
Abstract
Background
Advanced glycation endproducts (AGEs) contribute to the development of vascular complications of diabetes and have been recently implicated in the pathogenesis of diabetes. Since AGEs are generated within foodstuffs upon food processing, it is increasingly recognised that the modern diet is replete with AGEs. AGEs are thought to stimulate chronic low-grade inflammation and promote oxidative stress and have been linked to the development of insulin resistance. Simple therapeutic strategies targeted at attenuating the progression of chronic low-grade inflammation and insulin resistance are urgently required to prevent or slow the development of type 2 diabetes in susceptible individuals. Dietary modulation of the human colonic microbiota has been shown to confer a number of health benefits to the host, but its effect on advanced glycation is unknown. The aim of this article is to describe the methodology of a double-blind placebo-controlled randomised crossover trial designed to determine the effect of 12 week consumption of a prebiotic dietary supplement on the advanced glycation pathway, insulin sensitivity and chronic low-grade inflammation in adults with pre-diabetes.
Methods/Design
Thirty adults with pre-diabetes (Impaired Glucose Tolerance or Impaired Fasting Glucose) aged between 40–60 years will be randomly assigned to receive either 10 grams of prebiotic (inulin/oligofructose) daily or 10 grams placebo (maltodextrin) daily for 12 weeks. After a 2-week washout period, study subjects will crossover to receive the alternative dietary treatment for 12 weeks. The primary outcome is the difference in markers of the advanced glycation pathway carboxymethyllysine (CML) and methylglyoxal (MG) between experimental and control treatments. Secondary outcomes include HbA1c, insulin sensitivity, lipid levels, blood pressure, serum glutathione, adiponectin, IL-6, E-selectin, myeloperoxidase, C-reactive protein, Toll-like Receptor 4 (TLR4), soluble receptor for AGE (sRAGE), urinary 8-isoprostanes, faecal bacterial composition and short chain fatty acid profile. Anthropometric measures including BMI and waist circumference will be collected in addition to comprehensive dietary and lifestyle data.
Discussion
Prebiotics which selectively stimulate the growth of beneficial bacteria in the human colon might offer protection against AGE-related pathology in people at risk of developing type 2 diabetes.
Trial registration
Australia and New Zealand Clinical Trials Register (ANZCTR): ACTRN12613000130763.
Similar content being viewed by others
Background
Advanced Glycation Endproducts (AGEs) are formed via the Maillard reaction, which consists of a complex network of non-enzymatic reactions involving the carbonyl groups of reducing sugars which react with the amino groups of proteins [1]. AGEs are generated in vivo as a normal consequence of metabolism, but their formation is accelerated under conditions where blood glucose is chronically elevated such as poorly controlled diabetes [2]. AGE formation is also increased in the presence of oxidative stress, which is frequently observed in individuals with the metabolic syndrome [3, 4]. Overproduction of reactive oxygen species (ROS) can result in maladaptive responses including interruption of cellular glycolysis, which can generate highly reactive dicarbonyl compounds capable of rapid AGE formation [5].
Excessive AGE accumulation can lead to several pathophysiological consequences. AGE-modification of proteins results in changes in structure and/or function. For example, the AGE-modification of extracellular collagen reduces its elasticity and solubility, and results in increased stiffness, disturbed cellular adhesion and reduced turnover contributing to basement membrane thickening [6]. Intracellularly, AGE-modification of mitochondrial proteins is associated with suppression in the activity of respiratory chain enzymes and overproduction of ROS [7, 8]. Indeed, glycated proteins provide stable active sites for catalysing the formation of free radicals [9]. Finally, AGEs are able to bind and activate a range of receptors, which then trigger a downstream cascade of pathogenic mediators. Interaction of AGEs with the Receptor for AGEs (RAGE) promotes activation of the transcription factor nuclear factor kappa-B (NF-κB), with subsequent upregulation of chemokines, such as MCP-1 and profibrogenic mediators such as TGFβ in addition to pro-inflammatory cytokines which are known to be involved in thrombogenesis, vascular inflammation and pathological angiogenesis. These RAGE-mediated events contribute to many of the long-term complications of diabetes [10]. AGE/RAGE ligation also promotes overproduction of ROS which can then activate NF-κB [11], a key driver of inflammation.
More recently, AGEs have been implicated in the pathogenesis of both type 1 and type 2 diabetes. Several studies have shown that AGEs are associated with insulin resistance [12, 13], and can induce low-grade inflammation [14] and pancreatic beta cell dysfunction [15, 16].
In contrast to endogenous AGE formation, AGEs are also absorbed by the body from exogenous sources such as cigarette smoke and through consumption of processed foods [17]. Since AGEs are generated within foodstuffs upon heating and food processing, it is increasingly recognised that the modern diet is replete with AGEs [18]. AGE-restricted diets can arrest the development of type 2 diabetes in animal models [19]. A recent study found that excess consumption of AGE-precursors in mice over several generations led to the development of insulin resistance [20]. Human trials have found that dietary AGE restriction can improve insulin sensitivity [21, 22] and decrease markers of oxidative stress [23] or inflammation [24]. Further studies are required to confirm the long-term benefits of dietary AGE-restriction in humans [25]. However, simple, safe and effective interventions which prevent or minimise excessive AGE accumulation and subsequent AGE-related pathology in people with diabetes and/or in those at risk of developing the condition are warranted.
Interventions which influence the human intestinal microbiota are worthy of further investigation given that specific micro-organisms have the ability to significantly affect host metabolism. Gut bacteria play an important role in the host immune system, modulation of inflammatory processes, extraction of energy from the host’s diet, fermentation of dietary fibres to produce short-chain fatty acids, alteration of human gene expression, regulation of intestinal permeability, production of some vitamins and promotion of mineral absorption by the host [26–31]. Furthermore, the total quantity and relative proportions of distinct bacterial species found in the colon differ between lean and obese individuals as well as between individuals with and without diabetes [32, 33].
It is thought that certain dietary AGEs are largely undigested by human gut enzymes and eventually enter the colon, where they may act as a growth substrate for detrimental bacteria such as some Clostridium and Bacteroides species [34]. Therefore it is conceivable that individuals who consume highly processed diets (which contain large quantities of AGEs) may adversely alter their colonic microbial composition, potentially enhancing their risk for the development of metabolic diseases such as obesity and type 2 diabetes [35].
