
Abstract
Background
Although Tellinidae is one of the largest and most diverse families of bivalves, its taxonomy is utterly chaotic. This is mainly due to the morphological diversity and homoplasy displayed by their shells and to the scarcity of the molecular phylogenetic studies performed on them. A molecular cytogenetic analysis of four tellin shell species, Bosemprella incarnata, Macomangulus tenuis, Moerella donacina and Serratina serrata, was performed. To molecularly characterize the analyzed specimens, the sequence of a fragment of the mitochondrial cytochrome c oxidase subunit I (COI) was also studied.
Results
The karyotypes of the four species were composed of different amounts of bi-armed and telocentric chromosomes. The chromosomal mapping of 45S and 5S rDNA and H3 histone gene clusters by fluorescent in situ hybridization also revealed conspicuous differences on the distribution of these DNA sequences on their karyotypes. Vertebrate type telomeric sequences were located solely on both ends of each chromosome in all four tellin shells.
Conclusion
We present clear evidence of the valuable information provided by FISH signals in both analyzing chromosome evolution in Tellinidae and as a further tool in identifying tellin shell specimens for molecular phylogenies.
Similar content being viewed by others
Background
Tellin shells are fast-burrowing bivalves inhabiting marine and estuarine soft bottom ecosystems; they are distributed worldwide but are particularly abundant at tropical latitudes [1]. With over 550 living species organized in nine subfamilies and 107 genera, Tellinidae (Blainville, 1814) is one of the largest and most diverse families of bivalves [2]. In spite of that, the taxonomy of tellin shells is utterly chaotic, mainly as a consequence of the morphological diversity and homoplasy displayed by their shells [1, 2]. The situation is further aggravated by the scarcity of molecular phylogenetic studies on this family [2]. The few molecular analyses including Limecola and Serratina sequences [3,4,5] have also challenged traditional systematics and demonstrated improper placement and/or misidentification of some of the analyzed specimens. Therefore, to overcome these difficulties and clarify the classification of tellin shells further identification criteria are required in order to integrate traditional cladistic and molecular phylogenetic studies.
Although chromosome analyses have helped in both characterizing and solving identification problems in some groups of bivalves [6, 7], cytogenetic data in tellin shells is rather scarce and mostly limited to the description of diploid chromosome numbers (2n = 38) and karyotypes in three species, Macomangulus tenuis [8], Limecola balthica [8, 9] and Macoma nasuta [10], and to the location of the nucleolar organizing regions (NORs) by silver staining in L. balthica [11] and by fluorescent in situ hybridization (FISH) in M. nasuta [10]. Additionally, chromosome analysis has also been applied to study neoplastic cells in L. balthica [11,12,13,14].
To verify if a cytogenetic approach can help in solving some of the identification problems in tellin shells, in this work we characterized the chromosomes of four Tellinidae taxa by means of chromomycin A3 (CMA), 4′,6-diamidino-2-phenylindole (DAPI) and propidium iodide (PI) fluorescence staining and FISH mapping telomeric sequences and 45S rDNA, 5S rDNAs and H3 histone gene clusters. A fragment of the mitochondrial cytochrome c oxidase subunit I (COI) gene was also amplified and sequenced in the same tellin shell specimens.
Methods
Tellin shell specimens
Tellin shell specimens were collected in Ría de Pontevedra (NW Spain). The specimens were identified as Moerella donacina (Linnaeus, 1758), Serratina serrata (Brocchi, 1814), Macomangulus tenuis (da Costa, 1778) and Bosemprella incarnata (Linnaeus, 1758) according to shell morphology criteria. Representative specimens of the four taxa studied appear in Fig. 1. The nomenclature used for the taxa follows the World Register of Marine Species database (http://www.marinespecies.org/). To promote somatic growth, tellin shells were maintained in adequate laboratory conditions for 1 week [15].

