles were dried at 80 °C for 4 h. A total of 0.1 g dried samples (at least 15 pistils) were ground and digested in 5 ml nitric acid by ETHOS One high performance microwave digestion system (Milestone, Bergamo, Italy) for 1 h. The digested samples were diluted with nitric acid to 25 mL and analyzed by inductively coupled plasma-optical emission spectrometer (PerkinElmer Optima 2100DV, Massachusetts, USA). A total of nine ions were detected, including potassium (K+), calcium (Ca2+), magnesium (Mg2+), phosphorus (P5+), zinc (Zn2+), boron (B3+), Ferrous (Fe2+), aluminium (Al3+), and manganese (Mn2+). The contents of the ions were quantified by establishing standard curve.
RNA extraction, library construction and sequencing
Total RNA was extracted using Plant RNA extraction kit (Bioteke, China) according to the manual. RNA quality and concentration were assessed by 1% agarose gels, Qubit®2.0 Fluorometer (Invitrogen, Carlsbad, USA), and Agilent Bioanalyzer 2100 system (Agilent, Palo Alto, USA). A total of 3 μg RNA per sample was used for sequencing libraries preparation by NEBNext®Ultra™ RNA Library Prep Kit for Illumina® (NEB, USA) following manufacturer’s instructions and library quality was assessed on the Agilent Bioanalyzer 2100 system (Agilent Technologies, Palo Alto, CA). The clustering of samples was performed using TruSeq PE Cluster Kit v3-cBot-HS (Illumina) according to the manufacturer’s instructions. Finally, sequencing analysis was carried out with an Illumina Hiseq 2500 platform to generate pair-end reads.
Genome alignment and gene annotation
Raw data of fastq format were processed, and then were cleaned by trimming the adapter sequences, ploy-A containing reads and low quality reads. The clean reads were aligned to the reference genome (http://www.plantkingdomgdb.com/tea_tree/) by TopHat2 using the default parameters [22]. The mapped reads were assembled into possible transcripts by Cufflinks [23]. The unannotated transcripts were annotated by BLAST [24] based on the following databases: NR (NCBI non-redundant protein sequences) [25], COG (Clusters of Orthologous Groups of proteins) [26], Swiss-Prot (A manually annotated and reviewed protein sequence database) [27], KEGG (Kyoto Encyclopedia of Genes and Genomes) and GO (Gene Ontology) [28].
Identification of differentially expressed genes
Fragments per Kilobase of transcript per Million mapped reads (FPKM) estimates produced by RNA-Seq by Cuffquant and Cuffnorm of Cufflinks was used to evaluate the expression of transcripts [29]. The transcriptome comparisons of un-pollinated vs. pollinated groups and self-pollinated vs. cross-pollinated groups were conducted to find the differentially expressed genes (DEGs). Differential expression analyses were performed using the DESeq R package 1.10.1 [30], which provides statistical routines to determine DEGs based on a negative binomial distribution model. The P values were adjusted by the Benjamini and Hochberg’s approach for controlling the false discovery rate [31]. False discovery rate < 0.01 and fold change > 2 was considered to be significantly differentially expressed. Pearson’s Correlation Coefficient was used to evaluate the correlation of biological repeats [30].
Quantitative real time PCR verification
The first-strand cDNA was synthesized from 1 μg of total RNA by using the RevertAid™ First Strand cDNA Synthesis Kit (Thermo Scientific, MA, USA) according to the manual. Quantitative real time PCR (QRT-PCR) was performed using SYBR Premix EX Taq (Takara, Japan) on Roche LightCycler® 480II (Switzerland) as instruction specified. The qRT-PCR primers (Table 1) were designed by using Primer Premier 5.0 (Premier Biosoft International, Palo Alto, CA). The GAPDH (GenBank: GE651107) from tea plant was used as the reference gene. All of the PCR reactions were conducted in triplicate and the average expression values were calculated. The relative expression level of each gene was calculated with the 2-ΔΔCT method [32].
