
Abstract
Background
B-cell chronic lymphocytic leukemia (CLL) is a common type of adult leukemia. It often follows an indolent course and is preceded by monoclonal B-cell lymphocytosis, an asymptomatic condition, however it is not known what causes subjects with this condition to progress to CLL. Hence the discovery of prediagnostic markers has the potential to improve the identification of subjects likely to develop CLL and may also provide insights into the pathogenesis of the disease of potential clinical relevance.
Results
We employed peripheral blood buffy coats of 347 apparently healthy subjects, of whom 28 were diagnosed with CLL 2.0–15.7 years after enrollment, to derive for the first time genome-wide DNA methylation, as well as gene and miRNA expression, profiles associated with the risk of future disease. After adjustment for white blood cell composition, we identified 722 differentially methylated CpG sites and 15 differentially expressed genes (Bonferroni-corrected p < 0.05) as well as 2 miRNAs (FDR < 0.05) which were associated with the risk of future CLL. The majority of these signals have also been observed in clinical CLL, suggesting the presence in prediagnostic blood of CLL-like cells. Future CLL cases who, at enrollment, had a relatively low B-cell fraction (<10%), and were therefore less likely to have been suffering from undiagnosed CLL or a precursor condition, showed profiles involving smaller numbers of the same differential signals with intensities, after adjusting for B-cell content, generally smaller than those observed in the full set of cases. A similar picture was obtained when the differential profiles of cases with time-to-diagnosis above the overall median period of 7.4 years were compared with those with shorted time-to-disease. Differentially methylated genes of major functional significance include numerous genes that encode for transcription factors, especially members of the homeobox family, while differentially expressed genes include, among others, multiple genes related to WNT signaling as well as the miRNAs miR-150-5p and miR-155-5p.
Conclusions
Our findings demonstrate the presence in prediagnostic blood of future CLL patients, more than 10 years before diagnosis, of CLL-like cells which evolve as preclinical disease progresses, and point to early molecular alterations with a pathogenetic potential.
Similar content being viewed by others

Background
B-cell chronic lymphocytic leukemia (CLL) is the most common adult leukemia in the Western world, accounting for roughly 30% of all leukemias and with incidence rates in different countries ranging between <1 and 5.5 cases per 100,000 [1]. It is characterized by the presence of large numbers (>5000/μl) of clonal, mature B-cells with increased proliferation and prolonged survival and expressing a number of specific surface markers, including CD5, CD19 and CD23. The disease often follows an indolent course, with many patients having no symptoms at diagnosis or surviving for decades after diagnosis without any need for treatment [2]. Nevertheless, despite great progress in therapeutic protocols, CLL remains an incurable disease whose etiology is largely unknown, and some patients have recurrent relapses requiring several lines of treatment [3].
It is well established that CLL is preceded by monoclonal B-cell lymphocytosis (MBL) [4], an asymptomatic hematological condition characterized by the presence of small clones of B-cells, whose prevalence in the general population ranges from less than 1% to nearly 20%, depending on the sensitivity of the detection methods employed [5]. Τwo types of the MBL are recognized, a) low-count MBL (<500 clonal B-cells /μl), which appears to remain stable with near-zero risk of progression to CLL, and b) high-count (or clinical) MBL (500–5000 clonal B-cells /μl), which is believed to be a precursor of CLL with 1–2% of its carriers progressing to clinically defined CLL per year. Currently it is not known what causes some MBL carriers to progress to CLL, and the number of clonal B-cells appears to constitute the best marker predictive of such progression [6, 7]. A number of biomarkers which are present in CLL cells and have prognostic value in relation to the clinical progression or therapeutic response of the disease (e.g. levels of expression of CD38 or ZAP-70, deletions in chromosomes 11, 13 and 17, trisomy 12, mutations in the IgG hypervariable region, mutations in CLL driver genes such as NOTCH1, BIRC3 or SF3B1) are also observed in MBL and help to identify MBL patients who are more likely to ultimately develop advanced CLL affecting survival [8, 9]. However such markers of prognosis appear to be of limited value with regard to the prediction of the risk of MBL subjects making the transition to a CLL phenotype [10, 11].
During the past few years a number of studies reported on the ability of various biomarkers measured in the blood of apparently healthy subjects to predict the risk of future diagnosis of CLL [12,13,14,15,16]. These predictive biomarkers concern cell surface markers and mitochondrial DNA copy number and provide limited insight into cellular processes that precede clinical disease. On the other hand, we recently reported that a gene expression profile measured in peripheral blood leukocytes could identify with high accuracy individuals who were diagnosed with CLL 2–17 years later [17] and highlighted genes whose expression was deregulated long before disease diagnosis. In extended analyses we showed that these gene expression profiles showed marked overlap with expression profiles of clinical CLL samples [18] suggesting that circulating cells long before diagnosis harbor CLL-traits. A common limitation of these studies lies in the lack of information regarding the hematological status of the study subjects at recruitment, including the presence or not of undiagnosed CLL. Here we report on the genome-wide epigenomic (DNA methylation) and miRNA expression profiles in peripheral blood leukocytes of the same population as in our above-mentioned study and the identification of prediagnostic epigenetic profiles which predict with high efficiency the risk of future CLL. Importantly, in the current study we utilized DNA methylation profiles, obtained using the Illumina HumanMethylation450 platform, to estimate for each subject the proportions of different subpopulations of white blood cells (WBC) and were therefore able to derive CLL risk-associated profiles, including a revised genome-wide gene expression profile, adjusted for WBC composition. The results reported here offer insights into the evolution of CLL and provide a basis for the development of improved prediagnostic markers predictive of the risk of future development of CLL.
Methods
Population
The study was conducted in the context of the European EnviroGenomarkers project [19] and involved subjects from the European Prospective Investigation into Cancer and Nutrition study (EPIC-ITALY) and the Northern Sweden Health and Disease Study (NSHDS) (Table 1). Both studies used population-based recruitment with standardized lifestyle and personal history questionnaires, anthropometric data and blood samples collected at recruitment (1993–1998 for EPIC-ITALY; 1990–2006 for NSHDS). Buffy coats were isolated from the collected blood samples and placed in long-term cold storage. Although the EnviroGenomarkers project was originally designed as two nested case-control studies, one for B-cell lymphoma and one for breast cancer [17], in the presently reported study subjects who eventually developed disease other than CLL were excluded, leaving 28 subjects who developed CLL (cases) and 319 subjects who remained apparently healthy until the end of the observation period (controls). No participant was diagnosed with disease within less than 2 years of blood sample collection and for this reason all participants were treated as apparently healthy at recruitment. In order to minimize the effects of sample handling on the omic profiles, subjects were included in the current study only if, at recruitment, the processing of their blood samples and freezing of the buffy coats had been completed within 2 h of collection [20].
Estimation of WBC from DNA methylation profiles
WBC composition was estimated using a published algorithm [21] and DNA methylation data derived from purified normal blood cell sub-populations (CD4-, CD8-, T- and NK cells, monocytes, granulocytes) publicly available in the FlowSorted.Blood.450 k Bioconductor package, as previously described [22]. The applicability of this methodology to the estimation of the WBC composition in CLL patients was recently confirmed [23]. To evaluate further the reliability of this method with CLL blood we applied it to published 450 k methylation data of purified (>95%) CLL cells isolated from the blood of 139 patients as well as 26 samples of normal B-cells, available from the International Cancer Genome Consortium [24]. The proportions of B cells, as estimated via DNA methylation, showed a single distribution as evaluated by the maximum BIC criterion, with a mean value of 88.5% (variance = 0.4%; SD 6.6%), while all other cell types gave mean values below 1.8% except CD4 which gave a mean value of 7.1% (results not shown). The accuracy of these estimates is comparable to that exhibited by the same methodology in normal blood [21].