Therapeutic manipulation of the gut microbiota and restoration of normobiosis could potentially reduce circulating AGE levels and improve the metabolic health of individuals at risk for the development of type 2 diabetes. Regular consumption of prebiotics to promote the growth of beneficial gut bacterial flora is one such avenue currently under investigation. Prebiotics are non-digestable plant-derived carbohydrates which confer health benefits to the host by acting as a fermentation substrate in the colon, stimulating the preferential growth and activity of a limited number of beneficial microbial species [36]. Supplementation of the human diet with prebiotic fructans such as inulin or fructo-oligosaccharides alters the bacterial composition of the large intestine by favouring the selective proliferation of beneficial lactic acid-producing species such as bifidobacteria and lactobacilli. Prebiotic-stimulated increases in intestinal Bifidobacterium species have been shown to attenuate the production of ROS and markers of inflammation in individuals consuming high fat diets [37].
While the complex interactions between diet, intestinal microbiota and host metabolism are still being elucidated, no studies have investigated the effect of dietary prebiotics on circulating AGE concentrations. This trial was designed to investigate the effect of a prebiotic dietary supplement on AGE accumulation and explore changes to the growth and activity of specific gut microbiota in adults diagnosed with prediabetes.
Methods/Design
Study design and setting
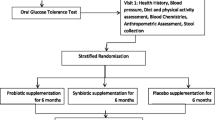
This is a 6.5-month randomised crossover controlled clinical trial (RCT) in which adults aged 40–60 years with diagnosed pre-diabetes will be enrolled. Potential study participants will be identified from General Practice (GP) clinics throughout South Gippsland, Victoria. The study design is presented in Figure 1.

Trial protocol.
Inclusion criteria
Individuals aged between 40–60 years and diagnosed with prediabetes (Impaired Fasting Glucose or Impaired Glucose Tolerance) within the previous 12 months. Diagnosis will have been made at each individual’s local GP clinic after undertaking an Oral Glucose Tolerance Test (OGTT). Prediabetes was defined as a fasting plasma glucose concentration ≥ 6.1 and < 7.0 mmol/L followed by a 2-hour post glucose load glucose concentration < 7.8 mmol/L, or a fasting plasma glucose < 7.0 mmol/L followed by a 2-hour post glucose load glucose concentration ≥ 7.8 and < 11.1 mmol/L [38].
Exclusion criteria
Individuals previously diagnosed with type 1, type 2 diabetes or impaired renal function (eGFR <90 mL/min/1.73 m2), individuals with known gastrointestinal pathology (coeliac disease, inflammatory bowel disease), pregnant women, smokers, individuals who have taken antibiotics, dietary prebiotic or probiotic nutritional supplements within the previous three months, individuals taking aspirin or Vitamin B, individuals who have made major dietary or lifestyle changes in the previous three months, individuals who are unwilling to provide blood, urine and stool samples or are unable to attend their local pathology collection centre.
Ethics
The trial has received ethical approval from the Monash University Human Research Ethics Committee.
Sample size calculation
The minimum difference we wish to detect is 0.4 micromol/L serum CML (20% reduction in CML), with a standard deviation of 0.4 [39] with 5% Significance and 80% Power. This calculates as a total sample size of 18 individuals, plus 12 individuals to allow for withdrawals = 30 subjects required. Epidemiological studies have demonstrated a positive correlation between serum CML and all-cause and cardiovascular mortality, cardiovascular disease, glucose intolerance, impaired insulin secretion, renal impairment and diabetic vascular complications [40, 41]. A 0.4 micromol/L increase in serum CML concentration represented a 68% increased risk for all-cause mortality over seven years in a large prospective cohort study [42].
Baseline assessment
The study timeline is presented in Figure 2. Following recruitment and screening, consenting participants will undergo a baseline assessment at their local GP clinic. The baseline assessment will be undertaken by the research dietitian, who will collect demographic details, medical and social history (living situation, marital status, current occupation), physical activity questionnaire, dietary intake assessment, anthropometric measurements including Body Mass Index (BMI) and waist circumference. Participants will also be instructed to attend the local pathology centre to provide a stool sample, 24-hour urine collection, and have blood taken for analysis.

Study flowchart.
Randomisation
Randomisation of participants to intervention/placebo sequence will be completed by a local pharmacist using a permuted-block randomisation stratified by gender via the web site http://www.randomization.com. The pharmacist will distribute the experimental and placebo nutritional supplements to participants in the appropriate sequence. Other than provision of supplements, the pharmacist will have no contact with study participants or involvement in data collection or analysis. Dietary supplements will be provided in sealed opaque packages which will be weighed at the conclusion of the study in order to assess compliance. The supplements will be packaged off-site by an external pharmaceutical packaging agency. Each package will contain a participant’s study identification number and will be labelled as either Supplement 1 or Supplement 2 corresponding to the first and second intervention periods. This will ensure all investigators and participants are blinded to the treatment. Blinding will cease only after statistical analysis of the data has been completed.
Intervention
Participants will be randomly assigned to receive either 10 g of chicory-derived inulin/oligofructose powder (kindly provided by Beneo-Orafti Active Food Ingredients, Belgium) (intervention) or 10 g of maltodextrin powder (placebo) daily. Previous studies have demonstrated the bifidogenic effect of daily consumption of 5 g and 8 g dietary inulin supplements [43]. The inulin and maltodextrin powders to be consumed are both tasteless and can be mixed into hot or cold liquids or semi-solid foods. Participants will be instructed how to incorporate their supplement into their usual diet, and advised to gradually increase their dose over ten successive days until the target dose is reached. This stepped escalation in supplement dose aims to minimise gastrointestinal discomfort for participants, as a sudden increase in dietary prebiotic intake may result in increased stool frequency, abdominal bloating and flatulence until the bowel adapts to the increased fibre intake [44]. Written instructions will also be provided. Participants will be advised to consume each dietary supplement daily for 90 days, and otherwise maintain their usual dietary intake and level of physical activity. Gastrointestinal tolerance to the dietary supplements will be indicated by each study participant using a visual analogue scale as described below.
Follow up visits
Visits to the GP clinic will be scheduled for each participant at the conclusion of both Intervention 1 and Intervention 2 treatment periods, in order to provide follow-up data. Information collected will include anthropometric measurements, dietary intake assessment and physical activity assessment. Each participant will complete a self-administered questionnaire designed to assess gastrointestinal tolerance to the dietary supplement. Completed questionnaires will be placed into sealed opaque envelopes in order to maintain blinding of the researcher collecting data during the follow-up visits. Participants will also be instructed to attend the local pathology centre to provide urine and stool samples, and have blood taken for analysis.
Safety considerations
All adverse events will be documented.