Statistical analysis
The statistical analysis was conducted using Excel 2016 and GraphPad Prism 5.0 (San Diego, USA). The significance analysis of difference between two samples was evaluated with Student t-test and multiple comparisons were analyzed using One-way ANOVA and P < 0.05 was considered to be statistically significant. The results were displayed as mean ± standard deviation.
Results
Pollen germination in vitro and fluorescence of pollen tubes
In order to evaluate the pollen vitality, pollen germination rate and tube growth between ‘Fudingdabai’ and ‘Yulv’ were assessed and compared. As shown in Additional file 1: Figure S1A, the pollen appearance between two cultivars has no significant difference. Pollen germination rate and mean length of pollen tubes of the two cultivars were similar and increased gradually with prolongation of the growth time (Additional file 1: Figure S1B).
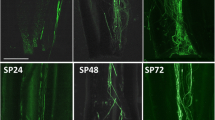
The fluorescence of pollen tube was observed to examine the growth of pollens in pistils (Additional file 2: Figure S2). After 8 h self-fertilization of ‘Fudingdabai’, the pollens germinated at stigma but no fluorescence was seen in styles. After 24 h, a few pollen tubes entered styles and the fluorescence on the base of styles was observed firstly after 48 h. Pollen tubes of ‘Yulv’ (♂) × ‘Fudingdabai’ (♀) cross-fertilized pistils showed higher growth rate than ‘Fudingdabai’ self-fertilization. The pollen tubes arrived at the base of styles after 24 and 8 h in self-fertilized ‘Yulv’ pistils and ‘Fudingdabai’ (♂) × ‘Yulv’ (♀) cross-fertilized pistils, respectively. Taken these results together, pollens from other cultivars would grow faster in pistils than that from themselves. This result was similar to that by Zhang et al. [21]. In addition, reciprocal cross-pollination showed that the pollen tube growth was slower when ‘Fudingdabai’ was used as maternal parent. However, pollen tubes in all of the self- and cross-pollinated samples reached the base of styles after 48 h.
Ion components in self- and cross-fertilized pistils
Ion components, especially Ca2+, are an indicator of self-incompatibility [33]. In tea pistils, the highest level of ion component observed in the tea pistil was K+, followed by Ca2+, Mg2+ and P5+ in sequence (Fig. 3a). Pistils of ‘Fudingdabai’ contained more K+ but less Ca2+ than those in ‘Yulv’. Ca2+ content in self-pollinated pistils of ‘Yulv’ (YLS) was higher than that in cross-pollinated pistils of ‘Yulv’ (YLC), but no apparent difference between self- (FDS) and cross-pollinated pistils of ‘Fudingdabai’ (FDC). In FDS pistils, the K+ content was higher than in FDC pistils, suggesting that Ca2+ and K+ may be involved in potential signal transduction in SI.

Ion components in self- and cross-pollinated pistils of tea plants. a 1 un-pollinated ‘Fudingdabai’ pistils; 2–5 self-pollinated ‘Fudingdabai’ pistils at 8 h, 24 h, 48 h and 72 h; 6–9 ‘Yulv’(♂) × ‘Fudingdabai’(♀) at 8 h, 24 h, 48 h and 72 h; 10 un-pollinated ‘Yulv’ pistils; 11–14 self-pollinated ‘Yulv’ pistils at 8 h, 24 h, 48 h and 72 h; 15–18 ‘Yulv’(♀) × ‘Fudingdabai’(♂) at 8 h, 24 h, 48 h and 72 h. b The Ca2+ and K+ content during self- and cross-pollinated pistils
Transcriptome assembly and function annotation
A total of 18 samples were sequenced and 122.75 Gb clean data were obtained. The percentages of clean reads having a base quality greater or equal than Q30 were above 85.01% indicating that the data produced by sequencing are of high quality. The clean reads from the 18 samples showed alignment ratios between 47.77% and 54.96% (SRA accession: SRP110788, Table 2). Based on the alignment with the reference genome of tea, 8136 unannotated genes were found, and 6621 of these genes were annotated after BLAST, with 1481 unigenes in COG database, 3465 in GO database, 2301 in KEGG database, 4285 in Swissprot database and 6588 in Nr database.