Analytical procedures and data processing
RNA and DNA extraction from buffy coats, genome-wide analysis of gene expression (Agilent 4 × 44 K human whole genome microarray platform), CpG methylation (Illumina Infinium HumanMethylation450 platform) and miRNA expression profiling [Agilent Human miRNA Microarray (Release 19.0, 8x60K), representing 2006 human miRNAs], were conducted as previously described [17, 20, 25]. Methylation data were preprocessed initially with GenomeStudio (version 2011.1) Methylation module (version 1.9; Illumina). Subsequently, data normalization to address the issue of unwanted technical variation was performed, using scripts written and ran in MATLAB environment (Mathworks, Release 2012b), making use of the DNA methylation measured in multiple replicates of a technical control sample randomly distributed among the study samples and utilising procedure involving two successive steps of intensity-based correction (a) within-chip and b) across all probes) as previously described [26]. Probes with background signal (p < 0.01) in more than 10% of the samples were filtered out. Probes containing SNPs at a distance less than 3 nucleotides from the interrogated CpG cytosine and minor allele frequency > 10% were also omitted as well as probes giving mean methylation for all samples in the range 0- 4% or 96–100%. Missing values imputation (k-nearest neighbor) was applied to the resulting final number of 396,808 target CpG sites. Methylation levels were expressed as M-values corresponding to the logarithmic ratio of the methylated versus the unmethylated signal intensities.
All unsupervised analyses (PCA, clustering) were performed using the denoised signals, correcting for batch effect (date of chip analysis for the epigenetics and date of hybridization for transcriptomics), gender, cohort and smoking status. Use of the batch removal processes built in the Combat in R (version 3.0.2) and the ArrayStudio (Omicsoft, Cary, NC, USA, version 8.0.1.32) software packages gave very similar results, and consequently the batch removal tool of Arraystudio was adopted for further analyses.
Statistical analyses
Generalized linear models (GLM) using the batch-corrected signals (date of chip analysis for the DNA methylation and date of hybridisation for gene expression data), as well as Linear Mixed Models (LMM) using as random variables those mentioned above, were applied using the ArrayStudio software package. M values for DNA methylation or log2 intensities of mRNA or miRNA expression were the dependent variables, CLL status the independent variable, while as confounder variables we included sex, age, BMI, cohort as well as the six cell type fractions (CD4, CD8, NK cells, monocytes, B-cells, granulocytes). GLM and LMM gave very similar results and consequently, in order to reduce the possibility of overfitting, the GLM was finally adopted and five out of the total six of the cell type fractions (excluding granulocytes) were included as confounders. The estimated adjusted effect sizes are expressed as the least square means (LSM) (also known as EMM - estimated marginal means) which, in an analysis of covariance model, correspond to the group means after having controlled for a covariate [27]. The LSM β values were derived from the corresponding estimated LSM M values and the equation β = 2M/(1 + 2M). Multiple testing was accounted for with high stringency by using Bonferroni or FDR Benjamini-Hochberg correction.
The selection of CpGs with minimal variation between WBC subtypes was based on the data by Jaffe and Irizarry [22], selecting sites which fulfill the following criteria: a) p > 0.00012 (1000fold greater than the raw p-value corresponding to Bonferoni-corrected p < 0.05) and b) coefficient of variation (CV) <5% across all leukocyte subpopulations (CD4-, CD8-, T- and NK cells, monocytes, granulocytes). PCA was performed using the denoised values of 1308 out of the 10,785 CLL risk-associated CpG (FDR < 0.05) and 1308 CpGs with FDR >0.8 (control) which fulfill the above mentioned criteria. The CV of the CLL risk- associated CpG sites thus selected for the PCA analysis ranged 0.13–5% (mean = 1.96%).
Non-negative matrix factorization (NMF) was performed in Arraystudio using the default parameters maximum iteration n = 100 and stopping rule 1X10−6, specifying the number of clusters as 2–4.
Bioinformatics analysis
Gene functional classification analysis was performed using the DAVID bioinformatics tool [28] (default DAVID values, low stringency criteria). Functional analysis of genes associated with DM sites or DE probes was performed using the BioinfoMiner web application, which enables systemic, functional interpretation of omic datasets through the exploitation of various biomedical ontologies, extracting highly enriched gene sets that form cross-talking functional clusters [29]. BioinfoMiner initially maps the input omic data at the gene level and subsequently, through a combination of advanced statistical and network topological criteria, probabilistically prioritizes the resulting genes according to their functionality by comparison with enrichments of random resamplings, thus facilitating the identification and rejection of false positives. The non-parametric, empirical nature of this prioritization approach permits its generic, broad applicability even to classes of statistical testing problems that deflect from traditional hypotheses, as is the case for DNA methylation profiles, ensuring robust performance. Pathway analysis with this tool exploits variations of the StRAnGER and GOrevenge algorithms [30], so that molecular information (functions, processes, cellular compartments) is highlighted according to multiple criteria (enrichment score, expression etc.) while in addition regulatory hub genes which play a pivotal role in the phenotype under study are identified. Differentially methylated or expressed genes were used as input to identify statistically significantly over-represented terms from four different ontologies: Gene Ontology, Human Phenotype Ontology, MGI Mammalian Phenotype Ontology, as well as Reactome pathways Ontology. For the KEGG pathways analysis part the original StRAnGER2 web service was used [29].
ROC analysis
A variety of classification algorithms were tested for their performance, including SVM with linear kernel, SVM with Gaussian kernel, Bayesian generalized linear regression, Naive Bayes classifier, random forest and k-NN optimized in two different sets of k options. These classifiers were trained on a subset of our data corresponding to 50% of the study subjects (“training set”). To eliminate the effect of the severely unbalanced training set with respect to the proportion of classes (8.5% cases, 91.5% controls), we implemented the ROSE algorithm from the ‘ROSE’ R package [31]. Thus we trained the classifiers using the ROSE-derived balanced data (consisting of the same number of samples as the original training set, but 46.4% cases and 53.6% controls). The performance of the classifiers was assessed by the AUC value of the resulting ROC curves when the rest of the data (the other 50% of the original dataset) were used as a “testing set”. For the implementation of the methodology described above the ‘caret’ R package was used, choosing also a 4-fold cross-validation scheme repeated 1000 times. Subsequently, a recursive feature elimination algorithm, with maximum number of predictors set to 40, was used to identify an optimal subset of predictors for each of 3 best-performing classifiers chosen.
Results
CLL risk-related profiles
First we employed the genome-wide DNA methylation profiles to estimate for each study subject the fraction of 6 major cell sub-populations (CD4-, CD8-, T- and NK cells, monocytes, granulocytes) among all WBCs. The main difference observed between case and control subjects was a large (on average 3.1fold) increase in the fraction of B-cells in cases (for details see Additional file 1: Text). As shown in Additional file 2 Table S1, a B-cell fraction >10% was a strong predictor of increased relative risks and shorter mean time to diagnosis, implying this group may have included subjects with undiagnosed MBL or CLL at recruitment. We also noted that the DNA methylation and transcriptomic profiles (adjusted for WBC composition) of control subjects with B-cell fraction >10% differed significantly from those of subjects with <10% and could not exclude the possibility that they may have been carriers, at the time of recruitment, of small clones of altered cells related to CLL (see Additional file 1: Text). Based on these observations, we opted to exclude control subjects with >10% B-cells from the derivation of all differential profiles of CLL cases discussed below (unless otherwise indicated) so as to ensure that the derived profiles reflect to the maximum degree early and mechanistically informative changes. A flowchart of the comparisons conducted using different subgroups of subjects, some of which are presented in detail in Supplementary, is shown in Fig. 1, while the numbers of differential signals obtained in the various comparisons are summarized in Additional file 2: Table S2.

