
Abstract
Background
Skeletal muscle injury activates satellite cells to initiate processes of proliferation, differentiation, and hypertrophy in order to regenerate muscle fibers. The number of microRNAs and their target genes are engaged in satellite cell activation. β-Hydroxy-β-methylbutyrate (HMB) is known to prevent exercise-induced muscle damage. The purpose of this study was to evaluate the effect of HMB on miRNA and relevant target gene expression in differentiating equine satellite cells exposed to H2O2. We hypothesized that HMB may regulate satellite cell activity, proliferation, and differentiation, hence attenuate the pathological processes induced during an in vitro model of H2O2-related injury by changing the expression of miRNAs.
Methods
Equine satellite cells (ESC) were isolated from the samples of skeletal muscle collected from young horses. ESC were treated with HMB (24 h) and then exposed to H2O2 (1 h). For the microRNA and gene expression assessment microarrays, technique was used. Identified miRNAs and genes were validated using real-time qPCR. Cell viability, oxidative stress, and cell damage were measured using colorimetric method and flow cytometry.
Results
Analysis of miRNA and gene profile in differentiating ESC pre-incubated with HMB and then exposed to H2O2 revealed difference in the expression of 27 miRNAs and 4740 genes, of which 344 were potential target genes for identified miRNAs. Special attention was focused on differentially expressed miRNAs and their target genes involved in processes related to skeletal muscle injury. Western blot analysis showed protein protection in HMB-pre-treated group compared to control. The viability test confirmed that HMB enhanced cell survival after the hydrogen peroxide exposition.
Conclusions
Our results suggest that ESC pre-incubated with HMB and exposed to H2O2 could affect expression on miRNA levels responsible for skeletal muscle development, cell proliferation and differentiation, and activation of tissue repair after injury. Enrichment analyses for targeted genes revealed that a large group of genes was associated with the regulation of signaling pathways crucial for muscle tissue development, protein metabolism, muscle injury, and regeneration, as well as with oxidative stress response.
Similar content being viewed by others

Background
β-Hydroxy-β-methylbutyrate (HMB) is a metabolite of the essential amino acid leucine and is naturally synthesized in animals, plants, and humans [1]. Dietary supplementation of HMB is used to enhance gains in strength and lean body mass associated with resistance training and for increasing lean mass in cancer-related cachexia [2, 3]. Unlike anabolic hormones which only increase muscle protein synthesis to accelerate muscular hypertrophy, HMB increases dynamic strength [4, 5] and lean body mass [6] acting as an anti-catabolic agent, reducing protein breakdown [5] and cellular damage which may accompany intense exercise [7]. Moreover, previous studies have demonstrated that HMB supplementation decreased plasma post-exercise creatine kinase and lactic acid in thoroughbreds [8].
Reactive oxygen species (ROS), such a hydrogen peroxide (H2O2), exert a critical regulatory role on skeletal muscle function [9, 10]. In resting muscle cells, free radicals and ROS are rapidly and efficiently neutralized by antioxidants. Exercise creates an imbalance between ROS and activates natural antioxidant mechanisms. Moreover, ROS produced during exercise by inflammatory cells may also be involved in delayed onset of muscle damage observed during inflammation [11]. The inflammatory response coincides with muscle repair, regeneration, and growth, involving activation and proliferation of satellite cells followed by their terminal differentiation. In response to the damage, quiescent satellite cells are activated and undergo several cycles of cell division prior to their withdrawal from the cell cycle through terminal differentiation and finally fusion with the damaged skeletal muscle fibers [12]. During training-related tissue microdamage, activation of satellite cells is considered to play a crucial role in injured muscle fibers by incorporating new myonuclei and thus increasing muscle size and strength (by hypertrophy) [13].
MicroRNAs (miRNAs) are small non-coding interfering RNA molecules (18–25 nucleotides) able to post-transcriptionally regulate gene expression through sequence-specific base pairing to messenger ribonucleic acid (mRNA). These molecules have been shown to be important key players in a variety of physiological and pathological processes (proliferation, differentiation, apoptosis, hypertrophy, timing development, inflammation, cancer, etc.). A group of miRNAs, highly enriched in skeletal and/or cardiac muscles (myomiRs), has recently been identified and includes miR-1, miR-133a, miR-133b, miR-206, miR-208, miR-208b, miR-486, and miR-499 [14] which regulate skeletal muscle development.
Szcześniak et al. [15] were the first who demonstrated the effect of HMB in ESC. Our study was performed to evaluate miRNA profile and relevant target genes in differentiating equine satellite cells incubated with HMB and also exposed to H2O2 an in vitro factor initiating cellular response similar to that observed in vivo during a short intensive physical exercise and post-exercise injury.
Methods
Muscle samples and cell culture
Samples of skeletal muscles (m. semitendinosus) were collected from 6 months old healthy stallions in a slaughter house. Muscle samples (0.5 × 0.5 × 0.5 cm) were taken immediately, washed in phosphate-buffered saline (PBS) with gradually decreasing antibiotic concentration [40.000 and 20.000 IU Penicillium crystalicum (PC; Polfa, Poland) per 100 ml PBS], cleaned from connective and fat tissue, cut and immediately suspended in sterile fetal bovine serum (FBS; Life Technologies, USA) with 10% addition of dimethylsulfoxide (DMSO), gradually frozen to − 80 °C, and finally stored in liquid nitrogen until use.
Satellite cell isolation, proliferation, and differentiation
Equine satellite cells (ESC) were isolated according to the following protocol. Protease from Streptomyces griseus (Pronase®, Sigma-Aldrich, USA) was reconstituted in low-glucose Dulbecco's modified Eagle’s medium (DMEM), GlutaMAX™, Pyruvate (Life Technologies, USA), and stirred for 1 h, pH 7.3. The incubation buffer (IB) consisted per sample of Pronase 0.5 mg/ml, 18 ml of DMEM, FBS 2 ml (Life Technologies, USA), and PC (20.000 IU). IB was filtered through a cellulose acetate membrane syringe filter (Sigma-Aldrich, USA). The fragmented muscle tissue was thawed, washed in PBS with PC (20.000 IU), and suspended in IB for 1.5 h at 37 °C, shaken every 15 min. Then, samples were sieved through cell strainer (70 μm, nylon, Falcon, USA). The filtrate was centrifuged for 20 min (350 g), which was repeated three times. After each centrifugation, supernatant was discarded, cell pellet was re-suspended in growth medium (GM; 10%FBS/10% horse serum (HS) in DMEM (Life Technologies, USA) and antibiotics (AB; 0.5% amphotericin B (Fungizone, Life Technologies, USA), 1% penicillin-streptomycin (Life Technologies, USA)). After the last centrifugation, cell suspension was transferred to polystyrene Petri dishes (Becton Dickinson, USA) for 1.5 h to allow adhesion of fibroblast. After that, supernatant with satellite cells was transferred into culture dishes (Primaria Cell Culture Flask, Becton Dickinson, USA) and cultured in GM. The growth medium was changed every 2 days. On the tenth day of proliferation, cells were trypsinized, counted by Scepter Cell Counter (Merck Millipore, Germany), transferred (30,000 cells from each isolation) to Collagen I Cellware six-well plate (Greiner Bio-One, USA), and cultured in GM. After reaching 80% confluency, the proliferation media was replaced by the differentiation media (DM; 2%HS in DMEM with AB).
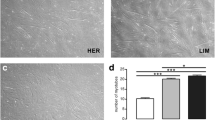
Primary satellite cell cultures from semitendinosus muscle of all horses were isolated, and the culture with the best scores of cell viability (MTT assay) [16] and fusion index was selected for further analysis (data not shown). Different stages of equine satellite cell culture are presented in Fig. 1.