Shells of studied specimens of the family Tellinidae. Representative shells of (a) Bosemprella incarnata, (b) Macomangulus tenuis, (c) Moerella donacina and (d) Serratina serrata. Scale bars, 1 cm
Chromosome preparation and fluorochrome staining
Following an overnight colchicine (0.005%) treatment, tellin shells were euthanized and gill and gonadic tissues dissected and immersed in 50% (20 min) and 25% (20 min) sea water prior to fixation with ethanol/acetic acid following previously described methods for cytogenetic studies in bivalves [16]. Chromosome spreads were obtained by disaggregating small pieces of tissue in 60% acetic acid and dropping the resulting cellular suspension onto preheated slides [17, 18].
After verifying the quality of the preparations by phase contrast microscopy, sequential fluorochrome staining was then performed [19,20,21]. Briefly, chromosomes were stained with CMA (0.25 mg/mL, 2 h) and DAPI (0.14 μg/mL, 8 min), mounted with antifade (Vectashield, Vector) and examined by fluorescent microscopy. For every single metaphase plate, images for each fluorochrome were acquired, pseudocolored and merged. After washing and staining with DAPI and PI (0.07 μg/mL, 8 min), the same metaphase plates were recorded again.
DNA isolation, PCR amplification and probe labeling
Genomic DNA was extracted with the EZNA Mollusc DNA Kit (Omega) following manufacturer’s instructions. A BioDrop μLITE (Biodrop) was employed to assess the purity and the concentration of the genomic DNA samples.
FISH probes were obtained by amplifying a 28S rDNA fragment with primers LR10R and LR12 [22], the entire 5S rDNA repeat with primers described by Pérez-García et al. [19] and the H3 histone gene with those reported by Giribet and Distel [23]. To molecularly characterize the studied tellin shells, a fragment of the mitochondrial COI gene was amplified in all karyotyped specimens using the primers developed by Nantón et al. [24] for Donacidae.
PCRs were carried out in a GeneAmp PCR system 9700 (Applied Biosystems) in 20 μL volumes containing 50 ng DNA, 1× reaction buffer, 2.5 mM MgCl2, 0.5 mM each dNTP (Thermo Fisher Scientific), 1 μM each primer and 1 U BIOTAQ DNA polymerase (Bioline). The amplification of the 28S rDNA included 30 cycles of 20 s denaturation at 95 °C, 20 s annealing at 48 °C and 30 s extension at 72 °C. For 5S rDNA, 40 cycles (95 °C, 44 °C and 72 °C, 20 s each) of amplification were employed. H3 histone gene amplification used 30 cycles (95 °C, 48 °C and 72 °C for 15 s each). For COI gene 35 amplification cycles (95 °C, 48 °C and 72 °C, 30 s each) were employed. All PCR reactions included an initial denaturation at 95 °C (10 min) and a final extension at 72 °C (5 min).
28S rDNA fragments were labeled either with biotin-16-dUTP (Roche) or digoxigenin-11-dUTP (Roche) by nick translation. Labeling of 5S rDNA and H3 histone gene probes was directly performed by supplementing the PCR mixture with either biotin-16-dUTP (20 μM) or digoxigenin-11-dUTP (5 μM).
DNA sequencing
COI PCR products were purified (FavorPrep™ GEL/PCR Purification Kit, Favorgen) and those corresponding to two specimens per taxa were sequenced in both directions using a BigDye terminator kit v3.1 (Applied Biosystems) in an ABI PRISM 3730 Genetic Analyzer (Applied Biosystems) by the genomic service (CACTI) of the University of Vigo. DNA sequences were edited with BioEdit v. 7.1.11 [25], aligned with MUSCLE in MEGAv7 [26] and trimmed to 545 bp after excluding primers.