Flowchart of data analyses
DNA methylation profile
Comparison of the DNA methylation profiles of the CLL cases and the controls, with adjustment for WBC composition included in the statistical model, resulted in the identification of 722 differentially methylated (DM) CpG sites significant at Bonferroni-corrected p < 0.05 (corresponding to 494 unique genes), of which 534 showed loss of methylation in cases (mean loss 4.9%, range 0.9–30.4%), while the remaining showed methylation gain (mean gain 1.8%, range 0.1–7.9%) (Table 2 and Additional file 2: Table S3; for a discussion of the corresponding analysis without adjustment for WBC composition, as well as an assessment of the profile robustness across the two cohorts, see Additional file 1: Text). Of the 722 DM CpG sites, 530 (73.4%) overlap with 33,653 sites reported to distinguish CLL from normal B-cells [24] and show the same direction of methylation change, indicating that the majority of the DNA methylation changes which characterize our pre-diagnostic CLL risk profile are among those which accompany the transformation of normal B-cells to overt CLL clones.
To explore the relationship of the CLL risk-related DNA methylation changes with B-cells, from the 10,785 FDR-significant DM CpGs of the CLL risk-related profile we selected those sites (1318) known to show minimal variation between different WBC subtypes [22] and performed principal component analysis (PCA) using the corresponding signal levels after denoising for batch effects, gender, cohort and smoking status (see Statistical Analysis in Additional file 1: Text). As can be seen in Fig. 2, the 28 cases are separated not only from the controls but also from each other according to their B-cell fraction. In contrast, use of an equal number of CpG sites, selected randomly from among those showing minimal variation between WBC subtypes but not included in our CLL risk-related profile, failed to yield analogous distributions. This indicates that the CLL risk-related DNA methylation signals arise in cells which carry the epigenetic hallmarks of B-cells and probably represent CLL-related B-cells.

Top: PCA based on 1308 CpG sites significant (FDR < 0.05) in CLL cases and with minimal variation between WBC sub-populations. Bottom: similar analysis with 1308 CpG sites randomly selected from among those with FDR > 0.8 in CLL cases and with minimal variation between WBC sub-populations. The signal intensities employed were denoised for various parameters, including B-cell content (see Methods). The numbers in the Figures on the right indicate the fractional B-cell content of the samples. The Figures on the left show all subjects while those on the right show only the CLL case subjects
A number of loci appear to serve as targets for extensive epigenetic modification. Thus, 176 genes and 50 intragenic CpG islands (CGIs) were represented by at least 3 (and up to 16) DM CpG sites (FDR < 0.05) each and had an enrichment (fraction of DM sites among the locus-associated sites analysed on the microarray) of at least 20% (and up to 66.7%). Furthermore, 8 of these 50 CGIs are located just upstream of the TSS1500 region of some of the same 176 genes (Additional file 2: Table S4). Gene functional classification analysis (see Bioinformatic Analysis in Additional file 1: Text) showed that the most abundant functional group among the 176 multiply DM genes consisted of 38 homeobox or homeobox-related genes, including those present in the HOX and IRX gene clusters.
Transcriptomic profile
Comparison of the transcriptomic profiles of 25 of our CLL cases, for which such profiles were also available, with those of control subjects (with <10% B-cells), yielded 16 differentially expressed (DE) probes, corresponding to 15 unique genes (Bonferroni-corrected p < 0.05; 117 probes/82 unique genes at FDR < 0.05) (Table 3 and Additional file 2: Table S5). Eleven of the Bonferroni- and 34 of the FDR-significant genes have been reported [32, 33] to be differentially expressed in B-cells isolated from CLL patients. Furthermore, 22 FDR-significant DE genes are among those found in CLL, with the same direction of change (Additional file 2: Table S5), in a meta-analysis of transcriptomic profiles of CLL patients [18]. These observations further support the suggestion that cells with changes characteristic of clinical CLL were present in our prediagnostic samples. As discussed in Supplementary, omission of adjustment for WBC composition results in a greatly increased number of significant signals, reflecting the influence of varying cell sub-populations. Nevertheless, the top 6 DE genes shown in Table 3 are the same as the top signals obtained without WBC correction (also previously reported by Chadeau-Hyam et al. [17]) and show the same direction of change. On the other hand, while the results obtained without WBC correction showed overexpression in cases of the vast majority of the significant signals, among the 117 DE signals obtained with WBC correction 51 were under-expressed in cases.
miRNA profile
For 11 CLL cases and 96 controls from the NHSDS cohort we were also able to examine the miRNA expression profile. We observed 2 significant signals (FDR < 0.05), miR-155-5p and miR-150-5p, both overexpressed (2.3- and 2.2fold, respectively) in cases. miR155-5p was also observed among the overexpressed genes in the transcriptomic profile, and it is notable that this gene is undermethylated in 2 out of the 6 related CpG sites in the DNA methylation profile. Both miRNAs have been reported to be overexpressed in B cells from individuals with MBL, and even more so in patients with CLL [34].
Taken together, the above results indicate that distinct epigenetic and gene expression changes, most of which are known to be associated with clinically diagnosed CLL, were present in our prediagnostic samples. Significantly, as discussed in Additional file 1: Text, we detected some of these signals also in the sub-group of cases with B-cell fraction <10% (which are less likely to have included subjects with undiagnosed CLL at recruitment) while the mean intensities of the differential signals (after adjustment for the number of B-cells) in this group were smaller than those observed in the group which includes all cases, suggesting a process of signal evolution as the size of the CLL-like cell clones increased.
CLL risk profiles in CLL cases with <10% B-cells
In view of the evidence, discussed above, of presence in blood samples of CLL cases of clones of cells related to CLL or precursor conditions, we checked whether this was also true for cases with B-cell fraction < 10%, who are less likely to have been suffering from undiagnosed MBL or CLL at recruitment. As indicated in Additional file 2: Table S2, this subgroup (11 cases) could also be differentiated in terms of CpG methylation from the controls, although with a dramatically reduced number of signals (4 and 45 CpG sites significant at p < 0.05 after Bonferroni or FDR correction, respectively). All but one of these DM CpGs (Additional file 2: Table S6) are among FDR-significant CpGs observed when all cases were taken into consideration, while 25 are among those reported to distinguish CLL from normal B-cells [24]. Turning to the transcriptomic profile (Additional file 2: Table S6, lower part), 12 DE probes significant at FDR < 0.05 (4 significant at Bonferroni-corrected p < 0.05) were observed in low-B-cell count cases, of which 7 were also found among signals (FDR < 0.05) observed when all cases were considered, while 3 have been reported to distinguish CLL from normal B-cells [32]. No differences significant at FDR < 0.05 were observed in the miRNA profiles.
Comparison of the p-rankings of DM and DE signals observed in all cases and in those with B-cells < 10% (not shown), as well as the denoised case-control methylation differences (Δβ) or fold-change expression ratios showed strong correlations, while the mean intensities of the differences from the controls, after adjusting for B-cell content, observed in low-B-cell cases are smaller than those in all cases (Fig. 3). These observations suggest that cases with B-cell fraction <10% contained clones of cells related to CLL which evolved further towards the CLL phenotype as the CLL-like cell clones grew larger.

Comparison of denoised case-control methylation differences (Δβ) (left) and expression differences (foldchange ratio) (right), obtained from the comparison of all cases with controls with <10% B-cells (vertical axes) or with all controls (horizontal axes). The light lines show slope = 1
Τime-To-diagnosis and CLL risk-related profiles
In view of the large variation in the time between the donation of the biological samples and clinical diagnosis of CLL (2.0–15.9 years), we derived risk-related profiles separately for subjects with time to diagnosis (TtD) longer or shorter than the median of 7.4 years. It is noted that diagnosis, as registered in the cohorts of the present study, may have corresponded to disease at different stages of progression for different case subjects, taking place in some cases accidentally during routine examination in some cases, after the appearance of clinical symptoms but not requiring treatment in others or coinciding with treatment in other subjects.