Outcome measurements
The primary outcome of interest is the difference in serum AGE and AGE-precursor concentration (measured as CML and MG respectively) between experimental and control treatments. Secondary outcomes include HbA1c, insulin resistance (measured indirectly by homeostasis model assessment), antioxidant capacity (reduced glutathione), markers of oxidative stress on lipid molecules (urinary 8-isoprostanes), inflammatory biomarkers (serum IL-6, high sensitivity C-reactive protein, MCP-1, sRAGE), adhesion molecules (E-selectin), gut barrier integrity (TLR4), 24-hour urine and faecal CML and MG concentrations, faecal bacterial composition (Bifidobacterium spp., Lactobacillus spp., Roseburia spp., Faecalibacterium prausnitzii and Akkermansia muciniphila) and faecal short chain fatty acid concentrations. Lipid levels (serum total cholesterol, LDL, HDL, TG) and blood pressure will also be measured. Anthropometric measurements including BMI and waist circumference will be collected, in addition to comprehensive dietary and lifestyle data. Gastrointestinal side-effects will be assessed using a visual analogue scale adapted from Lecerf et al.[45]. The scale rates nine items including flatulence, bloating, rumbling, abdominal cramps, a global digestive tolerance score calculated from the five previous items, stool consistency, stool frequency and general well-being through disturbances in usual and professional activities as well as disturbances in personal and social activities. Items are scored on a 10 cm linear scale.
Data collection
Dietary intake
An experienced research dietitian will obtain a comprehensive dietary history from each study participant at baseline and at the completion of each intervention period. A dietary history is a structured interview method consisting of questions regarding habitual food intake. It involves a 24-hour diet recall followed by a food frequency questionnaire to clarify information about usual consumption over the previous month. Usual portion sizes will be obtained in household measures and with the use of photographic aids. The dietitian will review the dietary history with each participant, probing for details on portion sizes and cooking methods in order to improve data accuracy. Dietary AGE content will be estimated from an open source database which lists the AGE concentration of foods using validated analytical techniques [46]. Dietary macro and micronutrient intakes will be estimated using the Foodworks nutrient software program (Xyris Software, NSW, Australia). Each participant will also be randomly contacted by telephone during the study and asked to provide a 24-hour diet recall, and changes in urinary urea excretion will be monitored in order to validate the dietary history data collected [47]. Physical activity will be estimated by asking participants to complete the International Physical Activity Questionnaire (Short Form) prior to and at the completion of experimental and placebo intervention periods [48].
Anthropometry
Body weight will be measured in participants wearing light clothes without shoes using a digital scale (Seca, Germany) to the nearest 0.1 kg. Height will be measured using a portable stadiometer (Seca, Germany) to the nearest 0.1 cm. BMI is calculated by dividing weight (kg) per height (m) square. Waist circumference will be measured at the midpoint of the lowest rib and iliac crest using a measuring tape to the nearest 0.1 cm. Body composition (total body water, fat mass, fat-free mass) will be determined using Bioelectrical Impedance Analysis (BodyStat-1500, Bodystat, Douglas, Isle of Man, United Kingdom). Ambulatory blood pressure will be measured using an electronic blood pressure machine (Omron Corporation, Kyoto, Japan), with subjects at rest in a seated position. All anthropometric measurements will be conducted in duplicate, with the mean measurement recorded.
Laboratory investigations
Biological samples will be collected at baseline and at completion of each three-month supplement intervention period and stored at −80°C immediately after collection. Twenty ml of fasting venous blood will be collected from each participant by phlebotomy into a sodium fluoride EDTA tube, a heparin-lined vacuum tube and a clean glass test tube. Twenty-four hour urine collections and morning stool samples will be collected in sterile containers.
Serum, urinary and faecal CML will be measured using a competitive ELISA (AGE-CML ELISA, Microcoat, Penzberg, Germany) [49]. This assay has been validated [50], is specific, and shows no cross-reactivity with other compounds [49]. The within assay and between-assay coefficient of variation are both less than 5%, respectively. Methylglyoxal will be measured by HPLC. Serum total cholesterol and triglyceride concentrations will be determined by enzymatic colorimetric assay (Technicon Instruments, Ltd., New York, N.Y., USA), while HDL cholesterol will be determined enzymatically in the supernatant after precipitation of other lipoproteins with dextran sulphate-magnesium. LDL-cholesterol will be calculated using the Friedewald formula. Plasma glucose levels will be determined by using an automated glucose oxidase method (Glucose analyser 2, Beckman Instruments, Fullerton, California). Insulin will be measured by enzymatic colorimetry (WAKO Pure-Chemical Industries, Osaka, Japan). Insulin Resistance (IR) will be estimated by the homeostasis model assessment (HOMA) index as [FI × (fasting glucose/22.5)], where FI is insulin in microunits per millilitre and fasting glucose is in millimoles per litre [51, 52]. HbA1c will be measured by autoanalyser (Roche Diagnostics, Mannheim, Germany). Plasma IL-6, MCP-1, E-selectin, hsCRP, TLR4, glutathione (GSH), and myeloperoxidase will be measured by commercial ELISA kits (Biosource International, Camarillo, CA, USA). Urine 8-isoprostanes will be measured by ELISA (Oxford Biomedical Research, MI, USA).
Stool samples will be homogenised in a blender and stored at −20°C for SCFA analysis. Samples will be thawed and 5 g aliquots placed in Centriprep fluid concentrators, MWCO 30,000 kDa (Amicon Inc., Beverly, MA, USA). Samples will be centrifuged for 30 minutes at 1000 × g, room temperature and supernatants placed in 15 ml polypropylene tubes. 0.3 ml of 25% m-phosphoric acid will be added to each tube, samples will be vortexed and incubated at room temperature for 25 minutes. Samples will be centrifuged at 5000 × g for 15 minutes at room temperature. Supernatants will be decanted and frozen overnight. The following day, samples will be thawed and the pH of each sample adjusted to 6.5 using 4 N KOH. Oxalic acid will be added at a final concentration of 0.03% and SCFA concentrations determined by gas chromatography with use of a Hewlett-Packard 5880A gas chromatograph (Hewlett Packard, Palo Alto, CA, USA) containing an 80/120 Carbopack B-DA/4% Carbowax 20 M column (Supelco Inc., Bellefonte, PA, USA).
