
Abstract
Background
p57Kip2, a cyclin-dependent kinase inhibitor, is considered to be a candidate tumor suppressor gene that has been implicated in Beckwith-Wiedemann syndrome and sporadic cancers. In addition, decreased expression of p57Kip2 protein has been frequently observed in pancreatic, lung, breast, bladder, gastrointestinal tract and prostate cancers. However, p57Kip2 gene mutations are rare in these cancers suggesting that other unknown mechanisms might be at play in reducing its expression. The aim of this study was to investigate the molecular mechanism of down-regulation of p57Kip2 in prostate cancer.
Findings
We observed a significant negative correlation between the expression of p57Kip2 and microRNA-21 (miR-21) in prostate cancer samples and after androgen deprivation with castration in the CWR22 human prostate cancer xenograft model. We report that miR-21 targeted the coding region and decreased p57Kip2 mRNA and protein levels in prostate cancer cells. Conversely, inhibition of endogenous miR-21 by an anti-miR-21 inhibitor strongly induced p57Kip2 expression. Furthermore, we found that knockdown of p57Kip2 reversed the effects of the anti-miR-21 inhibitor on cell migration and anchorage-independent cell growth.
Conclusions
Our results indicate that miR-21 is able to downregulate p57Kip2 expression by targeting the coding region of the gene and is also able to attenuate p57Kip2 mediated functional responses. This is the first report demonstrating that p57Kip2 is a novel target of miR-21 in prostate cancer and revealing a novel oncogenic function of this microRNA.
Similar content being viewed by others
Introduction
Uncontrolled cell proliferation due to aberrant regulation of cell cycle control can lead to the development of cancer. Cyclin dependent kinase inhibitors (CKIs) are the common inhibitors of cell cycle which consists of two families: INK4 family (consisting of p16INK4a, p15INK4b, p18INK4c and p19INK4d) and Cip/Kip family (p21CIP1/WAF1, p27Kip2 and p57Kip2) [1]. Human p57Kip2 gene is maternally expressed and paternally imprinted and is located on chromosome 11p15.5 [2], which is implicated in Beckwith-Wiedemann syndrome [3] and in sporadic cancers. p57Kip2 is also required for normal development as p57Kip2 null mice die at 2 weeks of age and show increased apoptosis and delayed differentiation during mouse development [4]. Because of its chromosomal location, imprinting status and functional activities, p57Kip2 is considered to be a candidate tumor suppressor gene. p57Kip2 overexpression in LNCaP prostate cancer cells resulted in the conversion of adenocarcinoma to a more differentiated squamous tumor in nude mice, with reduced cell proliferation and tumor invasion [5]. p57Kip2 is considered to be a tumor suppressor gene since it functions to block cell proliferation by inhibiting cell cycle progression, promotes apoptosis and cell differentiation, inhibits tissue invasion and metastasis and also inhibits angiogenesis [6]. Hence, cancer cells frequently down-regulate p57Kip2 in order to gain a proliferative advantage.
Loss of or reduced p57Kip2 expression occurs in carcinomas of the prostate, bladder, liver, pancreas, breast and others. The absence of p57Kip2 gene mutations in a wide variety of cancers suggests that other transcriptional or post-transcriptional mechanisms might be involved in its down-regulated protein expression. Inactivation of p57Kip2 gene due to promoter DNA methylation was observed in non-small cell lung cancer and in lymphoid malignancies of B-cells [7, 8]. Yang et al. found that p57Kip2 expression in breast cancer cells was repressed due to Polycomb protein EZH2-mediated H3K27me3 chromatin mark. Increased p57Kip2 degradation due to ubiquitylation by E3 ligase Skp1/ Cul1/ F-box (SCF complex) was observed in non-small cell lung carcinoma and hepatocellular carcinoma [9, 10].
Mature miRNAs are ~22 nucleotides long non-coding single-stranded RNAs, which upon binding to the 3’-UTR region of target mRNAs can result in mRNA cleavage, or translational repression. p57Kip2 has been reported to be targeted by miR-221/222 cluster in gastric carcinoma, ovarian cancer and hepatocellular carcinoma [11]. miR-92b and miR-25 have also been reported to down-regulate p57Kip2 expression in human embryonic stem cells and in gastric cancer respectively [12, 13]. Jin et al. demonstrated that expression of p57Kip2 is significantly decreased in human prostate cancer and overexpression of p57Kip2 in prostate cancer cells decreased cell proliferation and reduced invasiveness [5]. However, the mechanism behind p57Kip2 down-regulation in prostate cancer cells has not been investigated. Unexpectedly, we discovered that p57Kip2 is one of the novel downstream target genes of miR-21 in prostate cancer. We observed a very strong negative correlation between p57Kip2 and miR-21 expression in human prostate tumor samples and in CWR22, a human prostate cancer xenograft model. Our results for the first time show that miR-21 can down-regulate p57Kip2 mRNA and protein expression by targeting its coding region to attenuate its activity in prostate cancer cells. Thus, therapeutic approaches aimed at restoring p57Kip2 expression might be beneficial for prostate cancer prevention and therapy.