Fluorescent in situ hybridization (FISH)
Single, double and sequential FISH were performed following previously described protocols [21, 27, 28]. Chromosome spreads were digested with RNase and pepsin before denaturating them in 70% formamide (70 °C, 2 min). Hybridization was performed overnight at 37 °C. Biotin detection was carried out with fluorescein isothiocyanate (FITC) conjugated avidin and biotinylated anti-avidin (Vector), whilst probes labeled with digoxigenin were detected with mouse anti-digoxigenin and anti-mouse tetramethylrhodamine (TRITC) antibodies. Chromosome preparations were counterstained with DAPI. After employing a commercial (C3TA2)3 peptide nucleic acid (PNA) probe (Applied Biosystems) for FISH mapping vertebrate telomeric repeats, chromosome preparations were counterstained with DAPI and PI. As indicated above, separated images for each fluorochrome were recorded, pseudo-colored and merged. For karyotyping and mapping purposes, at least 10 specimens per taxa (5 female, 5 male) and 20 metaphase plates per specimen were surveyed.
Results
All specimens of Bosemprella incarnata, Macomangulus tenuis, Moerella donacina and Serratina serrata are 2n = 38. As shown in Fig. 2, these tellin shells present diverse karyotype compositions. Bosemprella incarnata exhibits three metacentric, nine submetacentric, two subtelocentric and five telocentric chromosome pairs. Macomangulus tenuis karyotype is composed of one metacentric, eight submetacentric and ten subtelocentric pairs. In Moerella donacina seven metacentric, five submetacentric, six subtelocentric and one telocentric chromosome pairs appear. Lastly, Serratina serrata showed a karyotype composed of four metacentric, six submetacentric, three subtelocentric and six telocentric chromosome pairs.

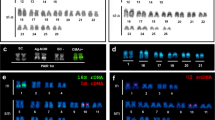
Mapping of rDNA and histone gene clusters to the chromosomes of four species of tellin shells counterstained with DAPI. FISH mapping of 5S rDNA (5S, red), 28S rDNA (28S, magenta) and H3 histone gene (H3, green) probes to mitotic metaphase plates, and corresponding karyotypes, of Bosemprella incarnata, Macomangulus tenuis, Moerella donacina and Serratina serrata. Scale bars, 5 μm
The combined use of AT-specific (DAPI), GC-specific (CMA) and unspecific fluorochromes revealed DAPI-dull/CMA-bright, GC-rich regions close to the centromeres in a single chromosome pair in Bosemprella incarnata, Macomangulus tenuis and Moerella donacina, and in two chromosome pairs in Serratina serrata. FISH experiments employing a 28S rDNA probe (Fig. 2) demonstrated that these GC-rich regions are coincident with the location of the 45S rDNA clusters.
A single H3 histone gene cluster was present in all four tellin shell species analyzed (Fig. 2). This cluster was subcentromeric to the short arms of both telocentric chromosome pair 15 in Bosemprella incarnata and submetacentric chromosome pair 3 in Serratina serrata and to the short arms of telocentric pair 17 in Moerella donacina. In contrast, H3 histone genes were subterminal to the long arms of subtelocentric pair 6 in Macomangulus tenuis.
Single 5S rDNA clusters mapped (Fig. 2) to subterminal positions on the long arms of metacentric pair 12 in Bosemprella incarnata and telocentric pair 9 in Serratina serrata. The remaining two tellin shells presented two 5S rDNA clusters. Subterminal to the long arms of telocentric pair 9 and subcentromeric to the long arms of subtelocentric pair 17 in Macomangulus tenuis; subterminal to the long arms of subtelocentric pair 5 and subcentromeric to the long arms of metacentric pair 7 in Moerella donacina.
Double FISH experiments using 28S and 5S rDNA probes labeled differently demonstrated that subtelocentric chromosome pair 9 of Macomangulus tenuis bears both subcentromeric 45S rDNA and subtelomeric 5S rDNA clusters in its long arms.
FISH analysis employing a telomeric (C3TA2)3 PNA probe revealed terminal signals at both ends of every chromosome in the four tellin shells.
A schematic representation of the cytogenetic results obtained is showed in Fig. 3 and a summary of the published cytogenetic data for Tellinidae species is presented in Table 1.