Quantitative Real-time PCR will be used to determine faecal concentrations of Bifidobacterium spp., Lactobacillus spp., Roseburia spp., Faecalbacterium prausnitzii and Akkermansia muciniphila. The primers used will be based on the following 16S rRNA gene sequences: Bifidobacterium spp: F-CTCCTGGAAACGGGTGG and R-GGTGTTCTTCCCGATATCTACA [53], Lactobacillus spp: F-AGCAGTAGGGAATCTTCCA and R-CACCGCTACACATGGAG [54], Roseburia spp: F-CGKACTAGAGTGTCGGAGG and R-GTCATCTAGAGTGTCGGAGG [55], Faecalbacterium prausnitzii: F-GGAGGAAGAAGGTCTTCGG and R-AATTCCGCCTACCTCTGCACT [56], and Akkermansia muciniphila: F-CAGCACGTGAAGGTGGGGAC and R-CCTTGCGGTTGGCTTCAGAT [57]. PCR amplification and detection will be achieved with an ABI 7300 Real-time PCR system (Applied Biosystems, Foster City, CA, USA) using Mighty Amp for Real-time (SYBR Plus) and Rox Reference Dye (Invitrogen, Carlsbad, CA, USA). Each assay will be performed in duplicate in the same run. The cycle threshold of each sample will then be compared with a standard curve (performed in duplicate) made by diluting genomic DNA (tenfold serial dilution). Prior to isolating the DNA, the cell counts will be determined in culture and expressed as “colony forming units” (CFU). Data will be expressed as log CFU/g of faeces.
Statistical analysis
Outcome analyses will be undertaken on an intention-to-treat basis. Data will be presented as means ± SD. The Kolmogorov-Smirnov goodness-of-fit test will be used to test for normal distribution, and data not normally distributed will be log-transformed. Correlation analyses will be performed using the Pearson correlation coefficient. Significance of changes during the study will be assessed by comparing change of means between placebo and prebiotic treatment periods by paired sample t-tests. Trial data will be analysed using a linear mixed model design based on repeated measures to account for fixed factors such as treatment sequence (inulin – placebo vs placebo - inulin) and treatment period (intervention 1 vs intervention 2) in addition to participants as a random factor. Significant differences will be defined as a value of P < 0.05 based on two-sided tests. Any differences in physical activity levels, anthropometry, energy or nutrient intake during the course of the trial will be identified using ANOVA. Gastrointestinal symptom data obtained by visual analogue scale will be analysed using the Wilcoxon signed rank test. Effect sizes including 95% confidence intervals will be calculated for all significant outcomes. Data analysis will be performed using SPSS 20.0 software (SPSS, Chicago, IL).
Discussion
AGEs are derived from both exogenous and endogenous sources, and the rate at which AGEs accumulate in the body is dependent to a large extent on the chronological age, lifestyle and metabolic health of an individual. Smoking cigarettes and consuming foods containing high concentrations of AGEs (and their precursors) increases the accumulation of AGEs from exogenous sources. Endogenous AGE formation is accelerated under conditions of hyperglycaemia, dyslipidaemia and increased oxidative stress, conditions that are common in individuals with diabetes and in those at risk of developing type 2 diabetes (such as those with prediabetes and the metabolic syndrome) [58]. Moreover, in individuals with impaired renal function, urinary AGE excretion may be diminished resulting in a greater accumulation of AGEs in the body [59].
Risk factors for the development of type 2 diabetes include obesity, hypertension and cardiovascular disease; conditions that are commonly associated with unhealthy lifestyles including poor food habits. Restricting the intake of foods high in AGEs might potentially reduce AGE accumulation, but adherence to such diets can be challenging given that foods high in AGEs are very palatable due to their enhanced flavour, colour and aroma [60].
Supplementation of the diet with bifidogenic prebiotic fibres (such as inulin) may reduce or retard the accumulation of AGEs in individuals at risk of developing type 2 diabetes. Prebiotics have been shown to improve and restore optimal microbial balance within the gastrointestinal tract, potentially reducing AGE absorption and/or production by the human host. Preliminary investigations indicate that consuming a high-AGE diet is sufficient to favour the proliferation of potentially pathogenic colonic bacteria over more beneficial species. Consumption of glycated proteins [34], fried meats [61] and toasted wheat flakes [62] encouraged the preferential growth of greater numbers of detrimental gram negative and sulphate-reducing colonic micro-organisms when compared to control diets. Short Chain Fatty Acids (SCFAs) produced as a bacterial by-product of prebiotic fermentation act to lower the intestinal pH, inhibiting the growth of protein-degrading micro-organisms capable of producing potentially toxic metabolites. SCFAs also stimulate colonic smooth muscle contractions, speeding intestinal transit and limiting the time available for protein fermentation and putrefaction to occur in the gut [63]. Therapeutic manipulation of the gut microbiota with prebiotics may restore gut normobiosis and reduce AGE accumulation in humans at risk for type 2 diabetes development by the following mechanisms:
Maintenance of gut barrier function
Kinetic studies have estimated that up to 30% of dietary AGEs consumed are intestinally absorbed [64]. Under circumstances of increased intestinal permeability, it is likely that greater quantities of dietary AGEs and their reactive dicarbonyl precursors may be able to gain entry into the systemic circulation. Elevated levels of circulating proinflammatory cytokines and ROS frequently observed in individuals with prediabetes are known to compromise tight junctions between cells, disrupting the integrity of the intestinal barrier and enabling the absorption of larger, potentially deleterious compounds [65]. Prebiotic fermentation products may reduce gastrointestinal permeability and as a result limit the absorption of exogenous AGEs.
The SCFAs acetate and butyrate are intricately involved in the maintenance of gut epithelial integrity. Acetate increases colonic blood flow and butyrate is the principal fuel for colonocytes, assisting to optimise epithelial cell health [66]. Butyrate reduces gastrointestinal permeability by enhancing the activation of the peroxisomal proliferator-activated receptor gamma (PPARgamma) gene, a nuclear receptor involved in the attenuation of inflammation in colonic epithelial cells [67]. Butyrate also upregulates the expression of mucin-associated genes important in maintaining the integrity of the intestinal mucosal barrier [68]. Oligofructose supplementation in mice has been shown to increase the expression of zonula and occludin, proteins important in the maintenance of tight junctions between gastrointestinal cells [69], and a dietary inulin intervention reduced markers of intestinal permeability in healthy adult males [70]. Butyrate is a histone deacetylase inhibitor and is likely to promote the transcription of these tight junction proteins. Prebiotic-induced changes in gut microbiota also increase endogenous production of Glucagon-like peptide 2 (GLP-2), which enhances gut barrier function by promoting the proliferation of crypt cells [71, 72].