Equine satellite cell culture. a Proliferating ESC, day 4. b Proliferating ESC, day 8. c Myotubes in differentiating ESC, day 2
Experimental design
After the second day of differentiation, 50 μM HMB (Metabolic Technologies Inc., USA) was added to the culture media, and then, cells were incubated for an additional 24 h. Ca-HMB was purchased from MTI (USA). The free HMB acid was extracted by acidification and organic extraction [8]. HMB dose was chosen based upon previous studies [3, 17] and MTT assay results which confirmed literature data (data not shown). During the last hour of incubation, 3 mM hydrogen peroxide (solution 30% (w/w) in H2O (Sigma-Aldrich, USA) was added to induce cell damage. Due to the lack of literature data on the doses of H2O2 used in the equine satellite cell culture, the MTT assay was performed using doses ranging for 0.125 to 50 mM. Compared to the previously described doses of H2O2 used in other cell culture models, those used for ESCs were relatively large. For this reason, we decided to use H2O2 dose 3 mM with DL-25 (Fig. 2). The experimental design is presented in Fig. 3.

H2O2 dose-dependent effect on ESC cell viability assessed by MTT assay

Experimental design. a Control group. b HMB-treated group
RNA isolation
After the H2O2 treatment, the differentiating ESCs were scraped and total RNA was isolated using a miRNeasy Mini Kit (Qiagen, USA) according to the manufacturer’s protocol. The quantity of RNA was measured spectrophotometrically using NanoDrop 2000 (Thermo Scientific, USA). The quality of the total RNA was verified by Bioanalyzer 2100 (Agilent, USA), and only samples with RIN ≥ 9.2 were used for further analysis.
Microarray analysis
For the microRNA profiling, the Custom Equine miRNA 8x15K Microarray slides were designed using eArray platform (https://earray.chem.agilent.com/earray, GEO database: GPL20990) and provided by Agilent Technologies (USA).
MiRNA was isolated from eight equine satellite cell cultures for both HMB pre-treated (n = 8) and control group (n = 8). As recommended by Agilent Technologies (USA), 100 ng of total RNA of each sample was taken and labeled using miRNA Complete Labeling and Hyb Kit (version 2.3, December 2010). For hybridization, Microarray Hybridization Chamber (Agilent, USA) and Hyb-Buffer (Agilent, USA) were used according to the manufacturer’s protocol. In the next step, slides were washed out using Gene Expression Wash Pack (Agilent, USA) and scanned in Microarray Scanner (model G2565CA) with SureScan High-Resolution Technology (Agilent, USA).
Microarray data were extracted, the background was subtracted, and normalization was performed using the standard procedures included in the Agilent Feature Extraction (FE) Software version 10.7.3.1.
Analysis of gene expression (GE) profile was performed using Horse Gene Expression Microarray, 4x44K (Agilent Technologies, USA) based on the same protocol as described by Szcześniak et al. (2016), [15, 18]. Briefly, two-color microarray, with 825 ng of cRNA from HMB-exposed cells (labeled by Cy5, n = 4) and 825 ng of cRNA from control cells (labeled by Cy3, n = 4), and RNA Spike-In Kit (Agilent Technologies, USA) as an internal control were used. The background was subtracted and Linear and Lowess normalization was performed using the standard procedures included in the Agilent Feature Extraction (FE) Software version 10.7.3.1. The data were statistically analyzed using Gene Spring 13.0 software (Agilent, USA). The statistical significance of the differences was evaluated using Student’s t test (p < 0.05) and Benjamini and Hochberg multiple testing correction. False Discovery Rate (FDR) ≤ 0.05 and fold change (FC) ≥ 1.3 were considered as statistically significant. Microarray data were deposited at the Gene Expression Omnibus data repository under the number GSE73779 for miRNA and GSE93025 for cDNA.
Real-time qPCR
The criteria for miRNA and differentially expressed gene (DEG) selection for real-time qPCR validation and further analysis were of biological relevance (miRNAs linked to muscular development, hypertrophy, muscle injuries, oxidative stress, and tissue regeneration) and were assessed based on Pathway Studio Mammalian (Elsevier, USA) and available literature.
For miRNA, real-time qPCR validation miRCURY LNA™ Universal RT microRNA PCR kit (Exiqon, USA) was used. A two-step protocol was applied: (1) polymerase activation at 95 °C for 10 min and (2) 40 amplification cycles at 95 °C for 10s and 60 °C for 1 min, according to the manufacturer protocol.
Primers were chosen based on the miRNA sequences assigned to microarray probes and were provided by Exiqon (Denmark) (Table 1). Calculation of the relative miRNA expression using the ΔΔCt method was applied using GenEX 6 software provided by MultiD (Sweden). Obtained data were statistically analyzed using two-tailed Student’s t test. Values of p ≤ 0.05 were considered statistically significant.
Based on previous studies in different species and the manufacturer recommendation (Exiqon, Denmark), a U6 snRNA reference was used. To verify GE microarray results, the real-time qPCR method was applied. All the steps of real-time qPCR procedure were made based on the protocols previously described by Szcześniak et al. (2016), [15]. The sequences of primers are listed in Table 2. Gapdh was used as a reference gene.
Target gene prediction and ontological analyses
MicroRNA target gene prediction was performed using the TargetScan database. The analysis was performed for all identified HMB-affected miRNAs. For each predicted target of individual miRNA, the sum of the context + scores was automatically calculated. Predicted targets of each miRNA family were automatically sorted by total context + score. Analysis was performed for the context score percentile (50) and conserved/non-conserved miRNA families and target sites [19]. For further analysis, common genes for those identified genes using GE microarray and predicted miRNA target genes were selected and considered as targets for HMB treatment-influenced miRNAs.
Ontological analyses revealing molecular functions, biological processes, and pathways of miRNA targets were performed in DAVID 6.7 using Fisher’s exact test with p ≤ 0.05. Detailed analysis of the role of HMB-modulated miRNAs, genes identified using GE, and target genes in various metabolic and signal pathways was performed using Pathway Studio Web (Elsevier, USA). Relationships between all differentially expressed miRNAs were visualized with Pathway Studio’s Build Pathway functionality which is based on the wave-propagation algorithm developed for the navigation through complex networks. Find Direct Links/All objects Directions Algorithm was used in this analysis.
Western blot analysis
The procedure of Western blot analysis was performed based on the previously described methodology by Zielniok et al. [20]. Antibodies used in Western blot were against the following: SOD1 (ab62800), SOD2 (ab13534), TGFβ2 (sc-90), α-tubulin (ab176560), BDNF (sc-546), MYF5 (sc-302), GAPDH (sc-20357), and β-actin (sc-47778).
Cell viability, cell damage, and oxidative stress
Hydrogen peroxide, used in the experiment as a damage factor, is known to affect various cellular processes. Several tests related to the cell viability, cell damage, and oxidative stress were performed to assess the impact of HMB on the cellular processes following incubation with H2O2. Experimental conditions (incubation time, doses of HMB and H2O2) were the same as previously in the part related to the microarray and real-time qPCR analysis.
CellROX® Green Reagent Kit (Life Technologies) was used to measure oxidative stress and cell death in ESC’s based on the manufacturer protocol. Cells were seeded on 24-well plates at 0.05 × 106 cells/cm2. Cells were incubated for 60 min with CellROX reagent in a final concentration 250 μM. During the last 15 min of staining, SYTOX Red Dead Cell was added (at the final concentration 5 nM). The samples were analyzed immediately after staining using FACS Aria II (BD Biosciences) flow cytometer. A total of 50,000 events per sample (n = 3) were collected. This staining was performed on live cells during the proliferative phase (90% confluency). Data were analyzed using FlowJo (TreeStar, USA) and GraphPad Prism software.
The second test related to oxidative stress called Total Antioxidant Capacity (TAC) Assay Kit (Abcam, UK) was used according to the manufacturer protocol. This test can measure either the combination of both small molecule antioxidants and proteins or small molecules alone in the presence of our proprietary Protein Mask. Cells were seeded on 96-well plates at 2 × 106 cells (n = 6). After a 90-min incubation, the plate was read on Tekan System reader at 570 nm wavelength. Data were analyzed using GraphPad Prism software.
Lipid peroxidation is the degradation of lipids that may accompany the activity of several cell damage factors including hydrogen peroxide. It is also one of the popular markers for oxidative stress. Lipid peroxidation Assay Kit (Sigma-Aldrich) was used to measure lipid peroxidation. All the procedure was performed based on the provided manufacturer protocol. The concentration of MDA was measured for n = 6. The staining was performed on live cells during differentiation phase.