Ideogrammatic karyotypes of tellin shells. Schematic representation of the karyotypes of (a) Bosemprella incarnata, (b) Macomangulus tenuis, (c) Moerella donacina and (d) Serratina serrata. The schematic representation of the chromosomal mapping results shows H3 histone genes in green, 5S rDNA in red and 45S rDNA (NORs) in magenta
A fragment of the mitochondrial COI gene was successfully amplified in the four Tellinidae taxa and the corresponding sequences stored in GenBank under accession numbers KY951457 and KY951458 for Bosemprella incarnata, KY951461 and KY951462 for Macomangulus tenuis, KY951455 and KY951456 for Moerella donacina and KY951459 and KY951460 for Serratina serrata. All partial COI gene sequences were independently compared using BLAST with those stored in NCBI GenBank and BOLD databases. Out of these, high similarity levels (>99%) were only found among our Macomangulus tenuis sequences (Genbank accession numbers KY951461 and KY951462) and a single sequence (Genbank accession number KR084511) from a North Sea specimen stored under the same specific assignation. The same comparison approaches using all the other sequences obtained in this work always displayed levels of similarity (<93%) well below those usually corresponding to the same taxa in bivalves.
Discussion
The diploid chromosome numbers, 2n = 38, found for the four tellin shell species studied here coincide with those previously reported for Macomangulus tenuis [8] and the other two Tellinidae species previously described, Limecola balthica [8, 9] and Macoma nasuta [10]. This is also the usual diploid number in most Heterodonta bivalve species [21, 28,29,30,31,32,33,34,35,36].
In regards to karyotype composition, the main difference with previous data for Tellinidae (Table 1) is the occurrence of telocentric chromosome pairs in three of the four tellin shells studied here, a relatively unusual condition in Heterodonta [34]. This is not the case for Macomagulus tenuis whose karyotype, in concordance with previous results [8], is exclusively composed of bi-armed chromosomes; the minor differences between the karyotype composition reported here and the previously published one (Table 1) can be attributed to methodological variation.
As in most other bivalves studied to date [6, 7, 19,20,21, 27, 28, 31,32,33,34,35,36], the application of diverse combinations of base-specific fluorochromes to the staining of tellin shell chromosomes demonstrated that GC-rich regions are scarce, two in Serratina serrata and a single one in the other three species. As is also the rule for most bivalve species, further FISH using 28S rDNA probes confirmed that these GC-rich regions are coincident with the chromosomal regions bearing 45S rDNA clusters, the NORs.
Tandemly repeated DNA sequences are helpful markers for identifying chromosomes. This is also the case in bivalves in which FISH mapping made possible establishing more reliable karyotypes and identifying some of the chromosomal changes accompanying bivalve evolution [6]. As shown in Table 1, this kind of approach has barely been applied to tellin shells. Only 45S rDNAs have been previously located, either by silver staining (Limecola balthica) [11] or FISH (Macoma nasuta) [10] in Tellinidae. The presence a single 45S rDNA subterminal in a chromosome pair in Macoma nasuta [10] coincided in number with our results for Bosemprella incarnata, Macomangulus tenuis and Moerella donacina and differed with the two clusters displayed by Serratina serrata. In regards to location, the situation is quite different because most of the 45S rDNA clusters found in this work were subcentromeric; this was also the case for one of the three silver-stained NORs found in Limecola balthica [11].
Regarding 5S rDNA and histone gene clusters, no previous data was available for any of the species of Tellinidae. The presence of one or two 5S rDNA clusters has also been described in another Heterodonta family, Veneridae, in which subcentromeric, intercalary and subtelomeric locations were also found [7, 21]. The occurrence of single minor histone gene clusters, either subterminal or subcentromeric, in all tellin shells are similar to the situation found in Mactridae [28] but differs with that in Veneridae in which 1 to 4 clusters have been described [7, 21, 33].
Additionally, the simultaneous presence of both 45S and 5S rDNA clusters on the same chromosome pair in Macomangulus tenuis is rather scarce in bivalves; Ruditapes decussatus, showing overlapping 45S and 5S rDNA signals, is the only other Heterodonta species presenting this particularity [36] that was also found in three Mytilidae, Perumytilus purpuratus [19], Brachidontes rodriguezii [20] and Mytilus trossulus [6].