Findings and discussion
MicroRNA-21 targets p57Kip2 gene
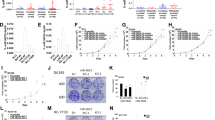
Inactivation of p57Kip2 is commonly observed in cancers. MicroRNAs offer another layer of complexity to the regulation of its gene expression. Recently, microRNAs (miR-221/222, miR-25, miR-92b) were reported to downregulate the expression of p57Kip2 transcript [13, 14]. In our studies, we found a very significant p57Kip2 downregulation in human prostate tumor samples compared to the normal prostate cells in consistence with the published studies [5, 15]. However, the mechanism of p57Kip2 repression has not been well studied in prostate cancer. We along with others have shown a significant upregulation in miR-21 expression in prostate cancer [16, 17]. Surprisingly, we found a significant (P < 0.05) inverse correlation between miR-21 and p57Kip2 expression in human prostate cancer samples downloaded from the Cancer Genome Atlas (TCGA) database (Figure 1A). We also checked miR-21 expression in different prostate cancer cells and found that all the cells expressed miR-21 albeit at different levels. Cells with relatively higher miR-21 levels such as MDA-PCa-2b and 22Rv1 expressed lower levels of p57Kip2 when compared to PC-3 cells that expressed relatively low miR-21 and higher p57Kip2 (Figure 1B). Hence, we hypothesized that p57Kip2 may be a downstream target gene of miR-21 in prostate cancer. Regulation of target genes by binding of microRNA’s to the 3' untranslated regions (UTRs) of their mRNA’s has been well characterized. Hence, most of the bioinformatics tools available to predict miRNA target sites on mRNAs have been mainly focused on target sites within the 3'-UTRs of genes. A study by Hausser et al. suggest that miRNAs may combine targeting of coding regions and 3’ UTRs to flexibly tune their post-transcriptional regulatory effects [18]. Available microRNA prediction target algorithms did not predict p57Kip2 gene to be targeted by miR-21. However, scanning of human p57Kip2 gene for the miR-21 recognition site, revealed a 5-mer complementary binding site to the 2-7 seed region of mature miR-21 and a 3-mer sequence complementary to the 3’-target site (18-20 nucleotide) in the coding region of p57Kip2, which is from nucleotides 1054 to 1077 in accession # NM_00007 of GenBank as shown in Figure 1C. To experimentally test if miR-21 targets p57Kip2, we cloned this putative miR-21 recognition sequence of p57Kip2 in the 3’-UTR region of the luciferase gene in the pMIR reporter vector. Transfection of miR-21 mimic in PC-3 prostate cancer cells significantly (P < 0.01) reduced the luciferase activity (Figure 1D). However, mutation of the putative miR-21 binding nucleotides (Figure 1C and Additional file 1) abolished this effect. Similarly, luciferase activity was significantly (P < 0.01) increased when 22Rv1 prostate cancer cells, which were previously also shown to express relatively high endogenous miR-21 [16], were transfected with an anti-miR-21 to inhibit the endogenous miR-21 levels (Figure 1D), and the mutation of the miR-21 binding sites abrogated this effect.

MicroRNA-21 targets p57Kip2gene. (A) miR-21 and p57Kip2 are inversely correlated in 37 matched normal (N) and tumor (T) samples downloaded from the TCGA database. r: Pearson product-moment correlation coefficient. ****P < 0.0001 with one-way analysis of variance. (B) miR-21 (left panel) and p57Kip2 (right panel) levels were measured in prostate cancer cells with real-time RT-PCR and western blotting respectively. *P < 0.05 with one-way analysis of variance test. (C) Sequence alignment between miR-21 seed sequence and a partial coding or mutated sequence of p57Kip2. (D) Luciferase assay was performed with miR-21 mimic or anti-miR-21 transfection in cells with co-transfection of pMIR-3’UTR- p57Kip2 or pMIR-3’UTR-mutated p57Kip2 and a ß-Gal expression construct. **P < 0.01 with one-way analysis of variance and Tukey-Kramer post hoc test.