Reduction of oxidative stress, inflammation and insulin resistance
Increased production of ROS stimulate endogenous AGE formation by oxidising glucose and unsaturated fatty acids to generate reactive dicarbonyls. Experimental drug treatments which attenuate oxidative stress have previously demonstrated reductions in serum AGE levels [73]. Cytokine production at sites of inflammation stimulate immune cell activation of NADPH oxidase (NOX) and production of myeloperoxidase, enzymes involved in the oxidation of amino acids to form AGE precursors [74]. Activated immune cells can also secrete the high-mobility group box 1 (HMGB-1) protein which is capable of binding to RAGE, thereby inducing further inflammation [75].
A high fat diet (independent of the level of obesity in the host) is associated with negative changes in bacterial communities within the colon [76]. In response to a high fat meal, bacterial lipopolysaccharide (LPS) translocates from the intestine into the host circulation, resulting in ‘metabolic endotoxemia’ [77, 78]. LPS is a major component of the outer membrane of gram negative bacteria, and is a potent activator of the mammalian immune system. LPS interaction with immune cells stimulates macrophage over-production of ROS, enhances the secretion of pro-inflammatory cytokines, and contributes to weight gain and development of insulin resistance [79, 80]. Individuals with type 2 diabetes have been found to possess endotoxemia levels 2-fold higher than people without diabetes [81]. Metabolic endotoxemia also positively correlates with total energy intake and fasting insulin levels in the general population [82]. In mice with high fat diet-induced metabolic endotoxemia, nutritional supplementation of the diet with prebiotics restored intestinal levels of gram positive bacteria (particularly Bifidobacterium species) and subsequently improved glucose tolerance and reduced circulating concentrations of LPS and pro-inflammatory cytokines [83]. Human trials involving dietary prebiotic supplementation have successfully reduced serum LPS levels [45] and markers of lipid peroxidation (a process which generates AGE precursors), possibly through the reduction of ROS production or the direct antioxidant ability of some Bifidobacterium and Lactobacillus bacterial species [84, 85].
SCFAs produced as a bacterial by-product of prebiotic fermentation are absorbed into the host circulation, effecting the expression of a wide range of genes in distal tissues associated with cell proliferation, differentiation and apoptosis. SCFAs are ligands for the G-protein coupled receptors (GPRs) GPR41 and GPR43 [86] on immune cells. These receptors are involved in down-regulating inappropriate immune cell production of pro-inflammatory cytokines, chemokines and ROS [87]. The SCFAs acetate, proprionate and butyrate exhibit a variety of anti-inflammatory actions through inhibition of NFκB activation, prevention of LPS-stimulated TNFα production in neutrophils and suppression of cytokine production [31, 88–90].
Numerous other immune modulating effects have been observed secondary to gut bacterial activity including the production of anti-inflammatory compounds such as polysaccharide A [91], peptidoglycan [92] and conjugated linoleic acid [93], and the induction of T-regulatory cells [26].
Promotion of weight reduction
An energy-restricted diet resulting in weight loss has been shown to reduce serum AGE levels in overweight and obese individuals [94]. The consumption of prebiotics in human clinical trials has promoted self-reported satiety [95], weight reduction, reduced production of the orexigenic hormone grehlin and stimulated expression of the appetite-reducing hormone peptide YY (PYY) [96]. Interaction with GPR41 by the SCFAs proprionate and butyrate increases satiety [97], upregulates PYY production and modulates the expression of leptin, a hormone important in controlling energy intake and expenditure [31].
In mice, the selective growth of certain lactobacillus species in the colon reduced body fat storage through the up-regulation of fiaf (fasting induced adipose factor) gene expression and inhibition of lipoprotein lipase (LPL) [98–100]. These findings may have the potential to affect weight reduction in humans and subsequently reduce circulating AGE levels.
Enhanced antioxidant capacity
Inulin enhances the proliferation of lactic acid producing bacteria capable of synthesising B-group vitamins, some of which have an antioxidant capacity [101]. These vitamins can be utilised by the human host to neutralise ROS. Vitamins B1 and B6 trap the carbonyl groups of highly reactive AGE precursors before they can react with proteins [102]. Some Lactobacillus and Bifidobacterium species are efficient scavengers of the lipid peroxidation product malandialdehyde, protecting the host from excessive accumulation of this toxic AGE precursor [84]. Inulin also exhibits antioxidant properties independent of altering gut bacterial growth and is able to scavenge a number of ROS, which may help to reduce lipid peroxidation in the stomach [103].
The SCFA butyrate, produced as a bacterial by-product of inulin fermentation, has been shown to increase colonic glutathione production [104]. Glutathione is an antioxidant co-factor required for glyoxalase I activity, an enzyme which degrades the AGE precursor methylglyoxyl. Increased production of ROS is also thought to deplete glutathione levels. Through the reduction of oxidative stress, prebiotics may assist in the maintainance or upregulation of the glyoxylase pathway.
Reduction of hyperglycemia
Both transient and chronic elevations in blood glucose increase endogenous AGE generation. Activation of GPR43 in adipocytes by proprionate inhibits lipolysis and lowers glycemia in healthy individuals [105]. Butyrate has been shown to reverse diet-induced insulin resistance in animal studies [106], possibly by enhancing PPARgamma expression which increases fatty acid oxidation in muscle.
Glucagon-like peptide 1 (GLP-1) is an incretin hormone released from intestinal L-cells in response to consumption of carbohydrates and fats. GLP-1 potentiates glucose-induced insulin secretion, reducing post-prandial blood glucose levels. GLP-1 also enhances satiety and slows gastric emptying. Prebiotic feeding in rats promoted L-cell differentiation in the colon and increased GLP-1 production [107, 108], probably through an increase in bacterial production of butyrate.
The consumption of inulin as a dietary supplement may also influence circulating AGE concentrations independently of its prebiotic function. High glycemic index (GI) diets and their resultant hyperglycemic effect have been shown to enhance AGE formation in healthy individuals [109]. Inulin is a soluble fibre which when consumed daily may play a role in the reduction of the GI of the diet. Many soluble fibres are known for their ability to delay gastric emptying and slow the rate of intestinal nutrient absorption, reducing the GI of the carbohydrates in the meal. Regular consumption of prebiotic soluble fibre reduces fasting and post-prandial serum glucose levels in people with impaired glucose tolerance [110] and type 2 diabetes [111].
Study strengths
-
To our knowledge, this is the first trial to investigate the potential effects of gut bacterial modulation on advanced glycation.
-
Random allocation of participants to treatment sequence and intention to treat analysis will ensure study bias is minimised.
-
Double-blind crossover placebo-controlled trial.