Concerning telomeric sequences, their presence exclusively at chromosome ends in Bosemprella incarnata, Macomangulus tenuis, Moerella donacina and Serratina serrata is the most common result found in bivalves [19,20,21, 27, 28, 32,33,34,35].
Despite their abundance and ecological importance, the systematics and taxonomy of tellin shells, mostly based on characters showing an extraordinarily high level of morphological homoplasy, is not a realistic reflection of the true phylogenetic relationships of its members and requires a profound revision [2]. This is further confirmed by the most recent molecular phylogeny of bivalves that, although including scarce tellin shell sampling, displays Tellinidae, and other Tellinoida families, as polyphiletic and highlights the need of denser taxon sampling in clades, such as Tellinoida, with poorly defined families [37]. This denser sampling has to be accompanied by an accurate determination of the specimens using, as proposed by integrative taxonomy [38, 39], a large number of characters to identify them. The information provided by the chromosomal location of the FISH signals can facilitate this task.
Conclusion
In summary, the results presented in this work clearly demonstrated that molecular karyotype is a useful character in tellin shell identification, therefore being helpful for both taxonomic and phylogenetic purposes.
Abbreviations
- CMA:
-
Chromomycin A3
- DAPI:
-
4′,6-diamidino-2-phenylindole
- FISH:
-
Fluorescence in situ hybridization
- NOR:
-
Nucleolus organizing region
- PCR:
-
Polymerase chain reaction
- PI:
-
Propidium iodide
- rDNA:
-
Ribosomal DNA
- rRNA:
-
Ribosomal RNA
References
Simone LRL, Wilkinson S. Comparative morphological study of some Tellinidae from Thailand (Bivalvia: Tellinoidea). Raffles Bull Zool. 2008;18:151–90.
Huber M. Compendium of bivalves 2. A full-color guide to the remaining seven families. A systematic listing of 8′500 bivalve species and 10′500 synonyms. Hackenheim: ConchBooks; 2015.
Taylor JD, Williams ST, Glover EA, Dyal P. A molecular phylogeny of heterodont bivalves (Mollusca: Bivalvia: Heterodonta): new analyses of 18S and 28S rRNA genes. Zool Scr. 2007;36:587–606.
Saunier A, Garcia P, Becquet V, Marsaud N, Escudié F, Pante E. Mitochondrial genomes of the Baltic clam Macoma balthica (Bivalvia: Tellinidae): setting the stage for studying mito-nuclear incompatibilities. BMC Evol Biol. 2014;14:259.
Sun S, Li Q, Kong L, Yu H, Zheng X, Yu R, Dai L, Sun Y, Chen J, Liu J, Ni L, Feng Y, Yu Z, Zou S, Lin J. DNA barcoding reveal patterns of species diversity among northwestern Pacific molluscs. Sci Rep. 2016;6:33367.
Pérez-García C, Morán P, Pasantes JJ. Karyotypic diversification in Mytilus mussels (Bivalvia: Mytilidae) inferred from chromosomal mapping of rRNA and histone gene clusters. BMC Genet. 2014;15:84.
García-Souto D, Qarkaxhija V, Pasantes J. Resolving the taxonomic status of Chamelea gallina and C. striatula (Veneridae, Bivalvia): A combined molecular cytogenetic and phylogenetic approach. Biomed Res Int. 2017;2017, Article ID 7638790:8. https://doi.org/10.1155/2017/7638790.
Cornet M, Soulard C. Karyometric analysis of two species of the family Tellinidae: Macoma balthica (L.) and Tellina tenuis da Costa (Mollusca, Bivalvia, Tellinacea). Cytobios. 1990;64:93–101.
Wolowicz M, Thiriot-Quievreux C. The karyotypes of the most common bivalve species from the South Baltic. Oceanol Stud. 1997;2-3:209–21.