We observed 238 and 937 DM CpGs (Bonferroni-corrected p < 0.05) in the long and short TtD subgroups, respectively, with most (181) of the former signals being at least FDR-significant in the latter (Additional file 2: Table S7). Similarly, there were 25 and 291 DE probes (FDR p < 0.05) in the long and short TtD subgroups, respectively, with an overlap of 8 signals. Of the 238 DM CpGs observed in the long TtD sub-group, 168 (70.5%) have been reported to be differentially modified in clinically diagnosed CLL [24]. Furthermore, of the 21 DE genes significant in the same sub-group, 12 are among 2095 genes (hypergeometric distribution test p = 0.013) reported to be differentially expressed in clinical CLL [32, 33] and 9 are among those found significant in a corresponding meta-analysis included in the report by Vlaanderen et al. [18]. Finally turning to miRNA, examination of the limited number of subjects for which data were available indicates that, while no significant changes in miRNA expression could be detected in the long TtD group, 3 miRNAs (miR-155-5p, miR-150-5p, both overexpressed, and miR-4486, underexpressed) were significant (FDR < 0.05) in the short TtD group.
The above observations indicate that epigenetic and gene expression changes characteristic of CLL are already present in subjects 7.4–15.7 years prior to clinical diagnosis of the disease. On the other hand, comparison of the intensities of CLL risk-related signals in control subjects and the two sub-groups with different TtD suggests the occurrence of progressive changes while approaching clinical manifestation and diagnosis of the disease. As shown in Fig. 4 (top panels), during this time the least squares means (LSM, adjusted for B-cell content – see Statistical Analysis in Supplementary) of the methylation levels of the top DM CpG sites obtained with all cases (Table 2) change in a consistent manner (mainly loss of methylation for the top 20 sites) and independently of the size of the B-cell clones. Additionally, a substantial increase in the multiplicity of gene methylation is observed in the shorter TtD group, with the average multiplicities of DM genes with at least 20% enrichment in the short TtD subgroup being 1.9 and 5.4 DM sites per gene in the long and short TtD groups, respectively (Additional file 2: Table S8). As regards the corresponding evolution of the gene expression signals (Fig. 4, bottom panels), while for most of the 16 Bonferroni-significant CLL risk-related DE signals observed in all cases expression increases in the long TtD subgroup and then remains relatively constant during the later period coming up to diagnosis, changes in both directions appear to occur, at lower statistical significance, in a large number of genes.

Normalized least squares means (LSM) of DM and DE signal values in the controls and the two TtD groups. Top left: 238 DM signals Bonferroni-significant in the short TtD subgroup; bottom left: 291 DE signals FDR-significant in the short TtD subgroup. Top right: top 20 DM signals observed in all cases; bottom right: Bonferroni -significant DE signals observed in all cases. In the Figures on the right are named the genes associated with the 3 DM or DE signals with the largest changes in each direction (only 2 DM genes show increased methylation with decreasing TtD)
Functional analysis
Gene enrichment analyses were conducted for 494 DM genes associated with DM CpGs (Bonferroni-corrected p < 0.05) in the CLL risk-related profile, using the recently developed BioInfoMiner web application [29] which is appropriate for the functional analysis of DNA methylation profiles (for more details see Bioinformatics analysis in Methods). For the corresponding analysis of the transcriptomic changes, in view of the small number of signals significant at Bonferoni-adjusted p < 0.05 (16 transcripts, 15 genes; see Table 3) which does not permit the conduct of functional analysis, we relaxed the significance criterion to FDR < 0.1. In combination with the additional statistical filters applied at the functional analysis level, such relaxation retains a strong overall statistical stringency and yields 163 DE genes. The over-represented terms thus obtained are shown in Additional file 2: Tables S9-S10. We conducted similar analyses also with the CLL risk-related gene profiles of cases with long and short TtD (Additional file 2: Tables S11-S12; no DE gene analysis was conducted for the long TtD subgroup owing to the small number of genes involved). The most notable observation is the predominance among GO terms derived from DM genes, for all sub-groups, of terms related to development and regulation of transcription as well as B-cell differentiation and physiology. Also of note is the presence of multiple DE gene-derived terms (including pathways) related to DNA damage response and WNT signaling.
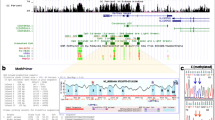
In order to identify genes which play a central role in the biological processes leading to CLL, we used the gene prioritization module of the BioInfoMiner application to identify hub genes which are linked to multiple ontology terms, thus identifying 84 DM and 18 DE such genes (Additional file 2: Table S13). DM hubs include numerous genes that encode for transcription factors, especially members of the homeobox family (PAX6, multiple HOX genes, FOXP1, EN2, GSC, EVX1, BARHL2). DE hub genes include, among others, 4 genes related to WNT signaling (WNT3, CTBP1, CTNNB1 and TCF4), while an additional WNT pathway gene, CTBP2, is among the DM hub genes. Examination, using the online resource Search Tool for the Retrieval of Interacting Genes (STRING) [35], of the protein-protein interaction network of the combined DM and DE hubs reveals the existence of 2 major nodes, centered on the important epigenetic modification gene HDAC1 and the WNT signaling pathway gene CTNNB1 (Fig. 5). The list of DM hub genes in the profile of long and short TtD sub-groups was also dominated by homeobox genes, while the corresponding DE genes included two genes related to WNT signaling, WNT3 for the long TtD group and CTNNB2 for the short TtD group (Additional file 2: Table S14).

Interaction network of the DM + DE hubs (STRING)
ROC analysis and development of a DNA methylation-based classification signature for predicting the development of clinical CLL
We recently reported on the advantages of using the semantics information included in the hierarchical nature of ontologies as a primary feature selection tool for the development of predictive profiles [36]. Based on this we assessed the ability of prediagnostic DM CpG sites to predict the future clinical manifestation of CLL among all subjects (i.e. without excluding case or control subjects with >10% B-cells) by focusing on 104 Bonferroni-significant DM CpG sites annotated to the hub genes. The reason for using here the profile obtained without exclusion of any of the controls is to facilitate the derivation of a predictive signature which can be of use in the general population. We employed as a training set a balanced subset corresponding to 50% of the study subjects and assessed the performance of a number of different classifiers using the remaining 50% of the population as a testing set (see ROC Analysis in Additional file 1: Text). An AUC value of 0.94 was obtained when the SVM linear model was used as a classifier. Subsequently, a recursive feature elimination algorithm, with maximum number of predictors set to 40, was used to identify an optimal subset of predictors, achieving an optimal accuracy of 95% using the Naïve Bayesian classifier and 12 predictors.
Discussion
B-cells with CLL-like DNA methylation and gene expression features are present in the blood more than 10 years before disease diagnosis
Using genome-wide gene expression and DNA methylation profiles observed in prediagnostic samples of whole blood leukocytes of subjects who were diagnosed with CLL 2–15.9 years later, we have derived WBC composition-corrected differential profiles which are associated with the risk of future diagnosis with CLL. Although the numbers of differential signals detected after adjustment for WBC composition are dramatically decreased relative to those found without such adjustment (Additional file 2: Table S2) [17], the majority of these signals (approx. 70% for both methylation and gene expression, both miRNAs), are known to be similarly modified in clinical CLL [18, 24, 32,33,34, 37]. The same holds for the signals detected in the sub-groups of cases with B-cell fraction <10% (Additional file 2: Table S6) and TtD > 7.4 years (Additional file 2: Table S7), supporting the idea that the differential risk profiles identified are unlikely to represent false findings resulting from residual confounding by variations in WBC composition but are in fact associated with the early phase of the pathogenesis of CLL. Further support for this comes from the fact that mutations in 12 of the DM genes (FARP2, ACTA2, AcOXL, BCL2, BMF, CLPTM1L, CPEB1, CSRNP1, IPCEF1, LPP, ODF1, SERPINB6, mostly overmethylated in cases, some at multiple CpG sites), as well as in 1 DE gene (C11orf21, overexpressed in cases at FDR = 0.086, have been found in GWAS studies to be associated with differential risk of CLL [38,39,40,41].