-
Dietary data will be obtained to determine dietary patterns as well as estimates of total energy intake, fat and AGE consumption.
-
CML in this study will be measured using a validated ELISA method.
-
Dietary AGE consumption will be estimated from an open-source food AGE database, containing AGE values obtained using validated measurement techniques.
Study limitations
Type 2 diabetes is a chronic condition which often develops over decades, making it difficult to conduct intervention studies using the presence or absence of diabetes as the primary biological end-point. This study will measure surrogate biochemical markers of early type 2 diabetes pathogenesis, which could be considered a limitation of the trial. Long-term studies will need to be conducted in order to confirm the results of this research.
This study will measure serum CML as an indicator of AGE concentration in blood samples. Multiple other forms of AGEs exist, many of which have not yet been characterised, so the findings of this trial cannot be applied to all members of the AGE family. However, serum CML concentration shows a moderate to high correlation with other known circulating AGEs [39].
Applicability of research findings
In Australia, conservative estimates predict that at least 2 million adults will have been diagnosed with type 2 diabetes by 2025 [112]. The burden of disease associated with diabetes has a substantial impact on costs associated directly with health care as well as loss of productivity and decreased quality of life. The widespread consumption of high fat, heat processed foods and the increasing prevalence of obesity in Australia warrant simple interventions including those that prevent AGE-mediated damage. If dietary treatments aimed at altering the gut microbiota prove to be effective strategies for preventing or slowing the development of type 2 diabetes, they could become mainstream therapies for individuals with diabetes risk factors.
Abbreviations
- AGE:
-
Advanced glycation endproduct
- sRAGE:
-
Soluble receptor for advanced glycation endproducts
- HOMA:
-
Homeostasis model assessment
- TAC:
-
Total antioxidant capacity
- TNFα:
-
Tumour necrosis factor alpha
- IL-6:
-
Interleukin 6
- VCAM-1:
-
Vascular cell adhesion molecule 1
- RAGE:
-
Receptor for advanced glycation endproducts
- CML:
-
Carboxymethyl-lysine
- MG:
-
Methylglyoxal
- LPS:
-
Lipopolysaccharide
- SCFA:
-
Short chain fatty acid
- GPR:
-
G-protein coupled receptor
- PYY:
-
Peptide YY
- GLP-2:
-
Glucagon-like peptide 2
- ACE:
-
Angiotensin converting enzyme
- DPP-4:
-
Dipeptidyl peptidase 4
- Fiaf:
-
Fasting-induced adipocyte factor
- ELISA:
-
Enzyme linked immunosorbent assay
- NFκB:
-
Nuclear Factor kappa-B
- TLR4:
-
Toll-like receptor 4.
References
Henle T: Protein-bound advanced glycation endproducts (AGEs) as bioactive amino acid derivatives in foods. Amino Acids. 2005, 29 (4): 313-322.
Brownlee M: Biochemistry and molecular cell biology of diabetic complications. Nature. 2001, 414 (6865): 813-820.
Folli F, Corradi D, Fanti P, Davalli A, Paez A, Giaccari A, Perego C, Muscogiuri G: The role of oxidative stress in the pathogenesis of type 2 diabetes mellitus micro- and macrovascular complications: avenues for a mechanistic-based therapeutic approach. Curr Diabetes Rev. 2011, 7 (5): 313-324.
Kotani K, Sakane N: C-reactive protein and reactive oxygen metabolites in subjects with metabolic syndrome. J Int Med Res. 2012, 40 (3): 1074-1081.
Thornalley PJ: Dicarbonyl intermediates in the maillard reaction. Ann N Y Acad Sci. 2005, 1043: 111-117.
Avery NC, Bailey AJ: The effects of the Maillard reaction on the physical properties and cell interactions of collagen. Pathol Biol. 2006, 54 (7): 387-395.
Rosca MG, Mustata TG, Kinter MT, Ozdemir AM, Kern TS, Szweda LI, Brownlee M, Monnier VM, Weiss MF: Glycation of mitochondrial proteins from diabetic rat kidney is associated with excess superoxide formation. Am J Physiol Renal Physiol. 2005, 289 (2): F420-F430.
Coughlan MT, Thorburn DR, Penfold SA, Laskowski A, Harcourt BE, Sourris KC, Tan AL, Fukami K, Thallas-Bonke V, Nawroth PP, Brownlee M, Bierhaus A, Cooper ME, Forbes JM: RAGE-induced cytosolic ROS promote mitochondrial superoxide generation in diabetes. J Am Soc Nephrol. 2009, 20 (4): 742-752.
Yim MB, Yim HS, Lee C, Kang SO, Chock PB: Protein glycation: creation of catalytic sites for free radical generation. Ann N Y Acad Sci. 2001, 928: 48-53.
Schmidt AM, Hori O, Chen JX, Li JF, Crandall J, Zhang J, Cao R, Yan SD, Brett J, Stern D: Advanced glycation endproducts interacting with their endothelial receptor induce expression of vascular cell adhesion molecule-1 (VCAM-1) in cultured human endothelial cells and in mice: A potential mechanism for the accelerated vasculopathy of diabetes. J Clin Invest. 1995, 96 (3): 1395-1403.
Morita M, Yano S, Yamaguchi T, Sugimoto T: Advanced glycation end products-induced reactive oxygen species generation is partly through NF-kappa B activation in human aortic endothelial cells. J Diabetes Complications. 2013, 27 (1): 11-15.
Tahara N, Yamagishi S, Matsui T, Takeuchi M, Nitta Y, Kodama N, Mizoguchi M, Imaizumi T: Serum levels of advanced glycation end products (AGEs) are independent correlates of insulin resistance in nondiabetic subjects. Cardiovasc Ther. 2012, 30 (1): 42-48.
Tan KC, Shiu SW, Wong Y, Tam X: Serum advanced glycation end products (AGEs) are associated with insulin resistance. Diabetes Metab Res Rev. 2011, 27 (5): 488-492.
Uribarri J, Cai W, Sandu O, Peppa M, Goldberg T, Vlassara H: Diet-derived advanced glycation end products are major contributors to the body's AGE pool and induce inflammation in healthy subjects. Ann N Y Acad Sci. 2005, 1043: 461-466.
Fiory F, Lombardi A, Miele C, Giudicelli J, Beguinot F, Van Obberghen E: Methylglyoxal impairs insulin signalling and insulin action on glucose-induced insulin secretion in the pancreatic beta cell line INS-1E. Diabetologia. 2011, 54 (11): 2941-2952.