Differentially expressed genes analysis
Correlation analysis showed that T02 of self-pollinated ‘Fudingdabai’ at 48 h (FDS48) revealed low correlation to other two FDS48 samples (T07 and T12) with R2 of 0.33 and 0.43, respectively. T15 of self-pollinated ‘Yulv’ pistil sample at 48 h (YLS48) deviated from other two replicates (T05 and T10) with R2 of 0.46 and 0.52, respectively. These two samples (T02 and T15) were therefore removed in further DEG analysis. All replicates of remaining samples showed high correlation (Fig. 4).

Correlation analysis of the samples for differential expression analysis. Different sample numbers represent un-pollinated ‘Fudingdabai’ pistils (FD0), self-pollinated ‘Fudingdabai’ pistils at 48 h (FDS48), ‘Yulv’(♂) × ‘Fudingdabai’(♀) at 48 h (FDC48), un-pollinated ‘Yulv’ pistils (YL0), self-pollinated ‘Yulv’ pistils at 48 h (YLS48) and ‘Yulv’(♀) × ‘Fudingdabai’(♂) at 48 h (YLC48), respectively. Bold values are R2 for replicates of each sample
A total of 1948, 3399, 3927, 3682, 145, 2061, 1343, 600 and 1859 genes were found to be differentially expressed between each of un-pollinated ‘Fudingdabai’ pistils (FD0) vs. FDS48, FD0 vs. cross-pollinated ‘Fudingdabai’ (♀) pistils after 48 h (FDC48), un-pollinated ‘Yulv’ pistils (YL0) vs. YLS48, YL0 vs. cross-pollinated ‘Yulv’ (♀) pistils after 48 h (YLC48), FDS48 vs. FDC48, FDS48 vs. YLC48, YLS48 vs. FDC48, YLS48 vs. YLC48 and FDC48 vs. YLC48, respectively (Fig. 5a). By comparing the pollinated groups with un-pollinated groups, 990 common DEGs were found (Fig. 5b). COG classification of these DEGs showed that ‘General function prediction only’ enriched most of DEGs, followed by ‘Transcription’, ‘Signal transduction mechanisms’, ‘Replication, recombination and repair’ and ‘Secondary metabolites biosynthesis, transport and catabolism’ (Fig. 5c). GO enrichment analysis revealed that metabolic process in biological process, cell part in cellular component and catalytic activity in molecular function enriched the most DEGs (Additional file 3: Figure S3).

Differentially expressed genes identified in different comparisons. a The number of DEGs found in different comparisons. b, c Venn diagram and COG annotation for DEGs between un-pollinated and pollinated groups
Differentially expressed genes between self- and cross-pollinated groups
In comparison of self- and cross-pollinated groups, only three common DEGs were found (Fig. 6a). The 1160 common DEGs identified at least in two comparisons were therefore considered in further analysis. COG function classification revealed that ‘General function prediction only’ contained the most common DEGs followed by ‘Replication, recombination and repair’, ‘Transcription’ and ‘Signal transduction’. This result shows that cross-fertilization caused a series of responses in transcriptional level. In addition, the three common DEGs in all comparisons were UDP-glycosyltransferase 74B1 (UGT74B1, CSA001819), Mitochondrial calcium uniporter protein 2 (MCU2, CSA014152) and G-type lectin S-receptor-like serine/threonine-protein kinase RLK1 (G-lecRLK, Camellia_sinensis_newGene_13508). These genes showed similar expression patterns in un-pollinated and pollinated pistils. They also expressed at same levels during reciprocal cross-pollinations, but adversely expressed during self-pollinations of the two cultivars (Fig. 6b). Function annotation found that G-lecRLK was functioned on ‘Signal transduction mechanisms’. MCU2 worked on ‘Energy production and conversion’ and ‘Carbohydrate transport and metabolism’. Finally, UGT74B1 was annotated to ‘General function prediction only’.