MicroRNA-21 downregulates p57Kip2 expression
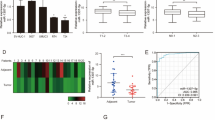
To further validate these findings, we checked the effect of miR-21 on p57Kip2 mRNA and protein levels. Inhibition of endogenous miR-21 with an anti-miR-21 in 22Rv1 and MDA-PCa-2b cells increased the p57Kip2 mRNA levels (P < 0.05), whereas miR-21 mimic decreased (P < 0.001) the basal level of p57Kip2 in PC-3 cells (Figure 2A). Transfection of anti-miR-21 in 22Rv1 and MDA-PCa-2b cells increased the p57Kip2 protein expression and miR-21 mimic transfection in PC-3 decreased p57Kip2 expression (Figure 2B). We used a CWR22 xenograft mouse model to study the effect on microRNA-21 and p57Kip2 expression under androgen depletion with castration [19]. We have previously shown that castration led to down-regulation of miR-21 as androgen signaling was shown to stimulate miR-21 [16]. We harvested tumors for miR-21 and p57Kip2 expression analysis at 14 and 40 days post castration, respectively. The results showed a significant (P < 0.0001) decrease in miR-21 expression levels in the castrated mice when compared to control non-castrated mice (Figure 2C) as was observed in our previous study. Interestingly, we observed an increase in p57Kip2 expression at 14 and 40 days in the castrated mice showing an inverse correlation between miR-21 and p57Kip2 expression in vivo (Figure 2C) (P < 0.0001). We show that for the first time, castration-mediated miR-21 decreases correlate with an increase in p57Kip2 under in vivo conditions. Currently, it is not clear how androgen/AR signaling regulates p57Kip2 expression in prostate cancer cells. Our results suggest that androgen/AR signaling-mediated inhibition of p57Kip2 expression appears to be mediated by miR-21 in CWR22 human prostate cancer xenografts. Further studies are needed to understand whether p57Kip2 expression contributes to castration-induced regression of prostate cancer.

MicroRNA-21 downregulates p57Kip2in prostate cancer cells. (A) 22Rv1 and MDA-PCa-2b cells were transfected with the inhibitor negative control and anti-miR-21 (50 nM) for 48 h. PC-3 cells were transfected with control siRNA and miR-21 mimic (20 nM) for 48 h. p57Kip2 gene expression was checked by real-time RT-PCR. *P < 0.05 and ***P < 0.001 with a Student’s t-test analysis. (B) Cell lysates were used for Western analysis to measure the level of p57Kip2 and GAPDH after anti-miR-21 inhibitor or miR-21 mimic transfection for 48 h. (C) p57Kip2 and miR-21 expressions were analyzed in CWR22 tumors after 14 and 40 days of castration. ****P < 0.0001 with one-way analysis of variance test.
MicroRNA-21 abrogates p57Kip2-mediated functional responses in prostate cancer cells
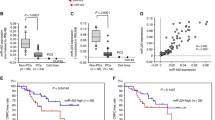
p57Kip2 has been shown previously to act as a tumor suppressor gene by inhibiting cell migration and invasion [20]. Hence, we tested whether endogenous miR-21 functionally abrogated p57Kip2 mediated tumor suppressive responses in prostate cancer cells. When we inhibited the endogenous miR-21 in PC-3 cells with an anti-miR-21 inhibitor, we observed an induction in p57Kip2 expression (Figure 3A). However, co-transfection with a p57Kip2 small-interfering RNA with anti-miR-21 inhibitor abolished the p57Kip2 induction by anti-miR-21 (Figure 3A). We also found that p57Kip2 inhibition significantly increased the cell migration in both PC-3 and 22Rv1 cells (Figure 3B). Transfection of cells with an anti-miR-21 inhibitor decreased the cell migration, although the results were not statistically significant. Interestingly, we found that the co-transfection of cells with anti-miR-21 and p57Kip2 siRNA, significantly increased cell migration when compared to anti-miR-21 treatment alone (Figure 3B) (P < 0.05 and P < 0.01). Similarly, while knocking down of endogenous miR-21 significantly reduced anchorage-independent growth, knocking down p57Kip2 reversed the inhibitory effect of the anti-miR-21 inhibitor on the anchorage-independent cell growth (Figure 3C) (P < 0.01 and P < 0.001). These results show that the functional effects of miR-21 in prostate cancer cells could be mediated partly by a downregulation in p57Kip2 expression.