In order to increase the reliability of the obtained results related to cell survival, the MTT test was also performed (n = 6) based on the previously published protocol [16]. Data for both tests were analyzed using GraphPad Prism software.
Qualitative flow cytometry assay for mitochondrial depolarization was also performed according to the manufacturer protocol. The 5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine iodide (JC-1, Sigma-Aldrich) was used. It is a cationic, lipophilic dye that accumulates in mitochondria and exhibits green fluorescence (525 nm) in its monomeric state. The mostly implemented application of JC-1 is detection of mitochondrial depolarization occurring in the early stages of apoptosis. JC-1 was dissolved in DMSO and medium II for a final concentration of 0.6 μM. The cells were incubated at 37 °C, washed, trypsinized, and resuspended in 2%FBS/PBS medium. Fifty thousand events were collected for each sample using FACS Aria II (BD Biosciences) flow cytometer. Fluorescence compensation was done for 525 nm. This staining was performed on live cells during the proliferative phase (90% of confluency; n = 3). Data were analyzed using FlowJo (TreeStar, USA) and GraphPad Prism software.
Results
In the “Results” and “Discussion” sections, gene symbols are marked in italics and lowercase. The arrows indicate the direction of expression change: ↓ and ↑ for down- and upregulation, respectively.
Microarray analysis
Analysis of the miRNA expression in differentiating equine satellite cells incubated with HMB (24 h) and exposed to H2O2 (1 h) revealed differences in 27 miRNAs. Among them, eight demonstrated higher expression and 19 lower expression when compared to control (Table 3).
Analysis of gene expression profile for the same experimental conditions as those mentioned above revealed difference in the expression of 4740 transcripts. After removing all duplicate values and unknown sequences, 1923 unique genes were found (Additional file 1: Table S1).
Functional analysis of identified miRNAs and differentially expressed genes (DEG)
Based upon the Pathway Studio Web Software (Elsevier, USA) and available literature, the results were divided into groups containing miRNAs related to the specific cellular processes, as follows: (1) cell proliferation and differentiation (miR-1, miR-133a/b, miR-206, miR-128, miR-146a/b, miR-204, miR-155, miR-193a, miR-221/222, miR-324, miR-331, miR-374b, miR-486, miR-675), (2) muscle regeneration and hypertrophy (miR-1, miR-133a/b, miR-142, miR-128, miR-146b, miR-208b, miR-675), (3) oxidative stress and inflammation (miR-146a/b), and (4) others (miR-149, miR-30c, miR-532-3p, miR-532-5p, miR-542) (Fig. 4).

Identified miRNAs and their involvement in some selected physiological and pathological processes. MiRNAs in blue clouds were RT-qPCR-validated
Functional analysis showed that GE microarray identified genes were significantly associated with the following biological processes: cellular processes, muscle organ development, proteolysis involved in cellular protein catabolic process, muscle cell differentiation, positive regulation of biological processes, cell death, apoptosis, regulation of cell proliferation, and positive regulation of inflammatory process (Additional file 2: Table S2).
Among identified genes (DEG), special attention has been focused on a few important groups which are known to be HMB affected: muscle organ development (e.g., six1, myf5, acta1, cav1, myh3, myh7, myl2, myl3, sgcd, tgfb2), response to wounding/injury (e.g., jak2, igf2, several members of cxcl and interleukin genes, sod1, sod2), inflammatory response/innate immune response/oxidative stress (tlr3, tlr4, tlr10, cd40, cd44, igf2, itgb6, il-5, il-6, il-15, il-23, sod1, sod2, and a large group of chemokine ligand: ccl-1, ccl-2, ccl-5, ccl-8).
RT-qPCR validation
From the microarray results, six miRNAs and six genes were selected as a single representative for the aforementioned processes for further RT-qPCR validation. The analysis confirmed statistically significant differences in the expression of six miRNAs (miR-204, miR-208b, miR-222, miR-675, miR-146a, and miR-146b) and six genes (sod1, sod2, tgfb2, myf5, bdnf, otud4) in HMB-treated ESC when compared to control condition (CTRL) (Fig. 5). All RT-qPCR validated miRNAs and genes presented the same trend as microarray results.

Expression of selected miRNAs (a) and genes (b) validated by RT-qPCR. Presented values are means ± SE (*p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001). HMB—cells treated with HMB and exposed to H2O2. CTRL—cells without HMB treatment, exposed to H2O2 (n = 6)
Prediction and ontological analysis of miRNA target genes (DET)
TargetScan analysis was performed to predict potential target genes for all identified miRNAs. The analysis revealed unique 3310 targets for downregulated and 2117 unique targets for upregulated miRNAs. We compared all identified HMB-regulated DEG and aforementioned predicted miRNA target genes to find those which could be regulated by HMB-induced miRNAs in ESC cultures exposed to H2O2. Finally, 344 differentially expressed target genes (DET) were identified.
Functional analysis showed that DET were associated significantly with several processes which plays an important role in the physiological (protein metabolism, muscle tissue development, cellular homeostasis, apoptosis) and pathological (inflammation, cancer) conditions in muscular tissue (Table 4).
Signaling pathway analysis showed that 27 identified miRNAs could affected target genes involved in several important signaling pathways related to the processes previously described as modified by HMB and also some other which HMB was suspected to affect. The most meaningful pathways are the following: MAPK, RIG-I, Toll-like receptor, hypertrophic cardiomyopathy, ubiquitin-mediated proteolysis, Ras, and response to oxidative stress.
Western blot analysis
Western blot analysis of the level of reference proteins and five proteins related to the muscle tissue, muscle damage, and oxidative stress was performed. However, the results are difficult to interpret. Protein degradation at different levels was observed in all samples treated only with hydrogen peroxide (Fig. 6). In groups pre-incubated with HMB and H2O2, protein degradation was smaller or not observed. It is related to the protein degradation which is strongly linked to hydrogen peroxide effect.

Degradation of protein in H2O2-treated cell cultures
Cell viability, cell damage, and oxidative stress
To measure cell viability, two tests were used—MTT and SYTOX Red Dead Cell (as a component of CellROX Green Reagent kit). In both tests, increased cell viability and decreased amount of dead cells were observed in a group pre-treated with HMB and incubated with H2O2 than in a control group (incubated only with H2O2). All the results from these two tests were statistically significant (p < 0.05). The results of SYTOX Red Dead Cell (A) and MTT test (B) are presented in Fig. 7.

Effect of HMB on cell viability measured by SYTOX Red Cell Dead assay (a) and MTT assay (b). Each value is the mean ± standard error of the results (n = 3, SYTOX Red Cell Dead assay; n = 6, MTT assay). Statistical analysis was performed using the one-way ANOVA (p < 0.05) and unpaired t test (p < 0.05) for SYTOX Red Cell Dead and MTT assay, respectively. H2O2—cells without HMB treatment and exposed to H2O2. H2O2 + HMB—cells treated with HMB and exposed to H2O2
Oxidative stress was measured using CellROX® Green Reagent. There was no significant difference between groups (Fig. 8a). Similar results were obtained with the test for lipid peroxidation. There were no statistically significant differences between HMB pre-treated group and control. However, surprisingly, higher lipid peroxidation trend was observed in a HMB pre-treated group compared to control (Fig. 8b).

Effect of HMB on oxidative stress—CellROX® Green assay (a) and Lipid peroxidation assay (b). Each value is the mean ± standard error of the results (n = 3, CellROX® Green assay; n = 6, lipid peroxidation assay). Statistical analysis was performed using the one-way ANOVA (p < 0.05) and unpaired t test (p < 0.05) for CellROX® Green assay and lipid peroxidation assay, respectively. H2O2—cells without HMB treatment and exposed to H2O2. H2O2 + HMB—cells treated with HMB and exposed to H2O2
Qualitative flow cytometry assay for mitochondrial depolarization (JC-1) showed significant differences between the Q2 population (monomers + aggregates in %) and Q4 population (JC-1) in control and HMB pre-treated group. There was no significant difference between Q1 population (% of aggregates) and Q3 population (% of monomers) (Fig. 9a).