DEGs identified between self- and cross-pollinated groups. a Venn diagram for DEGs between self- and cross-pollinated groups. b The expression of the three common DEGs between self- and cross-pollinated groups
In order to compare our results to the previous study [21], the data from self-pollinated (FDS48–1, SRR3290055) and cross-pollinated ‘Fudingdabai’ samples (FDC48–1, SRR3290084) at 48 h were downloaded and re-analyzed based on genome of tea plants. A total of 4262 DEGs were identified between FDS48–1 and FDC48–1 comparison. According to the large number of DEGs, we suggested that ‘Fudingdabai’ should be used as the paternal parent in the study of Zhang et al. [21]. As shown in Fig. 7, in comparison of self- and cross-pollinated groups (FDS48 vs. FDC48, FDS48 vs. YLC48 and FDS48–1 vs. FDC48–1), only five common DEGs were filtered. Therefore, the common DEGs identified at least in two comparisons were concerned. COG function classification revealed similar result to the four groups comparisons in our study (Fig. 7). Except for ‘general function prediction only’, the classes of ‘Replication, recombination and repair’, ‘Transcription’ and ‘Signal transduction’ enriched most of the DEGs. In addition, G-lecRLK, MCU2 and UGT74B1 were found in these common DEGs, suggesting that these DEGs played vital roles during SI process. In the five overlapping genes, MCU2 and UGT74B1 were found. G-lecRLK was only expressed in the comparisons in our study because it was annotated by database blast but not genome mapping.

Venn diagram and COG function classification for DEGs identified between self- and cross-pollinated groups. FDS48–1 and FDC48–1 are self- and cross pollinated samples from the study by Zhang et al.
DEGs between reciprocal cross-pollinations
In the present study, 1859 DEGs were identified in FDC48 vs. YLC48 comparison. Except for ‘General function prediction only’, most of these DEGs were involved in ‘Transcription’ and ‘Replication, recombination and repair’ (Fig. 8a). KEGG pathway enrichment analysis showed that ‘Galactose metabolism’ possessed the highest rich factor and ‘Phenylpropanoid biosynthesis’ had the most DEGs (Fig. 8b). This result suggests different responses between reciprocal cross-pollinations in tea cultivars.

The DEGs between reciprocal cross-pollinations. a COG function classification of DEGs between reciprocal cross-pollinations. b KEGG enrichment analysis of DEGs between reciprocal cross-pollinations
Verification of differentially expressed genes
In order to verify the reliability of RNA-Seq data, eight DEGs were selected for qRT-PCR analysis. As shown in Fig. 9, most of the DEGs showed similar expression trend compared to the RNA-Seq analysis. Therefore, the RNA-Seq analysis is credible.

The qRT-PCR verification for RNA-Seq analysis. * means different significantly (P < 0.05)
Discussion
Self-incompatibility is a common phenomenon in angiosperm. In order to understand the SI mechanism of tea plants, we studied the ion components and pollen growth in self- and cross-pollinated pistils from two tea cultivars. The results showed that pollen tubes grew faster in cross-pollinated pistils than those in self-pollinated pistils. Furthermore, Ca2+ in pistils increased after self-pollinations but decreased after cross-pollinations. In addition, comparative transcriptome analysis showed that G-type LecRLK, UGT74B1 and MCU2 genes might contribute the SI signal transduction mechanism in tea plant.
Signal transduction during self-incompatibility in tea plants
Self-incompatibility is genetically regulated by a multi-allelic S-locus which links pollen and pistil S-determinants and resulting in self-recognition. Interactions between pollen and pistil in the same haplotype triggered a SI response, which inhibits pollen tube growth and leads to failure of fertilization [34]. During SI process, a series of signal changes occurred in plants. The earliest identified physiological event caused by SI recognition is the increase of Ca2+ in incompatible pollen tubes or stigma papilla cells [35, 36]. In the present study, Ca2+ changes suggest a potential correlation between pollen tube growth and Ca2+ content. Furthermore, the opposing trend of Ca2+ ion content of the two cultivars between self- and cross-pollinated pistils reveals that Ca2+ may be an important signal for SI in tea plants.