Coughlan MT, Yap FY, Tong DC, Andrikopoulos S, Gasser A, Thallas-Bonke V, Webster DE, Miyazaki J, Kay TW, Slattery RM, Kaye DM, Drew BG, Kingwell BA, Fourlanos S, Groop PH, Harrison LC, Knip M, Forbes JM: Advanced glycation end products are direct modulators of beta-cell function. Diabetes. 2011, 60 (10): 2523-2532.
Birlouez-Aragon I, Saavedra G, Tessier FJ, Galinier A, Ait-Ameur L, Lacoste F, Niamba CN, Alt N, Somoza V, Lecerf JM: A diet based on high-heat-treated foods promotes risk factors for diabetes mellitus and cardiovascular diseases. Am J Clin Nutr. 2010, 91 (5): 1220-1226.
Tessier FJ, Birlouez-Aragon I: Health effects of dietary Maillard reaction products: the results of ICARE and other studies. Amino Acids. 2012, 42 (4): 1119-1131.
Sandu O, Song K, Cai W, Zheng F, Uribarri J, Vlassara H: Insulin resistance and type 2 diabetes in high-fat-fed mice are linked to high glycotoxin intake. Diabetes. 2005, 54 (8): 2314-2319.
Cai W, Ramdas M, Zhu L, Chen X, Striker GE, Vlassara H: Oral advanced glycation endproducts (AGEs) promote insulin resistance and diabetes by depleting the antioxidant defenses AGE receptor-1 and sirtuin 1. Proc Natl Acad Sci U S A. 2012, 109 (39): 15888-15893.
Uribarri J, Cai W, Ramdas M, Goodman S, Pyzik R, Chen X, Zhu L, Striker GE, Vlassara H: Restriction of advanced glycation end products improves insulin resistance in human type 2 diabetes: potential role of AGER1 and SIRT1. Diabetes Care. 2011, 34 (7): 1610-1616.
Mark AB, Poulsen MW, Andersen S, Andersen JM, Bak MJ, Ritz C, Holst JJ, Nielsen J, de Courten B, Dragsted LO, Bugel S: Consumption of a diet low in advanced glycation endproducts for 4 weeks improves insulin sensitivity in overweight women. Diabetes Care. 2014, 37 (1): 88-95.
Luevano-Contreras C, Garay-Sevilla ME, Wrobel K, Malacara JM: Dietary advanced glycation end products restriction diminishes inflammation markers and oxidative stress in patients with type 2 diabetes mellitus. J Clin Biochem Nutr. 2013, 52 (1): 22-26.
Harcourt BE, Sourris KC, Coughlan MT, Walker KZ, Dougherty SL, Andrikopoulos S, Morley AL, Thallas-Bonke V, Chand V, Penfold SA, de Courten MP, Thomas MC, Kingwell BA, Bierhaus A, Cooper ME, Courten B, Forbes JM: Targeted reduction of advanced glycation improves renal function in obesity. Kidney Int. 2011, 80 (2): 190-198.
Kellow NJ, Savige GS: Dietary advanced glycation end-product restriction for the attenuation of insulin resistance, oxidative stress and endothelial dysfunction: a systematic review. Eur J Clin Nutr. 2013, 67 (3): 239-248.
Round JL, Mazmanian SK: The gut microbiota shapes intestinal immune responses during health and disease. Nat Rev Immunol. 2009, 9 (5): 313-323.
Ding S, Chi MM, Scull BP, Rigby R, Schwerbrock NM, Magness S, Jobin C, Lund PK: High-fat diet: bacteria interactions promote intestinal inflammation which precedes and correlates with obesity and insulin resistance in mouse. PLoS One. 2010, 5 (8): e12191-
Diamant M, Blaak EE, de Vos WM: Do nutrient-gut-microbiota interactions play a role in human obesity, insulin resistance and type 2 diabetes?. Obes Rev. 2011, 12 (4): 272-281.
Backhed F, Manchester JK, Semenkovich CF, Gordon JI: Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc Natl Acad Sci U S A. 2007, 104 (3): 979-984.
Hill MJ: Intestinal flora and endogenous vitamin synthesis. Eur J Cancer Prev. 1997, 6 (Suppl 1): S43-S45.
Macfarlane GT, Macfarlane S: Fermentation in the human large intestine: its physiologic consequences and the potential contribution of prebiotics. J Clin Gastroenterol. 2011, 45 (Suppl): S120-S127.
Wu X, Ma C, Han L, Nawaz M, Gao F, Zhang X, Yu P, Zhao C, Li L, Zhou A, Wang J, Moore JE, Millar BC, Xu J: Molecular characterisation of the faecal microbiota in patients with type II diabetes. Curr Microbiol. 2010, 61 (1): 69-78.
Larsen N, Vogensen FK, van den Berg FW, Nielsen DS, Andreasen AS, Pedersen BK, Al-Soud WA, Sorensen SJ, Hansen LH, Jakobsen M: Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS One. 2010, 5 (2): e9085-
Mills DJ, Tuohy KM, Booth J, Buck M, Crabbe MJ, Gibson GR, Ames JM: Dietary glycated protein modulates the colonic microbiota towards a more detrimental composition in ulcerative colitis patients and non-ulcerative colitis subjects. J Appl Microbiol. 2008, 105 (3): 706-714.
Nakamura YK, Omaye ST: Metabolic diseases and pro- and prebiotics: mechanistic insights. Nutr Metab (Lond). 2012, 9 (1): 60-
Quigley EM: Therapies aimed at the gut microbiota and inflammation: antibiotics, prebiotics, probiotics, synbiotics, anti-inflammatory therapies. Gastroenterol Clin North Am. 2011, 40 (1): 207-222.
Delzenne NM, Neyrinck AM, Backhed F, Cani PD: Targeting gut microbiota in obesity: effects of prebiotics and probiotics. Nat Rev Endocrinol. 2011, 7 (11): 639-646.
Colagiuri SDD, Girgis S, Colagiuri R: National evidence based guideline for case detection and diagnosis of type 2 diabetes. 2009, Canberra, Australia: Diabetes Australia and the National Health and Medical Research Council
Semba RD, Arab L, Sun K, Nicklett EJ, Ferrucci L: Fat mass is inversely associated with serum carboxymethyl-lysine, an advanced glycation end product, in adults. J Nutr. 2011, 141 (9): 1726-1730.
Kilhovd BK, Juutilainen A, Lehto S, Ronnemaa T, Torjesen PA, Hanssen KF, Laakso M: Increased serum levels of methylglyoxal-derived hydroimidazolone-AGE are associated with increased cardiovascular disease mortality in nondiabetic women. Atherosclerosis. 2009, 205 (2): 590-594.