It is notable that the lists of DE and DM genes lost upon WBC adjustment overlap significantly (not shown), suggesting that their detection without WBC correction reflects variation in cell composition. Although the large impact of such correction in the present case is likely to be due to the large increase in the fraction of B-cells in CLL cases, it does underline the importance of variation in WBC composition being considered as a potential confounder not only in methylation but also in expression profiling studies of mixed cell populations [42].
The top genes in the DM and DE profiles include multiple genes related to the transport of ions and other small molecules (SEC14L1, SLC43A2, ATP9B, ABC6 and MCOLN2), as well as genes with GTPase-related activity (RASA3 and TBCD), however the role of these genes in CLL pathogenesis is not known. On the other hand, the top DE genes include 4 genes involved in WNT signaling - WNT3, TCF4, CDK14, all upregulated in cases, and CTNNB1, downregulated in cases (plus a fifth gene, CTBP1, also upregulated in cases with a statistical significance of FDR = 0.086), a key pathway in CLL (31). The DE profile also includes 2 important B-cell associated genes, CD24, upregulated in cases, which regulates B-cell growth, differentiation and activation and IGHD, downregulated in cases, the main antigen receptor on B-cells, involved in BCR signaling and the CLL-growth/survival pathway [43].
Using the significance cut-off values of Bonferroni-corrected p < 0.05 for DM and FDR < 0.05 for DE signals, only 1 gene (FAM193A, of unknown relevance to CLL) was found to be differentially both methylated and expressed in the profile obtained with WBC adjustment, supporting previous suggestions that DNA methylation is not a primary mechanism of gene regulation in CLL [24, 44].
We found two miRNAs, miR-150-5p and miR-155-5p to be consistently overexpressed in CLL cases, including the subgroups with long and short TtD. The latter gene was also differentially undermethylated at 2 CpG sites in cases. Both of these miRNAs are overexpressed in clinical CLL, the first being the most abundant miRNA in B-cells from CLL patients [34, 37]. A search on the Targetscan and miRDB databases reveals 3 of the genes which are differentially downregulated in cases to be among the targets of miR-150-5p, specifically TLDC1 and GLDC, of unknown function in relation to CLL, as well as the CTNNB1 gene which is related to WNT signaling, underlining the importance of this pathway in early stages of CLL pathogenesis.
To obtain a more global picture of the cellular perturbations represented by the CLL risk-related profiles, we searched for genes which are linked with multiple biological functions (hub genes) (Additional file 2: Table S13). Among the DM hub genes we identified numerous homeobox genes, including PAX6, multiple HOX genes, FOXP1, EN2, GSC, EVX1 and BARHL2, many of which (especially HOXA and HOXD) are targets for multiple differential methylation events. HOX genes are involved in normal and leukemic hematopoiesis and may contribute to the mechanism of leukemic transformation [45,46,47], while FOXP1 plays a particularly important role in B-cell development and enhances B-cell signaling in CLL cells [48, 49]. DE hubs include, among others, the CBX6 gene, upregulated in cases, which is part of a polycomb complex required to maintain the transcriptionally repressed state of many genes (including HOX) during development [50]. Other DE hub genes include CTBP1, a transcriptional repressor and a key downstream component of the WNT signaling pathway and NOS2, upregulated in cases, which regulates the levels of NO which enhances the synthesis of pro-inflammatory mediators [51, 52]. The interaction network of the DM and DE hubs involves two major nodes centered upon HDAC1 and CTNNB1, with a direct chain of interactions linking HDAC1, CTNNB1, SKI, CTBP1 and FOXP1 (Fig. 5). HDAC1 is overexpressed in CLL [53] and has been shown to be instrumental in the induction of early epigenetic alterations and subsequent gene expression changes in a mouse model of CLL [54], while the SKI gene, in addition to its oncogenic and oncosuppressor functions, has been suggested to contribute to memory B cell differentiation [55].
The picture emerging from the above observations suggests the presence in our prediagnostic samples of epigenetic and expression perturbations in genes with a key role in B-cell development and differentiation. This is further supported by the results of our functional analysis, which identified GO terms which correspond mostly to cell proliferation, differentiation, developmental and regulatory processes. Of particular note is the identification of multiple ontology terms relate to B-cell physiology and morphology, as well as abnormalities of immune system physiology, such as immunodeficiency and agammaglobulinemia (Additional file 2: Table S10 and Additional file 3 Figure S1), which are characteristic clinical findings in CLL [56]. Finally, pathway analysis revealed changes in lipid and lipoprotein metabolism, Fc gamma and epsilon receptor signaling, as well as NGF, VEGF, WNT, NOTCH and B-cell signaling and DNA damage response pathways (Additional file 2: Tables S9 and S10), all of which are known to be perturbed CLL [57,58,59,60,61,62].
CLL-related risk profiles in prediagnostic samples evolve as the B-cell clones grow and time to diagnosis decreases
The intensity of many of the early risk-related signals discussed above (especially DNA methylation-related signals) was lower in case subjects with B-cell fraction <10%, relative to that observed in all case subjects, even after adjusting for cell composition (Fig. 3). Furthermore, and while it is recognised that the use of time-to-diagnosis, rather than, e.g., time-to-first-treatment, has the limitation that diagnosis may relate to disease at varying stages of advancement, the intensity of such signals varied in a consistent manner as the time to clinical diagnosis of the disease became shorter (Fig. 4), implying that the perturbations of the cell clones in which these signals were located evolved as disease pathogenesis progressed while still remaining in a subclinical phase. Among the top risk-related epigenetic signals, DNA methylation showed mainly a pattern of decreasing levels throughout the observation period, while the top gene expression signals showed initially mainly upregulation followed by stabilization, however large numbers of signals of both types showed a consistent trend towards either over- or underexpression as the time to diagnosis became shorter (Fig. 4). Of particular interest is the evolution of WNT3 expression, a key gene in CLL, which appeared to be upregulated in cases with long TtD and subsequently remain unchanged as TtD decreases, implying that an increase in WNT signaling may be an early change in CLL.
The picture of an evolving preclinical profile is also evident in the increasing multiplicity of DM CpG sites at specific loci (Additional file 2: Table S11), including 2 members of the HOX and 3 members of the IRX families which are located as clusters at different chromosomal regions. Additional file 3: Figure S2 illustrates that the tendency to accumulate increased levels of methylation at shorter TtD holds for all clusters of these genes and affects additional genes associated with these loci, such as miR196b and the HOX paralog genes EVX1 and EVX2, as well as associated CGIs.
Focusing on DM gene hubs which are associated with long TtD (Additional file 2: Table S14) and therefore may play a role particularly in the early stages of CLL pathogenesis, the presence is noted of multiple developmentally important homeobox genes and transcription factors, implying developmental deregulation as an important part of early perturbations during CLL pathogenesis, as also indicated by the GO terms identified from the corresponding functional analysis (Additional file 2: Table S11). These genes include, among others, CSK which plays an important role in the regulation of cell growth, differentiation, migration and immune response and suppresses signaling by BCR [63], and CTBP1, involved in WNT and NOTCH signaling [64].