In the present study, a DEG MCU2 was identified between self- and cross-pollination, which undertook the mitochondrial Ca2+ uptake [37]. In animals, Ca2+ uptake could regulate the mitochondrial energy production that is a stimulation of sperm-induced Ca2+ release [38]. There is evidence that the Ca2+ uptake also occurred at fertilization in mammalian eggs [39]. Inhibition of the mitochondrial function also disrupted the sperm-induced Ca2+ oscillatory pattern and intracellular Ca2+ homeostasis, and resulted in low developmental competence in mammals [40]. Unlike in animals, the functional mechanism of MCU in tea plants has been less studies, and; therefore, needs verification except for Ca2+, K+ is also sensitive to SI. In Papaver rhoeas, conductance of some monovalent cations, such as K+ and NH4+ were also stimulated by SI [41]. Interestingly, content of K+ changes was opposite to Ca2+ after pollination (Fig. 3). We can therefore propose that SI activates a nonspecific ion channel in tea plants.
Role of self-incompatibility related genes in tea plants
LecRLK family has been classified to three subfamilies: L-type, G-type and C-type LecRLKs. This classification is supported by the structure analysis of these proteins. L-type LecRLK contains a legume lectin-like extracellular domain, and G-type LecRLK has a α-mannose binding bulb lectin domain, while C-type LecRLKs are characterized due to the presence of calcium-dependent carbohydrate-binding domain [42]. G-type LecRLKs were historically known as SRKs, since they hold the D-mannose binding lectin (B_lectin) and catalytic domain of the serine/threonine kinases.
SRK genes have a S domain which is responsible for SI in Brassicaceae [15, 43]. Recently, these genes were also reported to confer abiotic stress tolerance and delay dark-induced leaf senescence in rice [44]. Here, we screened a similar SRK gene (Camellia_sinensis_newGene_13508) from tea plant which differently expressed between self- and cross-pollinated pistils and might contribute to the signal transduction of SI in tea plant. In general, the SRK genes function in SI through the diversity of S domain. Therefore, the S domain of the SRK gene should be identified in different tea cultivars to explore the role of SRK on SI process of tea.
The previous studies have identified a LSI or an ovarian sterility (OS) type controlling self-incompatibility in tea plants [19, 20]. The same phenomenon was also observed in our study. However, the molecular mechanism of this SI system remained unclear until Zhang et al. [21, 45] proposed a gametophytic SI mechanism based on S-RNase control in tea plant. Unexpectedly, S-RNase gene was not found in tea plant in the present study, but three DEGs were identified in comparison between self- and cross-pollinated pistils: G-type LecRLK, MCU2 and UGT74B1. Pollen tube reception, the crosstalk between the male and female gametophytes when pollen tubes arrive at the synergid cells of the ovule in flowering plants, mutation of TURAN(TUN) and EVAN(EVN) genes led to overgrowth of the pollen tubes inside the female gametophyte and inhibited the rupture of pollen tubes. TUN encodes a UGT superfamily protein and is required for pollen tube growth and integrity by affecting the stability of the pollen-specific FERONIA RLKs [46, 47]. In this work, whether the UGT74B1 and G-type RLK genes work together on fertilization in tea plant remains unknown. Nevertheless, we can suggest that both of the genes may codetermine the SI mechanism in tea plant.
It is difficult to explain that G-type LecRLK, MCU2 and UGT74B1 showed so different expression patterns between self-pollinations. It may be due to the variety difference of tea plants. More cultivars should be adopted to detect the expression of these genes in self- and cross-pollinations to interpret their roles in SI. Besides, function analysis through transgenic test to clarify the mechanism of these two genes in SI will be a good way in the future if more studies can be conducted to overcome the barriers in tea plant transformation. Our study suggests a distinctive mode of action of SI in tea, and the results therein provide new guidance and reference for exploration of SI mechanism in tea.
Conclusion
The present study revealed that cross-pollination could promote the growth of pollens in styles and Ca2+ and K+ are involved in signal transduction in SI process of tea plants, and also G-type LecRLK and UGT74B1 may function together in controlling SI in tea plants. However, the specific role of these genes in SI process needs further identification. Our study will help understand the SI mechanism of tea plant further.