Qualitative flow cytometry assay for mitochondrial depolarization (a) and total antioxidant capacity (TAC) (b). Each value is the mean ± standard error of the results (n = 3, JC-1 assay; n = 6, TAC assay). Statistical analysis was performed using the one-way ANOVA (p < 0.05) and unpaired t test (p < 0.05) for JC-1 assay and TAC assay, respectively. H2O2—cells without HMB treatment and exposed to H2O2. H2O2 + HMB—cells treated with HMB and exposed to H2O2
Results obtained in a total antioxidant capacity (TAC) assay showed significant differences between HMB pre-treated and control group. Higher antioxidant capacity was observed in HMB pre-treated group (Fig. 9b).
Discussion
MicroRNAs are essential regulators for numerous biological processes by modulating gene expression at the post-transcriptional level. Several muscle-specific miRNAs (myomiRs) have been shown to play an important role in normal myoblast proliferation, differentiation, and muscle remodeling in response to different type of factors. Recent studies have begun to link miRNAs and certain muscle-related diseases [21]. Modulation of miRNAs by dietary factors and miRNA-based gene therapies seems to be a promising option for the treatment of cardiac and skeletal muscle diseases [22]. Among dietary additives, HMB seems to be an interesting potential myoprotectant for horses [8]. Previous studies suggest that HMB may be involved in the regeneration processes of skeletal muscles [23]. Moreover, HMB stimulates skeletal muscle satellite cell activation and may potentially increase skeletal muscle regenerative capacity after damage induction [24].
Our objective was to determine the influence of HMB on miRNA and gene expression in differentiating equine satellite cells subjected to damaging activity of hydrogen peroxide, as an in vitro model of short extreme effort-related muscle damage observed in racing and sport horses.
Microarray analysis of total RNA in differentiating ESC incubated with HMB (24 h) and treated with H2O2 (1 h) revealed the difference in the expression of 27 miRNAs (Table 3) and 4740 DEG (Additional file 1: Table S1) from which 344 DET were chosen (Table 4). Identified miRNAs and a large group of identified genes were previously described as these involved in the pathological and physiological processes in skeletal muscles as well as in other tissues. Selected miRNAs (miR-204 (↑), miR-208b (↓), miR-222 (↑), miR-675 (↓), miR-146a (↑), miR-146b (↑)) and genes (bdnf (↓), sod1 (↓), sod2 (↑), tgfb2 (↓), myf5 (↓), otud4 (↑)) were validated by RT-qPCR showing the same trend as in microarray analysis.
HMB effects on miRNAs related to satellite/muscle cell proliferation and differentiation
Of the 27 identified miRNAs, 9 are related to cell proliferation and 13 to differentiation in muscle tissue (Fig. 4). Some of miRNAs seem to be particularly interesting in the context of previous publications confirming proven and potential HMB effect on muscle. Among them, family of miR-146a/b able to balance the induction of muscle proliferation or differentiation with miR-146 up- and downregulation, respectively [25]. The miR-146a was one of the highest differentially expressed molecules showing 120.92 fold change in HMB-treated cells. It could suggest their possible involvement in promotion of HMB-induced myoblast proliferation. It is well-known that activation and proliferation of satellite cells is a prerequisite of skeletal muscle injury repair [12], and it is possible that HMB is capable to influence miRNA expression, increasing myoblast proliferation rate and thus facilitating the myofiber regeneration. Similar observations were done for miR-133, in which upregulation was described as proliferation-inducing while its downregulation was responsible for differentiation progression [26]. Interestingly, miRNA-222/221 which over-expression was noticed in myoblasts undergoing differentiation with its downregulation after differentiation [27] was downregulated in ESC cultures exposed to H2O2 and pretreated with HMB, when compared to control. The same expression trend (↓) was observed in miR-374b which over-expression is known to impair C2C12 cell differentiation, while inhibition promoted this process [28]. Moreover, three miRNAs (miR-675, miR-324, and miR-331) known to be over-expressed in muscle cell differentiation [29, 30] were downregulated in our experiment. Two other miRNAs, miR-206 and miR-1, known to be downregulated in muscle cell proliferation and upregulated during differentiation [31], have manifested downregulation in ESC cultures treated with HMB. Moreover, some of the identified miRNAs showed the opposite trend of expression change to this mentioned above (miR-1↓, miR-133↓, miR-206↓), promoting cell differentiation and proliferation in case of miRNA upregulation and downregulation, respectively. They were represented by miR-204 which was upregulated in differentiated human cardiomyocyte progenitor cells [32] and miR-155 (↑) and miR-193a (↑), known to regulate cell differentiation in muscle cells [33] and brown fat cells [34], respectively. All of them possessed the same expression trend which was observed in our experiment in the case of HMB-treated group.
The search of DET for the aforementioned miRNAs was done using Pathway Studio Web and has revealed a large group of genes involved in proliferation and differentiation, the processes previously described to be HMB modulated. The following cell proliferation-related genes were identified: jak2 (target of identified miR-101, miR-155), rarg (miR-142-3p, miR-30c), pten (miR-146a, miR-374b, miR-193a), ets1 (miR-221/222), and rarb (miR-146a, miR-146b); cell differentiation-related target genes: jak2 (miR-155), pten (miR-1), klf4 (miR-1, miR-146a, miR-206) and ets1 (miR-221/222). Moreover, we identified several target genes which are involved in muscle organ development: sgcd (miR-142-3p), scd (miR-1, miR-128), cav3 (miR-101), tcf12 (miR-101, miR-142-3p, miR-155, miR-204, miR-208, miR-221/222), and col19a1 (miR-1, miR-206), as modulated in ESC treated with HMB. Special attention deserves miR-206 together with described above miR-1 and miR-133 which regulate expression of one of its potential target genes cx43 involved not only in muscle development but also in muscle regeneration where its upregulation was observed [35]. The same expression trend of cx43 was observed in our experiment in HMB-treated group. MiR-206 decreased expression in our experiment may be related to the fact that inhibition of miR-206 robustly increases myotube development [36].
Taken together, changes in expression of pro-proliferative (miR-133a/b, miR-146a/b, miR-222/221) and differentiation-related miRNAs (miR-1, miR-133a/b, miR-155, miR-193a, miR-204, miR-206, miR-221/222, miR-331, miR-324, miR-374, miR-675) were observed following HMB incubation and exposition of ESC cultures to H2O2, with concomitant changes in expression of their corresponding DET. These results, presenting the pro-proliferation and pro-differentiation effects of the aforementioned miRNAs, could be considered as contradictory, but in fact, both processes are important for proper myogenesis—satellite cell proliferation necessary for proper myofiber regeneration manifested by myoblast fusion with damaged fibers or new myofiber formation, here shown at the very early stage of this process.
HMB involvement in oxidative stress and inflammation
In our study, we also observed HMB-related changes in the expression of miRNAs playing an important role in modulation of inflammation and oxidative stress. The acute inflammatory response is protective and stimulates repairing of injured tissue [11, 12]. The inflammatory infiltrate is a component of the satellite cell niche and also a source of locally released cytokines which regulate muscle regeneration.
One of the most interesting miRNAs involved in oxidative stress and inflammation appears to be miR-146 family which members are known as negative regulators of inflammatory cytokine expression during immune response [37, 38]. Curtale et al. [39] showed that miR-146b may mediate anti-inflammatory activities and modulates the TLR4 signaling pathway by direct targeting several genes which are most likely targets for our identified miRNAs (cxcl10, tlr4). Their study also provides evidence for a link between miR-146b and IL-10, indicating that miR-146b induction depends on the activity of IL-10, which is suspected to be realized by muscle cells, both in vivo and in vitro [40]. We did not observe changes in il-10 mRNA expression in our experiment; however, other interleukin and cytokine gene expression was changed (e.g., il-5, il-6, il-13, il-15, il-18, cxcl10, and ccl11).