González-Tizón AM, Martínez-Lage A, Rego I, Ausio J, Méndez J. DNA content, karyotypes, and chromosomal location of 18S-5.8S-28S ribosomal loci in some species of bivalve molluscs from the Pacific Canadian coast. Genome. 2000;43:1065–72.
Smolarz K, Wolowicz M, Thiriot-Quiévreux C. Argyrophilic nucleolar organizer regions (AgNORs) in interphases and metaphases of normal and neoplastic gill cells of Macoma balthica (Bivalvia: Tellinidae) from the Gulf of Gdansk, Baltic Sea. Dis Aquat Org. 2003;56:269–74.
Thiriot-Quiévreux C, Wolowicz M. Etude caryologique d’une néoplasie branchiale chez Macoma balthica (Mollusca, Bivalvia). CR Acad Sci Sér III Sci Vie. 1996;319:887–92.
Thiriot-Quiévreux C, Wolowicz M. Chromosomal study of spatial variation of the prevalence of a gill neoplasia in Macoma balthica from the Gulf of Gdansk (Baltic Sea). Ophelia. 2001;54:75–81.
Smolarz K, Renault T, Wolowicz M. Histology, cytogenetics and cytofluorymetry in diagnosis of neoplasia in Macoma balthica (Bivalvia, L.) from the southern Baltic Sea. Caryologia. 2005;58:212–9.
Pasantes J, Martínez-Expósito MJ, Martínez-Lage A, Méndez J. Chromosomes of Galician mussels. J Molluscan Stud. 1990;56:123–6.
Méndez J, Pasantes JJ, Martínez-Expósito MJ. Banding pattern of mussel (Mytilus galloprovincialis) chromosomes induced by 2xSSC/Giemsa-stain treatment. Mar Biol. 1990;106:375–7.
Martínez-Expósito MJ, Pasantes JJ, Méndez J. Proliferation kinetics of mussel (Mytilus galloprovincialis) gill cells. Mar Biol. 1994;120:41–5.
Martínez-Expósito MJ, Pasantes JJ, Méndez J. NOR activity in larval and juvenile mussels (Mytilus galloprovincialis Lmk.). J Exp Mar Biol Ecol. 1994;175:155–65.
Pérez-García C, Cambeiro JM, Morán P, Pasantes JJ. Chromosomal mapping of rDNAs, core histone genes and telomeric sequences in Perumytilus purpuratus (Bivalvia: Mytilidae). J Exp Mar Biol Ecol. 2010;395:199–205.
Pérez-García C, Guerra-Varela J, Morán P, Pasantes JJ. Chromosomal mapping of rRNA genes, core histone genes and telomeric sequences in Brachidontes puniceus and Brachidontes rodriguezi (Bivalvia: Mytilidae). BMC Genet. 2010;11:109.
García-Souto D, Pérez-García C, Morán P, Pasantes J. Divergent evolutionary behavior of H3 histone gene and rDNA clusters in venerid clams. Mol Cytogenet. 2015;8:40.
Vilgalys R. Conserved primer sequences for PCR amplification and sequencing from nuclear ribosomal RNA. http://lutzonilab.org/nuclear-ribosomal-dna/.
Giribet G, Distel D. Bivalve phylogeny and molecular data. In: Lydeard C, Lindberg DR, editors. Systematics and Phylogeography of Molluscks. Washington DC: Smithsonian Books; 2003. p. 45–90.
Nantón A, Freire R, Arias-Pérez A, Gaspar MB, Méndez J. Identification of four Donax species by PCR–RFLP analysis of cytochrome c oxidase subunit I (COI). Eur Food Res Technol. 2015;240:1129–33.
Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp Ser. 1999;41:95–8.
Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33:1870–4.
Pérez-García C, Morán P, Pasantes JJ. Cytogenetic characterization of the invasive mussel species Xenostrobus securis Lmk. (Bivalvia: Mytilidae). Genome. 2011;54:771–8.
García-Souto D, Pérez-García C, Kendall J, Pasantes J. Molecular cytogenetics in trough shells (Mactridae, Bivalvia): divergent GC-rich heterochromatin content. Genes. 2016;7:47.