Abbreviations
- DEGs:
-
Differentially expressed genes
- GSI:
-
Gametophytic self-incompatibility
- G-type RLK:
-
G-type lectin S-receptor-like serine/threonine-protein kinase
- LSI:
-
Late-acting self-incompatibility system
- MCU2:
-
Mitochondrial calcium uniporter protein 2
- SI:
-
Self-incompatibility
- SLG:
-
S-Locus glycoprotein gene
- SRK:
-
S-Locus receptor kinase
- SSI:
-
Sporophytic self-incompatibility
- UGT74B1:
-
UDP-glycosyltransferase 74B1
References
Newbigin E, Anderson MA, Clarke AE. Gametophytic self-incompatibility systems. Plant Cell. 1993;5(10):1315.
Charlesworth D. Evolution of homomorphic sporophytic self-incompatibility. Heredity. 1988;60(3):445–53.
McClure BA, Gray JE, Anderson MA, Clarke AE. Self-incompatibility in Nicotiana alata involves degradation of pollen rRNA. Nature. 1990;347(6295):757–60.
Li W, Chetelat RT. The role of a pollen-expressed Cullin1 protein in gametophytic self-incompatibility in solanum. Genetics. 2014;196(2):439–42.
Tao R, Yamane H, Sassa H, Mori H, Gradziel TM, Dandekar AM, Sugiura A. Identification of stylar RNases associated with gametophytic self-incompatibility in almond (Prunus dulcis). Plant Cell Physiol. 1997;38(3):304–11.
Mariette S, Tavaud M, Arunyawat U, Capdeville G, Millan M, Salin F. Population structure and genetic bottleneck in sweet cherry estimated with SSRs and the gametophytic self-incompatibility locus. BMC Genet. 2010;11(1):77.
Vieira J, Ferreira PG, Aguiar B, Fonseca NA, Vieira CP. Evolutionary patterns at the RNase based gametophytic self-incompatibility system in two divergent Rosaceae groups (Maloideae and Prunus). BMC Evol Biol. 2010;10(1):200.
Schopfer CR, Nasrallah ME, Nasrallah JB. The male determinant of self-incompatibility in Brassica. Science. 1999;286(5445):1697–700.
Takayama S, Isogai A. Self-incompatibility in plants. Annu Rev Plant Biol. 2005;56:467–89.
Sijacic P, Wang X, Skirpan AL, Wang Y, Dowd PE, McCubbin AG, Huang S. Kao T-h. Identification of the pollen determinant of S-RNase-mediated self-incompatibility. Nature. 2004;429(6989):302–5.
Qiao H, Wang F, Zhao L, Zhou J, Lai Z, Zhang Y, Robbins TP, Xue Y. The F-box protein AhSLF-S2 controls the pollen function of S-RNase–based self-incompatibility. Plant Cell. 2004;16(9):2307–22.
Williams JS, Natale CA, Wang N, Li S, Brubaker TR, Sun P. Kao T-h. Four previously identified Petunia inflata S-locus F-box genes are involved in pollen specificity in self-incompatibility. Mol Plant. 2014;7(3):567–9.
Kachroo A, Schopfer CR, Nasrallah ME, Nasrallah JB. Allele-specific receptor-ligand interactions in Brassica self-incompatibility. Science. 2001;293(5536):1824–6.
Stein JC, Howlett B, Boyes DC, Nasrallah ME, Nasrallah JB. Molecular cloning of a putative receptor protein kinase gene encoded at the self-incompatibility locus of Brassica oleracea. PNAS. 1991;88(19):8816–20.
Takasaki T, Hatakeyama K, Suzuki G, Watanabe M, Isogai A, Hinata K. The S receptor kinase determines self-incompatibility in Brassica stigma. Nature. 2000;403(6772):913–6.
Nasrallah JB, Nasrallah ME. Pollen-stigma signaling in the Sporophytic self-incompatibility response. Plant Cell. 1993;5(10):1325.
Nasrallah JB, Yu S-M, Nasrallah ME. Self-incompatibility genes of Brassica oleracea: expression, isolation, and structure. PNAS. 1988;85(15):5551–5.