A DNA methylation signature predictive of future CLL risk
We tested the ability of the set of 104 DM CpG sites associated with CLL risk-related hub genes (i.e. genes with major biological roles) to classify subjects with regard to their future risk of being diagnosed with CLL, using for this purpose a variety of classification algorithms (see ROC Analysis in Methods). An excellent predictive ability (AUC = 0.94) was obtained using all 104 of the above signals with a Naive Bayes classifier, and a similar value (AUC = 0.95) was found when just 12 predictors were selected from the above set. A high predictive ability was also observed when we used as the test set all the control samples mixed with CLL cases with <10% WBC (AUC = 0.86).
Conclusions
We have shown that changes in CpG methylation and gene (including miRNA) expression similar to those observed in clinical CLL are present in B-cells of apparently healthy subjects 2.1–15.9 years before the diagnosis of CLL and have provided evidence supporting the idea that such changes evolve as the disease progresses from its pre-clinical stage towards clinical diagnosis. We have also shown that a CpG methylation-based signature can predict future CLL diagnosis with great accuracy. Relative to other predictive markers also reported, this predictive signature has the advantage of increased biological plausibility since it is based on hub genes many of which are known to be involved in the pathogenesis of CLL. Although patients diagnosed with cMBL or early-stage CLL do not required therapy, the great progress being made in the characterization of the genetic and epigenetic landscapes of these conditions [44, 65] raises the prospect of the development of interventions which may slow down or block progression towards full disease. In this context, early disease biomarkers that define the patients that are likely to progress to symptomatic CLL and could provide data for an individualized surveillance program, with higher intensity for some patients. Also if strong negative predictors could be defined, this could identify patients that could be reassured and not in need for follow up.
Abbreviations
- CGI:
-
CpG island
- CLL:
-
Chronic lymphocytic leukemia
- DE:
-
Differentially expressed
- DM:
-
Differentially methylated
- LSM:
-
Least squares means
- MBL:
-
Monoclonal B-cell lymphocytosis
- NSHDS:
-
Northern Sweden Health and Disease Study
- TtD:
-
Time-to-diagnosis
- WBC:
-
White blood cells
References
Kipps TJ, Stevenson FK, Wu CJ, Croce CM, Packham G, Wierda WG, et al. Chronic lymphocytic leukaemia. Nat Rev Dis Primer. 2017;3:16096.
Rodríguez-Vicente AE, Díaz MG, Hernández-Rivas JM. Chronic lymphocytic leukemia: a clinical and molecular heterogenous disease. Cancer Genet. 2013;206:49–62.
Hallek M. Chronic lymphocytic leukemia: 2015 update on diagnosis, risk stratification, and treatment. Am J Hematol. 2015;90:446–60.
Landgren O, Albitar M, Ma W, Abbasi F, Hayes RB, Ghia P, et al. B-cell clones as early markers for chronic lymphocytic leukemia. N Engl J Med. 2009;360:659–67.
Shim YK, Middleton DC, Caporaso NE, Rachel JM, Landgren O, Abbasi F, et al. Prevalence of monoclonal B-cell lymphocytosis: a systematic review. Cytometry B Clin Cytom. 2010;78(Suppl 1):S10–8.
Morabito F, Mosca L, Cutrona G, Agnelli L, Tuana G, Ferracin M, et al. Clinical monoclonal B lymphocytosis versus Rai 0 chronic lymphocytic leukemia: a comparison of cellular, cytogenetic, molecular, and clinical features. Clin Cancer Res Off J Am Assoc Cancer Res. 2013;19:5890–900.
Rawstron AC, Bennett FL, O’Connor SJM, Kwok M, Fenton JAL, Plummer M, et al. Monoclonal B-cell lymphocytosis and chronic lymphocytic leukemia. N Engl J Med. 2008;359:575–83.
Barrio S, Shanafelt TD, Ojha J, Chaffee KG, Secreto C, Kortüm KM, et al. Genomic characterization of high-count MBL cases indicates that early detection of driver mutations and subclonal expansion are predictors of adverse clinical outcome. Leukemia. 2016;31:170–6.
Henriques A, Rodríguez-Caballero A, Nieto WG, Langerak AW, Criado I, Lécrevisse Q, et al. Combined patterns of IGHV repertoire and cytogenetic/molecular alterations in monoclonal B lymphocytosis versus chronic lymphocytic leukemia. PLoS One. 2013;8:e67751.
Rossi D, Sozzi E, Puma A, De Paoli L, Rasi S, Spina V, et al. The prognosis of clinical monoclonal B cell lymphocytosis differs from prognosis of Rai 0 chronic lymphocytic leukaemia and is recapitulated by biological risk factors. Br J Haematol. 2009;146:64–75.
Shanafelt TD, Kay NE, Rabe KG, Call TG, Zent CS, Schwager SM, et al. Survival of patients with clinically identified monoclonal B-cell lymphocytosis (MBL) relative to the age- and sex-matched general population. Leukemia. 2012;26:373–6.
Hosnijeh FS, Portengen L, Späth F, Bergdahl IA, Melin B, Mattiello A, et al. Soluble B-cell activation marker of sCD27 and sCD30 and future risk of B-cell lymphomas: a nested case-control study and meta-analyses. Int J Cancer. 2016;138:2357–67.
Kaaks R, Sookthai D, Łuczyńska A, Oakes CC, Becker S, Johnson T, et al. Lag times between lymphoproliferative disorder and clinical diagnosis of chronic lymphocytic leukemia: a prospective analysis using plasma soluble CD23. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2015;24:538–45.
Kim C, Bassig BA, Seow WJ, Hu W, Purdue MP, Huang W-Y, et al. Mitochondrial DNA copy number and chronic lymphocytic leukemia/small lymphocytic lymphoma risk in two prospective studies. Cancer Epidemiol. Biomark Prev Publ Am Assoc Cancer Res Cosponsored Am Soc Prev Oncol. 2015;24:148–53.
Nieters A, Łuczyńska A, Becker S, Becker N, Vermeulen R, Overvad K, et al. Prediagnostic immunoglobulin E levels and risk of chronic lymphocytic leukemia, other lymphomas and multiple myeloma-results of the European prospective investigation into cancer and nutrition. Carcinogenesis. 2014;35:2716–22.
Späth F, Wibom C, Krop EJM, Johansson A-S, Bergdahl IA, Vermeulen R, et al. Biomarker dynamics in B-cell lymphoma: a longitudinal prospective study of plasma samples up to 25 years before diagnosis. Cancer Res. 2017;77:1408–15.
Chadeau-Hyam M, Vermeulen RCH. Hebels DG a. J, Castagné R, Campanella G, Portengen L, et al. Prediagnostic transcriptomic markers of chronic lymphocytic leukemia reveal perturbations 10 years before diagnosis. Ann. Oncol. Off. J. Eur. Soc. med. Oncologia. 2014;25:1065–72.
Vlaanderen J, Leenders M, Chadeau-Hyam M, Portengen L, Kyrtopoulos SA, Bergdahl IA, et al. Exploring the nature of prediagnostic blood transcriptome markers of chronic lymphocytic leukemia by assessing their overlap with the transcriptome at the clinical stage. BMC Genomics. 2017;18:239.
Genomics Biomarkers of Environmental Health. http://www.envirogenomarkers.net. Accessed 25 May 2017.
Hebels DGAJ, Georgiadis P, Keun HC, Athersuch TJ, Vineis P, Vermeulen R, et al. Performance in omics analyses of blood samples in long-term storage: opportunities for the exploitation of existing biobanks in environmental health research. Environ Health Perspect. 2013;121:480–7.