In muscles, inflammatory response coincides with repair, regeneration, and growth, which involve activation and proliferation of satellite cells, followed by their terminal differentiation. Until now, limited number of data is available to distinguish features of muscle inflammation that promote injury from those that promote growth or repair of muscle. Moreover, dietary supplementation is known to be one of the ways to reduce skeletal and cardiac muscle damage by decreasing the inflammatory and oxidative stress response to exercise in sport horses [41]. In muscles, anti-inflammatory substances (e.g., NSAIDs) are used to control excess local tissue damage by limiting proteolysis from infiltrating inflammatory cells [42, 43]. HMB has been suggested to inhibit inflammation [44]. However, its anti-inflammatory mechanism is still not fully understood. Recent study conducted by Yakabe et al. [44] suggested that HMB has anti-inflammatory potential by downregulation of IL-6 expression. Surprisingly, il-6 was upregulated in our experiment (FC = 20.01). Interestingly, the local production of IL-6 by skeletal muscle cells and stromal cells promotes activation of satellite cells, thereby increasing myotube regeneration [45]. It is known that IL-6 mediates many aspects of the exercise-induced acute-phase response, including the upregulation of antioxidant defenses as response to oxidative stress [46]. Similar to the aforementioned authors who demonstrated that miR-146b may inhibit pro-inflammatory cytokine secretion, we observed over-expression of both miR-146a and miR-146b. Moreover, miR-155, known to be an immunomodulatory miRNA, acts as a broad limiter of pro-inflammatory gene expression in muscle [47] possessing the same trend which was observed in our experiment in the case of HMB-treated group. This, in turn, suggests that HMB may play an important role in the inflammation processes as an anti-inflammatory factor which could be related to the inhibition of pro-inflammatory cytokine secretion by HMB-induced miR-146 over-expression and activate innate immunity response by miR-155 over-expression.
Interestingly, among DEG (several of them were classified as DET), a large group with the highest FC was involved in different kind of processes related to immunity-acute phase of inflammatory, activation of immunity cells, innate immunity, and pro-inflammatory activity (Table 5). Several of them (e.g., ccl11, ccl2, cxcl10, and saa1) were strongly upregulated, and this tendency was previously described as associated with pro-inflammatory activity status [48] which is not fully consistent with tendency of identified in our experiment miRNA expression. Moreover, that a large group of DEG is involved in the inflammatory response and innate immune response in different kind of tissues (tlr3, tlr4, tlr10, cd40, cd44, igf2, itgb6, il-5, il-6, il-15, il-23, and a group of chemokine ligand: ccl-1, ccl-2, ccl-5, ccl-8). Both processes are necessary during regeneration, upon injury when immune cells rapidly infiltrate the muscle tissue to remove injured necrotic cells and secrete factors that are essential to activate satellite cells.
The search of target genes for the identified aforementioned miRNAs has revealed a number of genes involved in innate immunity and processes accompanying inflammation, which HMB affects, represented by jak2 (miR-101, miR-155), tlr4 (miR-146a/b, miR-155), cxcl10, cxcl11 (miR-146a/b), and cd47-target for oxidative stress and inflammation-related miRNAs (miR-221/222 and miR-155). HMB impact on the inflammatory processes and oxidative stress is not fully understood. However, our results show that this substance can modulate in the opposite way expression of pro- and anti-inflammatory miRNAs and genes. We assume this could be linked not only to the potential anti-inflammatory effect of HMB but also to the activation of early (innate) immune response (associated with H2O2-related damage), which is the initial phase of the regeneration process.
MicroRNAs related to cell reaction to injury—potential role of HMB as myoprotectant
Among all identified miRNAs, several were known to be involved in cell reaction to injury and different phases of regeneration.
MiR-675 seems to be one of the most interesting miRNAs which is the second of the most downregulated in HMB-treated group and is closely related to regeneration processes. Previous studies showed that miR-675 is expressed in skeletal muscles during myoblast differentiation and muscle regeneration [29]. Another miRNA, miR-146, is related to satellite cell activation [49], myoblast differentiation, and muscle regeneration in vivo [50]. Moreover, also, miR-208 is known to be involved in injury-induced satellite cell activation [51]. We suspect that HMB may stimulate and/or accelerate activation of equine satellite cells at very early stage of the regeneration process. These observations may suggest that HMB acting by the aforementioned miRNA induction may be involved in the satellite cell activation which accompanies regeneration. Hypertrophy is also an important phenomenon of regeneration process in muscle; however, it is related to the final stage of regeneration [13].
We identified several miRNAs that were previously described in relation to muscle hypertrophy. They were represented by downregulation in skeletal muscle hypertrophy miR-1 and miR-133a/b [52, 53] which possessed the same trend as was noticed in our experiment in the case of HMB-treated differentiating ESC cultures. Similar observation was done for miR-142 (↓), which as mentioned above presented one of the highest fold changes (FC = 105.23) among the identified miRNAs, and its downregulation was described during cardiac hypertrophy and is able to inhibit cytokine signaling and function in the myocardium [53]. It is possible that HMB incubation changes the expression of the aforementioned miRNAs facilitating for more efficient late regeneration (not observed during the first day after injury). More research is needed to evaluate the role of miR-133a/miR-1/miR-142-dependent hypertrophy mechanisms of HMB action in the activation of skeletal hypertrophy and muscle regeneration in different physiological and pathological conditions.
Several potential target genes for identified miRNAs were classified as follow: hypertrophy: tpm3 (miR-1, miR-206), tpm1 (miR-142-5p), sgcd (miR-142-5p), cacnb1 (miR-208b), itgav (miR-142-5p), itga2 (miR-30c, miR-128); muscle regeneration-related processes and pathways: cx43 (miR-206), klf4 (miR-30c), and vegfc (miR-128, miR-133).
Other HMB-related miRNAs and genes
The effect of several identified in our experiment miRNAs in muscle tissue is still not clear. However, our results suggest that HMB modulates miRNA expression which was previously described in connection with various physiological and pathological conditions and could also be affected by HMB in injured muscle tissue: miR-374b—related to lipid and protein metabolism [54], miR-14—regulator of mitochondria function in C2C12 [55, 56], miR-532-3p—regulates mitochondrial fission in cardiac muscle [57], miR-30c—cellular adipogenesis and cardiac hypertrophy and ischemia [58], miR-450a/c and miR-142—negative regulators of cardiac hypertrophy [59], and miR-542 family—regeneration of various tissues [60]. Interestingly, several identified genes (among them potential target genes for the aforementioned miRNAs) are also related to amino acid metabolism and protein ubiquitination and proteasome pathways (rnf6, cand2, ube2b, rnf138, rnf19b, rnf38, ube2l6, birc3, rnf114, mib1, trim36, and rnf4) which are closely related to the protein degradation during muscle atrophy in cachexia, and HMB is known to inhibit this process in humans and animals [2, 5, 6].
Target gene-related signaling pathways
Furthermore, ontological analysis of miRNA DET revealed that a large group of genes was associated with biological processes and signaling pathways that are closely related to the muscle cell, muscle tissue development, and protein metabolism presented in Tables 6 and 7, respectively.
Among the most strongly regulated pathways were these previously described as engaged in immunity and inflammation: Toll-like receptor, RIG-I, Ras, MAPK, and ubiquitin-mediated proteolysis. Toll-like receptor pathway is related to the inflammatory activity [61] and different kind of myopathies and could be modulated by miR-155 [62]. RIG-I-like receptor pathway is known to play a crucial role in innate response [63] which is necessary to activate early muscle regeneration and may be modulated by three identified miRNAs: miR-146a, miR-146b, and miR-155. Ras and MAPK pathways promote protein degradation in muscle cells [64] and oxidative stress (closely related to miR-146 activity) [61]. Moreover, MAPK pathway plays a pivotal role in the energy metabolism through modulating lipid metabolism, skeletal muscle growth, and different kind of muscular myopathies and atrophy [65]. Finally, ubiquitin-mediated proteolysis identified as a potential target gene-related pathway is closely related to muscle atrophy by protein degradation.