Takayama S, Shiba H, Iwano M, Shimosato H, Che F-S, Kai N, Watanabe M, Suzuki G, Hinata K, Isogai A. The pollen determinant of self-incompatibility in Brassica campestris. PNAS. 2000;97(4):1920–5.
Chen X, Hao S, Wang L, Fang W, Wang Y, Li X. Late-acting self-incompatibility in tea plant (Camellia sinensis). Biologia. 2012;67(2):347–51.
Wachira F, Kamunya S. Pseudo-self-incompatibility in some tea clones (Camellia sinensis (L.) O. Kuntze). J Hort Sci Biotech. 2005;80(6):716–20.
Zhang C-C, Wang L-Y, Wei K, Wu L-Y, Li H-L, Zhang F, Cheng H, Ni D-J. Transcriptome analysis reveals self-incompatibility in the tea plant (Camellia sinensis) might be under gametophytic control. BMC Genomics. 2016;17(1):359.
Trapnell C, Roberts A, Goff L, Pertea G, Kim D, Kelley DR, Pimentel H, Salzberg SL, Rinn JL, Pachter L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and cufflinks. Nat Protoc. 2012;7:562.
Ghosh S, Chan C. Analysis of RNA-Seq data using TopHat and cufflinks. Methods Mol Biol. 2015;1374:339–61.
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997;25(17):3389–402.
Deng Y, Li J, Wu S, Zhu Y, Chen Y, He F. Integrated nr database in protein annotation system and its localization. Comput Eng. 2006;32(5):71–4.
Tatusov RL, Galperin MY, Natale DA, Koonin EV. The COG database: a tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000;28(1):33–6.
Apweiler R, Bairoch A, Wu CH, Barker WC, Boeckmann B, Ferro S, Gasteiger E, Huang H, Lopez R, Magrane M. UniProt: the universal protein knowledgebase. Nucleic Acids Res. 2004;32(1):D115–9.
Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT. Gene ontology: tool for the unification of biology. Nat Genet. 2000;25(1):25–9.
Florea L, Song L, Salzberg SL. Thousands of exon skipping events differentiate among splicing patterns in sixteen human tissues. F1000Research. 2013;2:188.
Anders S, Huber W. Differential expression analysis for sequence count data. Genome Biol. 2010;11(10):R106.
Reiner A, Yekutieli D, Benjamini Y. Identifying differentially expressed genes using false discovery rate controlling procedures. Bioinformatics. 2003;19(3):368–75.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001;25(4):402–8.
Franklin-Tong N. Self-incompatibility: calcium signalling in Brassica. Nat Plants. 2015;1:15129.
Eaves DJ, Flores-Ortiz C, Haque T, Lin Z, Teng N, Franklin-Tong VE. Self-incompatibility in Papaver: advances in integrating the signalling network: Portland Press Limited; 2014.
Iwano M, Ito K, Fujii S, Kakita M, Asano-Shimosato H, Igarashi M, Kaothien-Nakayama P, Entani T, Kanatani A, Takehisa M. Calcium signalling mediates self-incompatibility response in the Brassicaceae. Nat Plants. 2015;1:15128.
Jiang X, Gao Y, Zhou H, Chen J, Wu J, Zhang S. Apoplastic calmodulin promotes self-incompatibility pollen tube growth by enhancing calcium influx and reactive oxygen species concentration in Pyrus pyrifolia. Plant Cell Rep. 2014;33(2):255–63.
Kirichok Y, Krapivinsky G, Clapham DE. The mitochondrial calcium uniporter is a highly selective ion channel. Nature. 2004;427(6972):360–4.
Campbell K, Swann K. Ca2+ oscillations stimulate an ATP increase during fertilization of mouse eggs. Dev Biol. 2006;298(1):225–33.
Dumollard R, Duchen M, Sardet C. Calcium signals and mitochondria at fertilisation. In: Seminars in cell & developmental biology: 2006: Elsevier; 2006. p. 314–23.
Miao Y-L, Williams CJ. Calcium signaling in mammalian egg activation and embryo development: the influence of subcellular localization. Mol Rep Dev. 2012;79(11):742–56.