Nakamura HK. A review of molluscan cytogenetic information based on the CISMOCH-computerized system for molluscan chromosomes. Bivalvia, Polyplacophora and Cephalopoda. Venus (Jap J Malacol). 1985;44:193–225.
Thiriot-Quiévreux C. Advances in cytogenetics of aquatic organisms. In: Beaumont AR, editor. Genetics and evolution of aquatic organisms. London: Chapman and Hall; 1994. p. 369–88.
Thiriot-Quiévreux C. Review of the literature on bivalve cytogenetics in the last 10 years. Cah Biol Mar. 2002;43:17–26.
Leitão A, Chaves R: Banding for chromosomal identification in bivalves: A 20-year history. In: Aquaculture 1. Dynamic Biochemistry, Process Biotechnology and Molecular Biology 2 (Special Issue 1). Edited by Russo R. Global Science Books; 2008:44–49. http://www.globalsciencebooks.info/Journals/images/GSB_Journal_Information_Stats.pdf.
Carrilho J, Pérez-García C, Leitão A, Malheiro I, Pasantes JJ. Cytogenetic characterization and mapping of rDNAs, core histone genes and telomeric sequences in Venerupis aurea and Tapes rhomboides (Bivalvia: Veneridae). Genetica. 2011;139:823–30.
González-Tizón AM, Rojo V, Vierna J, Jensen T, Egea E, Martínez-Lage A. Cytogenetic characterisation of the razor shells Ensis directus (Conrad, 1843) and E. minor (Chenu, 1843) (Mollusca: Bivalvia). Helgol Mar Res. 2013;67:73–82.
Pérez-García C, Hurtado NS, Morán P, Pasantes JJ. Evolutionary dynamics of rDNA clusters in chromosomes of five clam species belonging to the family Veneridae (Mollusca, Bivalvia). Biomed Res Int. 2014;2014:754012.
Hurtado N, Pérez-García C, Morán P, Pasantes JJ. Genetic and cytological evidence of hybridization between native Ruditapes decussatus and introduced Ruditapes philippinarum (Mollusca, Bivalvia, Veneridae) in NW Spain. Aquaculture. 2011;311:123–8.
Combosch DJ, Collins TM, Glover EA, Graf DL, Harper EM, Healy JM, Kawauchi GY, Lemer S, McIntyre E, Strong EE, Taylor JD, Zardus JD, Mikkelsen PM, Giribet G, Bieler R. A family-level tree of life for bivalves based on a Sanger-sequencing approach. Mol Phylogenet Evol. 2017;107:191–208.
Dayrat B. Towards integrative taxonomy. Biol J Linn Soc Lond. 2005;85:407–15.
Will KP, Mishler BD, Wheeler QD. The perils of DNA Barcoding and the need for integrative taxonomy. Syst Biol. 2005;54:844–51.
Acknowledgements
We wish to thank Ben Moore for revising the English text. D. García-Souto was partially supported by a FPU fellowship from “Ministerio de Educación, Cultura y Deporte” (Spain).
Funding
This work was partly funded by a grant from Xunta de Galicia and Fondos FEDER: “Unha maneira de facer Europa” (Axudas do programa de consolidación e estruturación de unidades de investigacións competitivas do SUG: ED431C 2016–037).
Availability of data and materials
Authors can confirm that all relevant data are included in the article.
Author information
Authors and Affiliations
Contributions
DGS performed most experiments and collaborated in the discussion of the data and the writing of the manuscript. GR participated in the experimental work and contributed to the discussion of data. JJP coordinated the study, discussed the data and wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable. The animals studied in this work are bivalve mollusks, non-cephalopod invertebrates.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
García-Souto, D., Ríos, G. & Pasantes, J.J. Karyotype differentiation in tellin shells (Bivalvia: Tellinidae). BMC Genet 18, 66 (2017). https://doi.org/10.1186/s12863-017-0535-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12863-017-0535-1