Western blot analysis—HMB protective effect on proteins
Western blot analysis was performed to check the level of proteins corresponding with several genes and miRNA. However, the results that were obtained are impossible to use in this way. It is related to the protein degradation which is strongly linked to hydrogen peroxide effect. Previous studies related to hydrogen peroxide effect on different kind of proteins showed that this substance cause protein degradation. For example, exposure of skeletal muscle myotubes to ROS (i.e., hydrogen peroxide) can activate proteases leading to protein degradation as it was described by Li et al. [21] and Clung et al. [55]. Finnegan et al. [9] observed the interactions of a liquid and gaseous H2O2 with amino acids and proteins (bovine serum albumin and aldolase). In this study, authors observed that two dosages of hydrogen peroxide cause total degradation of BSA (the absence of bands). In our study, we observed similar situation. We checked three different housekeeping genes: β-actin, gapdh, and α-tubulin. For all of them, we noticed the same trend of protein degradation in samples where the cells were exposed to hydrogen peroxide (Fig. 6). Moreover, we checked several different proteins where we also noticed protein degradation on a similar level as in samples mentioned above. We suspect that samples where the cells were previously incubated with HMB were protected from protein degradation related to hydrogen peroxide. This degradation has already occurred at the cellular level, not during the sample preparation for western blotting. During the sample preparation procedure, we used RIPA buffer with protease inhibitor cocktail. Based on this observation and previous studies [66,67,68] related to HMB, we think that this substance may decrease protein degradation in equine satellite cells exposed to hydrogen peroxide; however, the comparison of protein levels between experimental and control conditions is impossible.
Cell viability, cell damage, and oxidative stress
Five different tests were performed to check the effect of potential antioxidant properties of HMB and its cell protection activity. Based on our results from the test where we observed direct cell oxidative stress together with typical damage (lipid peroxidation together with mitochondria activity changes) related to oxidative stress, it cannot be unambiguously determined whether HMB in experimental dosage has properties that limit oxidative stress (Figs. 8a, b and 9a). However, we observed that HMB increase total antioxidant capacity (Fig. 9b). According to our knowledge, there are limited data considering direct antioxidant activity of HMB; however, as it was mentioned, our study revealed several miRNAs, genes, and pathways related to oxidative stress and antioxidant activity which were modulated by HMB. Increased total antioxidant capacity may represent an adaptive response to the enhanced generation of ROS related to hydrogen peroxide activity or likely responsible for the attenuation of oxidative damage.
Two independent tests showed that HMB increased cell viability: MTT and SYTOX Red Dead Cell (Fig. 7). These observations are consistent with the study presented by Vallejo et al. [69]. Authors also observed that HMB enhanced myoblast viability (in C2C12 cell line). Moreover, earlier study by Pacini et al. [70] showed that HMB significantly prevented dexamethasone-induced cell mortality. Based on our results, we think that HMB may prevents cells from the hydrogen peroxide-induced mortality and enhance ESC’s viability.
Conclusions
Our study presents several new findings of the mechanisms of action of HMB and its potential role in muscle physiology and pathology. We focused on HMB-induced miRNA expression changes previously described as those associated with the muscle tissue injury (inflammation), regeneration, and the accompanying processes such as cell activation, proliferation, migration, and differentiation. The scheme of putative mode of miRNAs and DEG-depended HMB action is proposed in Fig. 10.

Potential role of HMB-induced miRNAs and selected target genes in muscle regeneration process
We demonstrated for the first time that HMB-treated equine satellite cells exposed to H2O2 have modulated expression of 27 miRNAs which could affect the abovementioned processes. Many of them were not known for being differentially expressed during myogenic proliferation, differentiation, or processes related to muscle injury and activation in early stage of regeneration. That is why, it would also be interesting to investigate whether some of the abovementioned miRNAs participate in skeletal muscle degeneration/regeneration process as well as degeneration-related equine muscular diseases, such as muscle inflammation related to the extreme effort or recurrent rhabdomyopathy.
Moreover, we found DET for identified HMB-modulated miRNAs which are related to key processes in muscle physiology and pathology. Also, identified pathways MAPK, Ras, and RIG-I together with those involved in oxidative stress response seem to support our knowledge about the potential mechanisms of HMB action. Based on the obtained results, we also believe that HMB increases the survival of cells treated with hydrogen peroxide. Further analyses evaluating the effect of HMB on injured, recovering muscle tissue are needed to verify the collected data.
Abbreviations
- CTRL:
-
Control condition
- DEG:
-
Differentially expressed genes
- DET:
-
Differentially expressed target genes
- DM:
-
Differentiation medium
- DMSO:
-
Dimethylsulfoxide
- ESCs:
-
Equine satellite cells
- FBS:
-
Fetal bovine serum
- GE:
-
Gene expression
- GM:
-
Growth medium
- HMB:
-
β-Hydroxy-β-methylbutyrate
- HS:
-
Horse serum
- IB:
-
Incubation buffer
- PBS:
-
Phosphate-buffered saline
- ROS:
-
Reactive oxygen species
References
Van Koevering MT, Dolezal HG, Gill DR, Owens FN, Strasia CA, Buchanan DS, Lake R, Nissen S. Effects of beta-hydroxy-beta-methyl butyrate on performance and carcass quality of feedlot steers. J Anim Sci. 1994;72:1927–35.
Aversa Z, Bonetto A, Costelli P, Minero VG, Penna F, Baccino FM, Lucia S, Rossi Fanelli F, Muscaritoli M. ß-Hydroxy-ß-methylbutyrate (HMB) attenuates muscle and body weight loss in experimental cancer cachexia. Int J Oncol. 2011;38:713–20.
Szcześniak KA, Ostaszewski P, Fuller JC, Ciecierska A, Sadkowski T. Dietary supplementation of β-hydroxy-β-methylbutyrate in animals—a review. J Anim Physiol Anim Nutr. 2015;99(3):405–17. https://doi.org/10.1111/jpn.12234.
Norton LE, Layman DK. Leucine regulates translation initiation of protein synthesis in skeletal muscle after exercise. J Nutr. 2006;136:533S–7S.
Panton LB, Rathmacher JA, Baier S, Nissen S. Nutritional supplementation of the leucine metabolite β-hydroxy-β-methylbutyrate (HMB) during resistance training. Nutrition. 2000;16:734–9.
Deutz NE, Pereira SL, Hays NP, Oliver JS, Edens NK, Evans CM, Wolfe RR. Effect of β-hydroxy-β-methylbutyrate (HMB) on lean body mass during 10 days of bed rest in older adults. Clin Nutr. 2013;32:704–12.
Van Someren KA, Edwards AJ, Howatson G. Supplementation with [beta]-hydroxy-[beta]-methylbutyrate (HMB) and [alpha]-ketoisocaproic acid (KIC) reduces signs and symptoms of exercise-induced muscle damage in man. Int J Sport Nutr Exerc Metab. 2005;15:413–24.
Ostaszewski P, Kowalska A, Szarska E, Szpotański P, Cywinska A, Bałasińska B, Sadkowski T. Effects of β-hydroxy-β-methylbutyrate and γ-oryzanol on blood biochemical markers in exercising thoroughbred race horses. J Equine Vet Sci. 2012;32:542–51.
Finnegan M, Linley E, Denyer SP, McDonnell G, Simons C, Maillard JY. Mode of action of hydrogen peroxide and other oxidizing agents: differences between liquid and gas forms. J Antimicrob Chemother. 2010;65(10):2108–15.
Hirai DM, Copp SW, Schwagerl PJ, Musch TI, Poole DC. Acute effects of hydrogen peroxide on skeletal muscle microvascular oxygenation from rest to contractions. J Appl Physiol. 2011;110:1290–8.
Aoi W, Naito Y, Takanami Y, Kawai Y, Sakuma K, Ichikawa H, Yoshida N, Yoshikawa T. Oxidative stress and delayed-onset muscle damage after exercise. Free Radic Biol Med. 2004;37:480–7.
Ciecierska A, Chodkowska K, Motyl T, Sadkowski T. Myogenic cells applications in regeneration of post-infarction cardiac tissue. J Physiol Pharmacol. 2013;64:401–8.
Van der Meer SF, Jaspers RT, Jones DA, Degens H. The time course of myonuclear accretion during hypertrophy in young adult and older rat plantaris muscle. Ann Anat. 2011;193(1):56–63.
Kovanda A, Režen T, Rogelj B. MicroRNA in skeletal muscle development, growth, atrophy, and disease. Wiley Interdiscip Rev RNA. 2014;5(4):509–25.