Houseman EA, Accomando WP, Koestler DC, Christensen BC, Marsit CJ, Nelson HH, et al. DNA methylation arrays as surrogate measures of cell mixture distribution. BMC Bioinformatics. 2012;13:86.
Jaffe AE, Irizarry RA. Accounting for cellular heterogeneity is critical in epigenome-wide association studies. Genome Biol. 2014;15:R31.
Smith EN, Ghia EM, DeBoever CM, Rassenti LZ, Jepsen K, Yoon K-A, et al. Genetic and epigenetic profiling of CLL disease progression reveals limited somatic evolution and suggests a relationship to memory-cell development. Blood Cancer J. 2015;5:e303.
Kulis M, Heath S, Bibikova M, Queirós AC, Navarro A, Clot G, et al. Epigenomic analysis detects widespread gene-body DNA hypomethylation in chronic lymphocytic leukemia. Nat Genet. 2012;44:1236–42.
Coonen ML, Theunissen DH, Kleinjans JC, Jennen DG. MagiCMicroRna: a web implementation of AgiMicroRna using shiny. Source Code Biol Med. 2015;10:4.
Valavanis I, Sifakis EG, Georgiadis P, Kyrtopoulos S, Chatziioannou AA. A composite framework for the statistical analysis of epidemiological DNA methylation data with the Infinium human Methylation 450K BeadChip. IEEE J Biomed Health Inform. 2014;18:817–23.
Lenth RV. Using lsmeans [Internet]. 2016 [cited 2016 Dec 22] Available from: https://cran.r-project.org/web/packages/lsmeans/vignettes/using-lsmeans.pdf
DAVID Functional Annotation Bioinformatics Microarray Analysis. https://david.ncifcrf.gov/home.jsp. Accessed 25 May 2017.
Koutsandreas T, Binenbaum I, Pilalis E, Valavanis I, Papadodima O, Chatziioannou A. Analyzing and visualizing genomic complexity for the derivation of the emergent molecular networks. Int J Monit Surveill Technol Res IJMSTR. 2016;4(2):30–49.
Moutselos K, Maglogiannis I, Chatziioannou A. GOrevenge: a novel generic reverse engineering method for the identification of critical molecular players, through the use of ontologies. IEEE Trans Biomed Eng. 2011;58:3522–7.
Lunardon N, Menardi GG, Torelli NN. ROSE: a package for binary imbalanced learning. R J. 2014;6:79–89.
Ferreira PG, Jares P, Rico D, Gómez-López G, Martínez-Trillos A, Villamor N, et al. Transcriptome characterization by RNA sequencing identifies a major molecular and clinical subdivision in chronic lymphocytic leukemia. Genome Res. 2014;24:212–26.
Liao W, Jordaan G, Nham P, Phan RT, Pelegrini M, Sharma S. Gene expression and splicing alterations analyzed by high throughput RNA sequencing of chronic lymphocytic leukemia specimens. BMC Cancer. 2015;15:714.
Ferrajoli A, Shanafelt TD, Ivan C, Shimizu M, Rabe KG, Nouraee N, et al. Prognostic value of miR-155 in individuals with monoclonal B-cell lymphocytosis and patients with B chronic lymphocytic leukemia. Blood. 2013;122:1891–9.
Franceschini A, Szklarczyk D, Frankild S, Kuhn M, Simonovic M, Roth A, et al. STRING v9.1: Protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013;41:D808–15.
Valavanis I, Pilalis E, Georgiadis P, Kyrtopoulos S, Chatziioannou A. Cancer biomarkers from genome-scale DNA Methylation: comparison of evolutionary and semantic analysis methods. Microarrays Basel Switz. 2015;4:647–70.
Mraz M, Chen L, Rassenti LZ, Ghia EM, Li H, Jepsen K, et al. miR-150 influences B-cell receptor signaling in chronic lymphocytic leukemia by regulating expression of GAB1 and FOXP1. Blood. 2014;124:84–95.
Berndt SI, Camp NJ, Skibola CF, Vijai J, Wang Z, Gu J, et al. Nat Commun. 2016;7:10933.
Berndt SI, Skibola CF, Joseph V, Camp NJ, Nieters A, Wang Z, et al. Genome-wide association study identifies multiple risk loci for chronic lymphocytic leukemia. Nat Genet. 2013;45:868–76.
Speedy HE, Di Bernardo MC, Sava GP, Dyer MJS, Holroyd A, Wang Y, et al. A genome-wide association study identifies multiple susceptibility loci for chronic lymphocytic leukemia. Nat Genet. 2014;46:56–60.
Crowther-Swanepoel D, Broderick P, Di Bernardo MC, Dobbins SE, Torres M, Mansouri M, et al. Common variants at 2q37.3, 8q24.21, 15q21.3 And 16q24.1 Influence chronic lymphocytic leukemia risk. Nat. Genet. 2010;42:132–6.
Trapnell C. Defining cell types and states with single-cell genomics. Genome Res. 2015;25:1491–8.
Hobeika E, Maity PC, Jumaa H. Control of B cell responsiveness by Isotype and structural elements of the antigen receptor. Trends Immunol. 2016;37:310–20.
Oakes CC, Seifert M, Assenov Y, Gu L, Przekopowitz M, Ruppert AS, et al. DNA methylation dynamics during B cell maturation underlie a continuum of disease phenotypes in chronic lymphocytic leukemia. Nat Genet. 2016;48:253–64.
Fournier M, Lebert-Ghali C-É, Krosl G, Bijl JJ. HOXA4 Induces expansion of hematopoietic stem cells in vitro and confers enhancement of pro-B-cells in vivo. Stem Cells Dev. 2012;21:133–42.
Owens BM, Hawley RG. HOX and non-HOX homeobox genes in leukemic hematopoiesis. Stem Cells Dayt Ohio. 2002;20:364–79.
Taghon T, Stolz F, De Smedt M, Cnockaert M, Verhasselt B, Plum J, et al. HOX-A10 regulates hematopoietic lineage commitment: evidence for a monocyte-specific transcription factor. Blood. 2002;99:1197–204.
Hu H, Wang B, Borde M, Nardone J, Maika S, Allred L, et al. Foxp1 Is an essential transcriptional regulator of B cell development. Nat Immunol. 2006;7:819–26.
Sagardoy A, Martinez-Ferrandis JI, Roa S, Bunting KL, Aznar MA, Elemento O, et al. Downregulation of FOXP1 is required during germinal center B-cell function. Blood. 2013;121:4311–20.
Golbabapour S, Majid NA, Hassandarvish P, Hajrezaie M, Abdulla MA, Hadi AHA. Gene silencing and Polycomb group proteins: an overview of their structure, mechanisms and Phylogenetics. OMICS J Integr Biol. 2013;17:283–96.
Korhonen R, Lahti A, Kankaanranta H, Moilanen E. Nitric oxide production and signaling in inflammation. Curr Drug Targets Inflamm Allergy. 2005;4:471–9.
Thomas DD, Heinecke JL, Ridnour LA, Cheng RY, Kesarwala AH, Switzer CH, et al. Signaling and stress: the redox landscape in NOS2 biology. Free Radic Biol Med. 2015;87:204–25.
Wang JC, Kafeel MI, Avezbakiyev B, Chen C, Sun Y, Rathnasabapathy C, et al. Histone deacetylase in chronic lymphocytic leukemia. Oncology. 2011;81:325–9.
Chen S-S, Raval A, Johnson AJ, Hertlein E, Liu T-H, Jin VX, et al. Epigenetic changes during disease progression in a murine model of human chronic lymphocytic leukemia. Proc Natl Acad Sci U S A. 2009;106:13433–8.