Wu J, Wang S, Gu Y, Zhang S, Publicover SJ, Franklin-Tong VE. Self-incompatibility in Papaver rhoeas activates nonspecific cation conductance permeable to Ca2+ and K+. Plant Physiol. 2011;155(2):963–73.
Vaid N, Macovei A, Tuteja N. Knights in action: lectin receptor-like kinases in plant development and stress responses. Mol Plant. 2013;6(5):1405–18.
Goring DR, Rothstein SJ. The S-locus receptor kinase gene in a self-incompatible Brassica napus line encodes a functional serine/threonine kinase. Plant Cell. 1992;4(10):1273–81.
Chen L-J, Wuriyanghan H, Zhang Y-Q, Duan K-X, Chen H-W, Li Q-T, Lu X, He S-J, Ma B, Zhang W-K. An S-domain receptor-like kinase, OsSIK2, confers abiotic stress tolerance and delays dark-induced leaf senescence in rice. Plant Physiol. 2013;163(4):1752–65.
Zhang C-C, Tan L-Q, Wang L-Y, Wei K, Wu L-Y, Zhang F, Cheng H, Ni D-J. Cloning and characterization of an S-RNase gene in Camellia sinensis. Sci Hortic. 2016;207:218–24.
Lindner H, Kessler SA, Müller LM, Shimosato-Asano H, Boisson-Dernier A, Grossniklaus U. TURAN and EVAN mediate pollen tube reception in Arabidopsis Synergids through protein glycosylation. PLoS Biol. 2015;13(4):e1002139.
Escobar-Restrepo J-M, Huck N, Kessler S, Gagliardini V, Gheyselinck J, Yang W-C, Grossniklaus U. The FERONIA receptor-like kinase mediates male-female interactions during pollen tube reception. Science. 2007;317(5838):656–60.
Funding
This work was supported by the National Natural Science Foundation of China (31470690, 31570689) and the China Earmarked Fund for Modern Agro-industry Technology Research System (CARS-19). The funders had no role in study design, data collection, analysis and interpretation, or preparation of the manuscript.
Availability of data and materials
The datasets generated and/or analysed during the current study are available in the Sequence Read Archive (https://www.ncbi.nlm.nih.gov/sra/) with the accession number of SRR3290055.
Author information
Authors and Affiliations
Contributions
QM performed pollination experiment, RNA extraction, RNA-Seq analysis and drafted the manuscript. CC cultivated the tea plants and participated in pollination experiment and samples collection. ZZ was involved in samples collection and detected the pollen activity of tea flowers. ZZ contributed to samples collection and observation of pollen tubes fluorescence. HL conducted the ion assay and edited the figures in the manuscript. QZ carried out qRT-PCR experiment. XC was involved in the study design and RNA-Seq analysis. KS performed statistical analysis and participated in the revision and finalization of the manuscript. XL conceived of the study, participated in its design and coordination, and helped to draft the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Tea plants used in this study were cultivated in tea germplasm repository of Tea Research Institute of Fujian Academy of Agricultural Sciences (FAAS). This study was permitted by FAAS and conducted in accordance with stipulations of FAAS.
Competing interests
The authors declare that they have no competing financial interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1:
Figure S1. Pollen appearance and activity between ‘Fudingdabai’ and ‘Yulv’ in vitro. A Pollen germination and phenotype of ‘Fudingdabai’ and ‘Yulv’. B Pollen germination rate and average length of pollen tubes of ‘Fudingdabai’ and ‘Yulv’. (TIF 1137 kb)
Additional file 2:
Figure S2. Fluorescence of pollen tubes in self- and cross-pollinated pistils of tea plants at 48 h. “Top” and “Base” means the stigma and the base of the style of tea flower, respectively. Arrows indicate the pollen tubes with fluoresce. (TIF 4517 kb)
Additional file 3:
Figure S3. Gene Ontology enrich analysis of DEGs between unpollinated and pollinated samples. (TIF 755 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Ma, Q., Chen, C., Zeng, Z. et al. Transcriptomic analysis between self- and cross-pollinated pistils of tea plants (Camellia sinensis). BMC Genomics 19, 289 (2018). https://doi.org/10.1186/s12864-018-4674-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-018-4674-1