Szcześniak KA, Ciecierska A, Ostaszewski P, Sadkowski T. Characterisation of equine satellite cell transcriptomic profile response to β-hydroxy-β-methylbutyrate (HMB). Br J Nutr. 2016;116(8):1315.
Mossman T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods 1983;65:55.
Ostaszewski P, Kostiuk S, Balasińska B, Jank M, Papet I, Glomot F. The leucine metabolite 3-hydroxy-3-methylbutyrate (HMB) modifies protein turnover in muscles of laboratory rats and domestic chickens in vitro. J Anim Physiol Anim Nutr. 2000;84:1–8.
Szcześniak KA, Ciecierska A, Ostaszewski P, Sadkowski T. Transcriptomic profile adaptations following exposure of equine satellite cells to nutriactive phytochemical gamma-oryzanol. Genes Nutr 2016;11(1):1.
Grimson A, Farh KK, Johnston WK, Garrett-Engele P, Lim LP, Bartel DP. MicroRNA targeting specificity in mammals: determinants beyond seed pairing. Mol Cell. 2007;27(1):91–105.
Zielniok K, Motyl T, Gajewska M. Functional interactions between 17β-estradiol and progesterone regulate autophagy during acini formation by bovine mammary epithelial cells in 3D cultures. Biomed Res Int. 2014;2014. Article ID 382653. https://doi.org/10.1155/2014/382653.
Li YP, Chen Y, Li AS, Reid MB. Hydrogen peroxide stimulates ubiquitin-conjugating activity and expression of genes for specific E2 and E3 proteins in skeletal muscle myotubes. Am J Physiol Cell Physiol. 2003;285(4):C806–12.
Pasiakos SM, McClung JP. MiRNA analysis for the assessment of exercise and amino acid effects on human skeletal muscle. Adv Nutr. 2013;4(4):412–7.
Jowko E, Ostaszewski P, Jank M, Sacharuk J, Zieniewicz A, Wilczak J, Nissen S. Creatine and beta-hydroxy-beta-methylbutyrate (HMB) additively increase lean body mass and muscle strength during a weight-training program. Nutrition. 2001;17:558–66.
Kornasio R, Riederer I, Butler-Browne G, Mouly V, Uni Z, Halevy O. Beta-hydroxy-beta-methylbutyrate (HMB) stimulates myogenic cell proliferation, differentiation and survival via the MAPK/ERK and PI3K/Akt pathway. Biochim Biophys Acta. 2009;1793:755–63.
Kuang W, Tan J, Duan Y, Duan J, Wang W, Jin F, Jin Z, Yuan X, Liu Y. Cyclic stretch induced miR-146a upregulation delays C2C12 myogenic differentiation through inhibition of Numb. Biochem Biophys Res Commun. 2009;378:259–63.
Chen JF, Mandel EM, Thomson JM, Wu Q, Callis TE, Hammond SM, Conlon FL, Wang DZ. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat Genet. 2006;38:228–33.
Cardinali B, Castellani L, Fasanaro P, Basso A, Alemà S, Martelli F, Falcone G. Microrna-221 and microrna-222 modulate differentiation and maturation of skeletal muscle cells. PLoS One. 2009;4(10):e7607. https://doi.org/10.1371/journal.pone.0007607.
Motohashi N, Alexander MS, Shimizu-Motohashi Y, Myers JA, Kawahara G, Kunkel LM. Regulation of IRS1/Akt insulin signaling by microRNA-128a during myogenesis. J Cell Sci. 2013;126:2678–91.
Dey BK, Pfeifer K, Dutta A. The H19 long noncoding RNA gives rise to microRNAs miR-675-3p and miR-675-5p to promote skeletal muscle differentiation and regeneration. Genes Dev. 2014;28(5):491–501.
Dmitriev P, Barat A, Polesskaya A, O’Connell MJ, Robert T, Dessen P, Walsh TA, Lazar V, Turki A, Carnac G, et al. Simultaneous miRNA and mRNA transcriptome profiling of human myoblasts reveals a novel set of myogenic differentiation-associated miRNAs and their target genes. BMC Genomics. 2013;14:265.
Chen JF, Tao Y, Li J, Deng Z, Yan Z, Xiao X, Wang DZ. MicroRNA-1 and microRNA-206 regulate skeletal muscle satellite cell proliferation and differentiation by repressing Pax7. J Cell Biol. 2010;190(5):867–79.
Xiao J, Liang D, Zhang H, Liu Y, Zhang D, Liu Y, Pan L, Chen X, Doevendans PA, Sun Y, et al. MicroRNA-204 is required for differentiation of human-derived cardiomyocyte progenitor cells. J Mol Cell Cardiol. 2012;53:751–9.
Seok HY, Tatsuguchi M, Callis TE, He A, Pu WT, Wang DZ. MiR-155 inhibits expression of the MEF2A protein to repress skeletal muscle differentiation. J Biol Chem. 2011;286:35339–46.
Sun L, Xie H, Mori MA, Alexander R, Yuan B, Hattangadi SM, Lodish HF. Mir193b-365 is essential for brown fat differentiation. Nat Cell Biol. 2011;13:958–65.
Araya R, Eckardt D, Maxeiner S, Krüger O, Theis M, Willecke K, Sáez JC. Expression of connexins during differentiation and regeneration of skeletal muscle: functional relevance of connexin43. J Cell Sci. 2005;118(1):27–37.
Winbanks CE, Beyer C, Hagg A, Qian H, Sepulveda PV, Gregorevic P. miR-206 represses hypertrophy of myogenic cells but not muscle fibers via inhibition of HDAC4. PLoS One. 2013;8:e73589.
Bhaumik DL, Scott GK, Schokrpur S, Patil CK, Orjalo AV, Rodier F, Lithgow GJ, Campisi J. MicroRNAs miR-146a/b negatively modulate the senescence-associated inflammatory mediators IL-6 and IL-8. Aging. 2009;1:402.
Taganov KD, Boldin MP, Chang KJ, Baltimore D. NF-κB-dependent induction of microRNA miR-146, an inhibitor targeted to signalling proteins of innate immune responses. Proc Natl Acad Sci. 2006;103:12481–6.
Curtale G, Mirolo M, Renzi TA, Rossato M, Bazzoni F, Locati M. Negative regulation of Toll-like receptor 4 signaling by IL-10-dependent microRNA-146b. Proc Natl Acad Sci U S A. 2013;110(28):11499–504.
Divangahi M, Demoule A, Danialou G, Yahiaoui L, Bao W, Xing Z, Petrof BJ. Impact of IL-10 on diaphragmatic cytokine expression and contractility during Pseudomonas infection. Am J Respir Cell Mol Biol. 2007;36(4):504–12.
Ducharme NG, Fortier LA, Kraus MS, Hobo S, Mohammed HO, McHugh MP, Hackett RP, Soderholm LV, Mitchell LM. Effect of a tart cherry juice blend on exercise-induced muscle damage in horses. Am J Vet Res. 2009;70(6):758–63.
Liu N, Bassel-Duby R. Regulation of skeletal muscle development and disease by microRNAs. Results Probl Cell Differ. 2015;56:165–90.
Mehallo CJ, Drezner JA, Bytomski JR. Practical management: nonsteroidal antiinflammatory drug (NSAID) use in athletic injuries. Clin J Sport Med. 2006;16(2):170–4.
Yakabe M, Ogawa S, Ota H, Iijima K, Eto M, Ouchi Y, Akishita M. Beta-hydroxy-beta-methylbutyrate inhibits lipopolysaccharide-induced Interleukin-6 expression by increasing protein phosphatase-1α expression. RNA Transcription. 2015:1–5. https://doi.org/10.11648/j.rnat.20150101.11.
Steensberg A, Febbraio MA, Osada T, Schjerling P, Hall G, Saltin B, Pedersen BK. Interleukin-6 production in contracting human skeletal muscle is influenced by pre-exercise muscle glycogen content. J Physiol. 2001;537(2):633–9.