MicroRNA-21 attenuates p57Kip2mediated functional responses in prostate cancer cells. Cells were transfected with 50 nM of control siRNA, anti-miR-21 or p57Kip2 siRNA either alone or in combination for 48 h. Cells were trypsinized and replated for Western analysis, cell migration or soft agar assay. (A) Western analysis in PC-3 cells for p57Kip2 and GAPDH was performed. (B) Number of cells migrated were quantified. *P < 0.05 and **P < 0.01 with one-way analysis of variance test. (C) After 14 days, colonies were stained and quantified in PC-3 and 22Rv1 cells for soft-agar colony formation assay. *P < 0.05, **P < 0.01 and ***P < 0.001 with one-way analysis of variance test.
In summary, we discovered p57Kip2 to be a novel target gene of microRNA-21 in prostate cancer. Our findings provide a novel mechanism of p57Kip2 downregulation in prostate cancer. These findings warrant further research to test if p57Kip2 is also a novel target gene of miR-21 in other cancer types as well. In our previous study, we found miR-21 to be an oncogenic regulator in prostate cancer by targeting the tumor suppressive effects of TGF-beta signaling pathway in cancer cells [16]. Given, the regulation of p57Kip2 by microRNA-21 in the current study, we provide a strong rationale to perform preclinical testing of microRNA-21 inhibitor as a novel therapeutic drug for prostate cancer.
References
Park MT, Lee SJ: Cell cycle and cancer. J Biochem Mol Biol. 2003, 36: 60-65. 10.5483/BMBRep.2003.36.1.060
Matsuoka S, Edwards MC, Bai C, Parker S, Zhang P, Baldini A, Harper JW, Elledge SJ: p57KIP2, a structurally distinct member of the p21CIP1 Cdk inhibitor family, is a candidate tumor suppressor gene. Genes Dev. 1995, 9: 650-662. 10.1101/gad.9.6.650
Lam WW, Hatada I, Ohishi S, Mukai T, Joyce JA, Cole TR, Donnai D, Reik W, Schofield PN, Maher ER: Analysis of germline CDKN1C (p57KIP2) mutations in familial and sporadic Beckwith-Wiedemann syndrome (BWS) provides a novel genotype-phenotype correlation. J Med Genet. 1999, 36: 518-523.
Yan Y, Frisen J, Lee MH, Massague J, Barbacid M: Ablation of the CDK inhibitor p57Kip2 results in increased apoptosis and delayed differentiation during mouse development. Genes Dev. 1997, 11: 973-983. 10.1101/gad.11.8.973
Jin RJ, Lho Y, Wang Y, Ao M, Revelo MP, Hayward SW, Wills ML, Logan SK, Zhang P, Matusik RJ: Down-regulation of p57Kip2 induces prostate cancer in the mouse. Cancer Res. 2008, 68: 3601-3608. 10.1158/0008-5472.CAN-08-0073
Kavanagh E, Joseph B: The hallmarks of CDKN1C (p57, KIP2) in cancer. Biochim Biophys Acta. 1816, 2011: 50-56.
Li Y, Nagai H, Ohno T, Yuge M, Hatano S, Ito E, Mori N, Saito H, Kinoshita T: Aberrant DNA methylation of p57(KIP2) gene in the promoter region in lymphoid malignancies of B-cell phenotype. Blood. 2002, 100: 2572-2577. 10.1182/blood-2001-11-0026
Hagiwara K, Li Y, Kinoshita T, Kunishma S, Ohashi H, Hotta T, Nagai H: Aberrant DNA methylation of the p57KIP2 gene is a sensitive biomarker for detecting minimal residual disease in diffuse large B cell lymphoma. Leuk Res. 2010, 34: 50-54. 10.1016/j.leukres.2009.06.028
Kamura T, Hara T, Kotoshiba S, Yada M, Ishida N, Imaki H, Hatakeyama S, Nakayama K, Nakayama KI: Degradation of p57Kip2 mediated by SCFSkp2-dependent ubiquitylation. Proc Natl Acad Sci U S A. 2003, 100: 10231-10236. 10.1073/pnas.1831009100
Calvisi DF, Pinna F, Ladu S, Muroni MR, Frau M, Demartis I, Tomasi ML, Sini M, Simile MM, Seddaiu MA, Feo F, Pascale RM: The degradation of cell cycle regulators by SKP2/CKS1 ubiquitin ligase is genetically controlled in rodent liver cancer and contributes to determine the susceptibility to the disease. Int J Cancer J Int du Cancer. 2010, 126: 1275-1281.