Bhattacharya D, Cheah MT, Franco CB, Hosen N, Pin CL, Sha WC, et al. Transcriptional profiling of antigen-dependent murine B cell differentiation and memory formation. J Immunol. 2007;179:6808–19.
Forconi F, Moss P. Perturbation of the normal immune system in patients with CLL. Blood. 2015;126:573–81.
Aguirre Palma LM, Gehrke I, Kreuzer K-A. Angiogenic factors in chronic lymphocytic leukaemia (CLL): where do we stand? Crit Rev Oncol Hematol. 2015;93:225–36.
Filip AA. New boys in town: prognostic role of SF3B1, NOTCH1 and other cryptic alterations in chronic lymphocytic leukemia and how it works. Leuk Lymphoma. 2013;54:1876–81.
Fridman WH, Mathiot C, Montcuit J, Teillaud JL. Fc receptors, immunoglobulin-binding factors and B chronic lymphocytic leukemia. Nouv Rev Fr Hematol. 1988;30:311–5.
Gandhirajan RK, Poll-Wolbeck SJ, Gehrke I, Kreuzer K-A. Wnt/β-catenin/LEF-1 signaling in chronic lymphocytic leukemia (CLL): a target for current and potential therapeutic options. Curr Cancer Drug Targets. 2010;10:716–27.
Molica S, Vitelli G, Levato D, Crispino G, Dell’Olio M, Dattilo A, et al. CD27 In B-cell chronic lymphocytic leukemia. Cellular expression, serum release and correlation with other soluble molecules belonging to nerve growth factor receptors (NGFr) superfamily. Haematologica. 1998;83:398–402.
Rozovski U, Grgurevic S, Bueso-Ramos C, Harris DM, Li P, Liu Z, et al. Aberrant LPL expression, driven by STAT3, mediates free fatty acid metabolism in CLL cells. Mol Cancer Res MCR. 2015;13:944–53.
Robak T, Robak P. BCR signaling in chronic lymphocytic leukemia and related inhibitors currently in clinical studies. Int Rev Immunol. 2013;32:358–76.
Gujral TS, MacBeath G. A system-wide investigation of the dynamics of Wnt signaling reveals novel phases of transcriptional regulation. PLoS One. 2010;5:e10024.
Fabbri G, Dalla-Favera R. The molecular pathogenesis of chronic lymphocytic leukaemia. Nat Rev Cancer. 2016;16:145–62.
Acknowledgements
The authors wish to thank Margarita Bekyrou and Stella Kaila for their technical contributions. Epigenomics sample analyses were conducted under contract by CBM (Cluster in Biomedicine) S.c.r.l., Trieste, Italy, an Illumina Certified Service Provider.
Additional members of the EnviroGenomarkers consortium:
Ralph Gottschalk1, Danitsja van Leeuwen1, Leen Timmermans1, Maria Botsivali2, Benedetta Bendinelli3, Rachel Kelly4, Lutzen Portengen5, Fatemeh Saberi-Hosnijeh5, Göran Hallmans6, Per Lenner6, Hector C. Keun7, Alexandros Siskos7, Toby J. Athersuch4,7, Manolis Kogevinas8, Euripides G. Stephanou9, Antonis Myridakis9, Lucia Fazzo10, Marco De Santis10, Pietro Comba10, Hannu Kiviranta11, Panu Rantakokko11, Riikka Airaksinen11, Päivi Ruokojärvi11, Mark Gilthorpe12, Sarah Fleming12, Thomas Fleming12, Yu-Kang Tu12, Bo Jonsson13, Thomas Lundh13, Wei J. Chen14, Wen-Chung Lee14, Chuhsing Kate Hsiao14, Kuo-Liong Chien14, Po-Hsiu Kuo14, Hung Hung14, Shu-Fen Liao14.
Affiliations: 1Dept. of Toxicogenomics, Maastricht University, 6229 ER, Maastricht, Netherlands; 2Institute of Biology, Medicinal Chemistry and Biotechnology, National Hellenic Research Foundation, Athens 11635, Greece; 3The Institute for Cancer Research and Prevention, 50141 Florence, Italy; 4MRC-HPA Centre for Environment and Health, Department of Epidemiology and Biostatistics, School of Public Health, Faculty of Medicine, Imperial College, London W2 1PG, UK; 5Institute for Risk Assessment Sciences, Utrecht University, Utrecht; 6Department of Biobank Research, and Occupational and Environmental Medicine, Department of Public Health and Clinical Medicine, Umeå University, 901 87 Umeå, Sweden; 7Biomolecular Medicine, Department of Surgery and Cancer, Faculty of Medicine, Imperial College London, Sir Alexander Fleming Building, South Kensington, London, SW7 2AZ, UK; 8IS Global alliance, Doctor Aiguader 88, 08003 Barcelona, Spain; 9University of Crete, Heraklion, Greece; 10Istituto Superiore di Sanita, Rome, Italy; 11National Institute for Health and Welfare, Kuopio, Finland; 12University of Leeds, UK; 13Lund University, Sweden; 14National Taiwan University, Taipei, Taiwan.
Funding
This work was supported by the European Union [Grant number 226756]. The funding body played no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Availability of data and materials
The datasets generated and analysed during the current study are not publicly available due restrictions imposed by Swedish legislation on the protection of personal data.
Author information
Authors and Affiliations
Consortia
Contributions
SAK, PG, JCSK and PV designed the research. IAB, BM, FS and DP organised the epidemiologic part of the work. PG, DGH, JK, IL and IV performed the experiments and analysed the data. The EnviroGenomarkers consortium contributed to the design and conduct of the research and the evaluation of the results. SAK, AC, TMCMK, JCSK, RCHV, JV, MC-H and PV supervised the research. SAK and PG wrote and all authors commented on the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The EnviroGenomarkers project and its associated studies and experimental protocols were approved by the Regional Ethical Review Board of the Umeå Division of Medical Research, for the Swedish cohort, and the Florence Health Unit Local Ethical Committee, for the Italian cohort. All participants gave written informed consent.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1:
Text. Results on WBC composition in case and control subjects, omic profiles in different subgroups of subjects and assessment of the profile robustness across the cohorts. (PDF 614 kb)
Additional file 2: Table S1.
Relative risk and TtD of case sub-groups with different B-cell fraction. Table S2. Numbers of differential signals observed in different subject groups. Table S3. Top 100 DM CpGs. Table S4. Genes and CGI’s with at least three DM CpG sites and 20% enrichment. Table S5. CLL risk-related DE probes (FDR < 0.05). Table S6. CLL risk-associated DM and DE signals in cases with <10% B-cells compared to controls with <10% B-cells; Table S7. Top 100 DM and DE signals significant at FDR < 0.05 in the long and short TtD subgroups. Table S8. Methylation multiplicity, in the long and short TtD subgroups. Table S9. Ontology terms and pathways derived from the DM genes. Table S10. Ontology terms and pathways derived from the DE genes. Table S11. Ontology terms and pathways derived from the DM genes significant in different TtD subgroups. Table S12. Ontology terms and pathways derived from the DE genes significant in the short TtD subgroup. Table S13. CLL risk-related DM & DE hub genes. Table S14. DM and DE hub genes significant in different TtD subgroups. (XLSX 115 kb)
Additional file 3: Figure S1.
The ontological tree based on DM genes. Figure S2. Methylation of HOX and IRX gene clusters in the short- and long-TtD subgroups. Figure S3. B-cell distribution in controls and CLL cases. (PDF 1281 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Georgiadis, P., Liampa, I., Hebels, D.G. et al. Evolving DNA methylation and gene expression markers of B-cell chronic lymphocytic leukemia are present in pre-diagnostic blood samples more than 10 years prior to diagnosis. BMC Genomics 18, 728 (2017). https://doi.org/10.1186/s12864-017-4117-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-017-4117-4