Sacheck JM, Cannon JG, Hamada K, Vannier E, Blumberg JB, Roubenoff R. Age-related loss of associations between acute exercise-induced IL-6 and oxidative stress. Am J Physiol Endocrinol Metab. 2006;291(2):E340–9.
Nie M, Liu J, Yang Q, Seok HY, Hu X, Deng ZL, Wang DZ. MicroRNA-155 facilitates skeletal muscle regeneration by balancing pro-and anti-inflammatory macrophages. Cell Death Dis. 2016;7(6):e2261.
Zhang JM, An J. Cytokines, inflammation and pain. Int Anesthesiol Clin. 2007;45(2):27.
Cheung TH, Quach NL, Charville GW, Liu L, Park L, Edalati A, Rando TA. Maintenance of muscle stem-cell quiescence by microRNA-489. Nature. 2012;482(7386):524–8.
Khann N, Ge Y, Chen J. MicroRNA-146b promotes myogenic differentiation and modulates multiple gene targets in muscle cells. PLoS One. 2014;9(6):e100657.
Ji X, Takahashi R, Hiura Y, Hirokawa G, Fukushima Y, Iwai N. Plasma miR-208 as a biomarker of myocardial injury. Clin Chem. 2009;55:1944–9.
Carè A, Catalucci D, Felicetti F, Bonci D, Addario A, Gallo P, Bang ML, Segnalini P, Gu Y, Dalton ND, et al. MicroRNA-133 controls cardiac hypertrophy. Nature med. 2007;13:613–8.
Sharma S, Liu J, Wei J, Yuan H, Zhang T, Bishopric NH. Repression of miR-142 by p300 and MAPK is required for survival signalling via gp130 during adaptive hypertrophy. EMBO Mol Med. 2012;4(7):617–32.
Pan S, Zheng Y, Zhao R, Yang X. MicroRNA-130b and microRNA-374b mediate the effect of maternal dietary protein on offspring lipid metabolism in Meishan pigs. Br J Nutr. 2013;109(10):1731–8.
McClung JM, Judge AR, Talbert EE, Powers SK. Calpain-1 is required for hydrogen peroxide-induced myotube atrophy. Am J Physiol Cell Physiol. 2009;296(2):C363–71.
Mohamed JS, Hajira A, Pardo PS, Boriek AM. MicroRNA-149 inhibits PARP-2 and promotes mitochondrial biogenesis via SIRT-1/PGC-1α network in skeletal muscle. Diabetes. 2014;63(5):1546–59.
Wang JX, Zhang XJ, Feng C, Sun T, Wang K, Wang Y. MicroRNA-532-3p regulates mitochondrial fission through targeting apoptosis repressor with caspase recruitment domain in doxorubicin cardiotoxicity. Cell Death Dis. 2015;6(3):e1677.
Irani S, Hussain MM. Role of microRNA-30c in lipid metabolism, adipogenesis, cardiac remodeling and cancer. Curr Opin Lipidol. 2015;26(2):139–46.
Jeong MH, Lee JS, Park WJ, Yang DK. Identification of novel microRNAs negatively regulating cardiac hypertrophy. Biochem Biophys Res Commun. 2012;428(1):191–6.
Kureel J, Dixit M, Tyagi AM, Mansoori MN, Srivastava K, Raghuvanshi A, Singh D. miR-542-3p suppresses osteoblast cell proliferation and differentiation, targets BMP-7 signaling and inhibits bone formation. Cell Death Dis. 2014;5(2):e1050.
Pandur S, Pankiv S, Johannessen M, Moens U, Huseby NE. γ-Glutamyltransferase is upregulated after oxidative stress through the Ras signal transduction pathway in rat colon carcinoma cells. Free Radic Res. 2007;41(12):1376–84.
Tournadre A, Lenief V, Miossec P. Expression of toll-like receptor 3 and toll-like receptor 7 in muscle is characteristic of inflammatory myopathy and is differentially regulated by Th1 and Th17 cytokines. Arthritis Rheum. 2010;62(7):2144–51.
Kawai T, Akira S. Toll-like receptor and RIG-1-like receptor signaling. Ann N Y Acad Sci. 2008;1143(1):1–20.
Szewczyk NJ, Peterson BK, Jacobson LA. Activation of Ras and the mitogen-activated protein kinase pathway promotes protein degradation in muscle cells of Caenorhabditis elegans. Mol Cell Biol. 2002;
Geisler HW, Shi H, Gerrard DE. MAPK pathway in skeletal muscle diseases. J Vet Sci Anim Husb. 2013;1(1):e104. https://doi.org/10.15744/2348-9790.1.e1004.
Girón MD, Vílchez JD, Shreeram S, Salto R, Manzano M, Cabrera E, López-Pedrosa JM. β-Hydroxy-β-methylbutyrate (HMB) normalizes dexamethasone-induced autophagy-lysosomal pathway in skeletal muscle. PLoS One. 2015;10(2):e0117520. 22(12):4181–88
Noh KK, Chung KW, Choi YJ, Park MH, Jang EJ, Park CH, Chung HY. β-Hydroxy β-methylbutyrate improves dexamethasone-induced muscle atrophy by modulating the muscle degradation pathway in SD rat. PLoS One. 2014;9(7):e102947.
Smith HJ, Wyke SM, Tisdale MJ. Mechanism of the attenuation of proteolysis-inducing factor stimulated protein degradation in muscle by β-hydroxy-β-methylbutyrate. Cancer Res. 2004;64(23):8731–5.
Vallejo J, Spence M, Cheng AL, Brotto L, Edens NK, Garvey SM, Brotto M. Cellular and physiological effects of dietary supplementation with β-hydroxy-β-methylbutyrate (HMB) and β-alanine in late middle-aged mice. PLoS One. 2016;11(3):e0150066.
Pacini A, Mannelli LDC, Tenci B, Maresca M, Celi C, Gabriele R, Ghelardini C. Therapeutic option by synergistic administration of the β-hydroxy-β-methyl butyrate and R (+) lipoic acid in a cellular model of dexamethasone-dependent sarcopenia. Ital J Anat Embryol. 2015;120(1):152.
Acknowledgements
The authors would like to thank Dr. Małgorzata Gajewska and Emmanuel Gonzalez Escobar for their help in preparing the manuscript.
Funding
This research was funded by National Science Centre (Poland), Grant No. 2011/03/B/NZ5/05697. Publication of this manuscript was supported by KNOW (Leading National Research Centre) Scientific Consortium “Healthy Animal - Safe Food,” decision of the Ministry of Science and Higher Education No. 05-1/KNOW2/2015.
Availability of data and materials
All data generated or analyzed during this study are included in this published article (and its additional files).
Author information
Authors and Affiliations
Contributions
KCH carried out the muscle sampling, cell viability, oxidative stress and lipid peroxidation assays, RT-qPCR validation of microarray results, western blot, ontological analysis, and interpretation of the obtained data and wrote the manuscript. AC carried out the equine satellite cell isolation and culture, RNA isolation and microarray analysis, and western blot. KM carried out the flow cytometry analysis and data interpretation. PO participated in the study design and helped in the manuscript revision. TS participated in the study design; supervised the project; performed the muscle sampling, statistical analyses of microarray and RT-qPCR data, cell viability, oxidative stress, and lipid peroxidation; assisted in the manuscript preparation and revision; and prepared the figures. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study complies with the national and institutional guidelines of the use of animals in research according to the Polish legal act from January 21, 2005. Since sample collection was performed during the routine slaughter and no additional procedures that would be harmful and painful for animals were applied, this study did not require formal ethics approval.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1:
Table S1. Genes differentially expressed in HMB-incubated equine satellite cells exposed to H2O2, compared to control. FDR ≤ 0.05, FC ≥ 1.3, n = 4. (XLSX 550 kb)
Additional file 2:
Table S2. Biological function of differentially expressed genes (DEG). (XLSX 28 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Chodkowska, K.A., Ciecierska, A., Majchrzak, K. et al. Effect of β-hydroxy-β-methylbutyrate on miRNA expression in differentiating equine satellite cells exposed to hydrogen peroxide. Genes Nutr 13, 10 (2018). https://doi.org/10.1186/s12263-018-0598-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12263-018-0598-2