Fornari F, Gramantieri L, Ferracin M, Veronese A, Sabbioni S, Calin GA, Grazi GL, Giovannini C, Croce CM, Bolondi L, Negrini M: MiR-221 controls CDKN1C/p57 and CDKN1B/p27 expression in human hepatocellular carcinoma. Oncogene. 2008, 27: 5651-5661. 10.1038/onc.2008.178
Sengupta S, Nie J, Wagner RJ, Yang C, Stewart R, Thomson JA: MicroRNA 92b controls the G1/S checkpoint gene p57 in human embryonic stem cells. Stem Cells. 2009, 27: 1524-1528. 10.1002/stem.84
Kim YK, Yu J, Han TS, Park SY, Namkoong B, Kim DH, Hur K, Yoo MW, Lee HJ, Yang HK, Kim VN: Functional links between clustered microRNAs: suppression of cell-cycle inhibitors by microRNA clusters in gastric cancer. Nucleic Acids Res. 2009, 37: 1672-1681. 10.1093/nar/gkp002
Medina R, Zaidi SK, Liu CG, Stein JL, van Wijnen AJ, Croce CM, Stein GS: MicroRNAs 221 and 222 bypass quiescence and compromise cell survival. Cancer Res. 2008, 68: 2773-2780. 10.1158/0008-5472.CAN-07-6754
Lodygin D, Epanchintsev A, Menssen A, Diebold J, Hermeking H: Functional epigenomics identifies genes frequently silenced in prostate cancer. Cancer Res. 2005, 65: 4218-4227. 10.1158/0008-5472.CAN-04-4407
Mishra S, Deng JJ, Gowda PS, Rao MK, Lin CL, Chen CL, Huang T, Sun LZ: Androgen receptor and microRNA-21 axis downregulates transforming growth factor beta receptor II (TGFBR2) expression in prostate cancer. Oncogene. 2014, 33: 4097-4106. 10.1038/onc.2013.374
Shen J, Hruby GW, McKiernan JM, Gurvich I, Lipsky MJ, Benson MC, Santella RM: Dysregulation of circulating microRNAs and prediction of aggressive prostate cancer. Prostate. 2012, 72: 1469-1477. 10.1002/pros.22499
Hausser J, Syed AP, Bilen B, Zavolan M: Analysis of CDS-located miRNA target sites suggests that they can effectively inhibit translation. Genome Res. 2013, 23: 604-615. 10.1101/gr.139758.112
Nagabhushan M, Miller CM, Pretlow TP, Giaconia JM, Edgehouse NL, Schwartz S, Kung HJ, de Vere White RW, Gumerlock PH, Resnick MI, Amini SB, Pretlow TG: CWR22: the first human prostate cancer xenograft with strongly androgen-dependent and relapsed strains both in vivo and in soft agar. Cancer Res. 1996, 56: 3042-3046.
Guo H, Lv Y, Tian T, Hu TH, Wang WJ, Sui X, Jiang L, Ruan ZP, Nan KJ: Downregulation of p57 accelerates the growth and invasion of hepatocellular carcinoma. Carcinogenesis. 2011, 32: 1897-1904. 10.1093/carcin/bgr220
Acknowledgements
We thank Dr. Majeet Rao for the pMIR luciferase construct and Dr. Clifford G. Tepper for the CWR22 xenograft tissue. This work was supported in part by funding from NIH grants RO1CA079683, RO1CA172886, and P30CA054174.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
SM was involved in the conception and design of the project, acquisition and analysis of the data, drafting of the manuscript and final approval. CLL, TH and HB have made substantial contributions to the acquisition and analysis of the data and were involved in the drafting of the manuscript. LZS conceived the study, was involved in planning of the project, participated in the design and analysis of the data, drafting of the manuscript, and has given final approval on the manuscript. All authors read and approved the final manuscript.
Electronic supplementary material
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Mishra, S., Lin, CL., Huang, T.HM. et al. MicroRNA-21 inhibits p57Kip2 expression in prostate cancer. Mol Cancer 13, 212 (2014). https://doi.org/10.1186/1476-4598-13-212
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1476-4598-13-212