Lapolla A, Piarulli F, Sartore G, Ceriello A, Ragazzi E, Reitano R, Baccarin L, Laverda B, Fedele D: Advanced glycation end products and antioxidant status in type 2 diabetic patients with and without peripheral artery disease. Diabetes Care. 2007, 30 (3): 670-676.
Semba RD, Bandinelli S, Sun K, Guralnik JM, Ferrucci L: Plasma carboxymethyl-lysine, an advanced glycation end product, and all-cause and cardiovascular disease mortality in older community-dwelling adults. J Am Geriatr Soc. 2009, 57 (10): 1874-1880.
Kolida S, Meyer D, Gibson GR: A double-blind placebo-controlled study to establish the bifidogenic dose of inulin in healthy humans. Eur J Clin Nutr. 2007, 61 (10): 1189-1195.
Causey JL: Effects of dietary inulin on serum lipids, blood glucose and the gastrointestinal environment in hypercholesterolemic men. Nutr Res. 2000, 20 (2): 191-201.
Lecerf JM, Depeint F, Clerc E, Dugenet Y, Niamba CN, Rhazi L, Cayzeele A, Abdelnour G, Jaruga A, Younes H, Jacobs H, Lambrey G, Abdelnour AM, Pouillart PR: Xylo-oligosaccharide (XOS) in combination with inulin modulates both the intestinal environment and immune status in healthy subjects, while XOS alone only shows prebiotic properties. Br J Nutr. 2012, 108 (10): 1847-1858.
Hull GLJ, Ames JM, Cuskelly GJ: NEpsilon-(carboxymethyl)lysine content of foods commonly consumed in a Western style diet. Food Chem. 2012, 131: 170-174.
Bihuniak JD, Simpson CA, Sullivan RR, Caseria DM, Kerstetter JE, Insogna KL: Dietary protein-induced increases in urinary calcium are accompanied by similar increases in urinary nitrogen and urinary urea: a controlled clinical trial. J Acad Nutr Diet. 2013, 113 (3): 447-451.
Craig CL, Marshall AL, Sjostrom M, Bauman AE, Booth ML, Ainsworth BE, Pratt M, Ekelund U, Yngve A, Sallis JF, Oja P: International physical activity questionnaire: 12-country reliability and validity. Med Sci Sports Exerc. 2003, 35 (8): 1381-1395.
Boehm BO, Schilling S, Rosinger S, Lang GE, Lang GK, Kientsch-Engel R, Stahl P: Elevated serum levels of N(epsilon)-carboxymethyl-lysine, an advanced glycation end product, are associated with proliferative diabetic retinopathy and macular oedema. Diabetologia. 2004, 47 (8): 1376-1379.
Zhang X, Frischmann M, Kientsch-Engel R, Steinmann K, Stopper H, Niwa T, Pischetsrieder M: Two immunochemical assays to measure advanced glycation end-products in serum from dialysis patients. Clin Chem Lab Med. 2005, 43 (5): 503-511.
Matthews DR, Hosker JP, Rudenski AS, Naylor BA, Treacher DF, Turner RC: Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia. 1985, 28 (7): 412-419.
Bonora E, Targher G, Alberiche M, Bonadonna RC, Saggiani F, Zenere MB, Monauni T, Muggeo M: Homeostasis model assessment closely mirrors the glucose clamp technique in the assessment of insulin sensitivity: studies in subjects with various degrees of glucose tolerance and insulin sensitivity. Diabetes Care. 2000, 23 (1): 57-63.
Matsuki T, Watanabe K, Fujimoto J, Miyamoto Y, Takada T, Matsumoto K, Oyaizu H, Tanaka R: Development of 16S rRNA-gene-targeted group-specific primers for the detection and identification of predominant bacteria in human feces. Appl Environ Microbiol. 2002, 68 (11): 5445-5451.
Dewulf EM, Cani PD, Claus SP, Fuentes S, Puylaert PG, Neyrinck AM, Bindels LB, de Vos WM, Gibson GR, Thissen JP, Delzenne NM: Insight into the prebiotic concept: lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut. 2013, 62 (8): 1112-1121.
Ramirez-Farias C, Slezak K, Fuller Z, Duncan A, Holtrop G, Louis P: Effect of inulin on the human gut microbiota: stimulation of Bifidobacterium adolescentis and Faecalibacterium prausnitzii. Br J Nutr. 2009, 101 (4): 541-550.
Wang RF, Cao WW, Cerniglia CE: PCR detection and quantitation of predominant anaerobic bacteria in human and animal fecal samples. Appl Environ Microbiol. 1996, 62 (4): 1242-1247.
Everard A, Belzer C, Geurts L, Ouwerkerk JP, Druart C, Bindels LB, Guiot Y, Derrien M, Muccioli GG, Delzenne NM, de Vos WM, Cani PD: Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci U S A. 2013, 110 (22): 9066-9071.
Sebekova K, Boor P, Valachovicova M, Blazicek P, Parrak V, Babinska K, Heidland A, Krajcovicova-Kudlackova M: Association of metabolic syndrome risk factors with selected markers of oxidative status and microinflammation in healthy omnivores and vegetarians. Mol Nutr Food Res. 2006, 50 (9): 858-868.
Vlassara H, Uribarri J, Cai W, Striker G: Advanced glycation end product homeostasis: exogenous oxidants and innate defenses. Ann N Y Acad Sci. 2008, 1126: 46-52.
Goldberg T, Cai W, Peppa M, Dardaine V, Baliga BS, Uribarri J, Vlassara H: Advanced glycoxidation end products in commonly consumed foods. J Am Diet Assoc. 2004, 104 (8): 1287-1291.
Shen Q, Chen YA, Tuohy KM: A comparative in vitro investigation into the effects of cooked meats on the human faecal microbiota. Anaerobe. 2010, 16 (6): 572-577.
Connolly ML, Lovegrove JA, Tuohy KM: In vitro fermentation characteristics of whole grain wheat flakes and the effect of toasting on prebiotic potential. J Med Food. 2012, 15 (1): 33-43.
De Preter V, Hamer HM, Windey K, Verbeke K: The impact of pre- and/or probiotics on human colonic metabolism: does it affect human health?. Mol Nutr Food Res. 2011, 55 (1): 46-57.
Faist V, Erbersdobler HF: Metabolic transit and in vivo effects of melanoidins and precursor compounds deriving from the Maillard reaction. Ann Nutr Metab. 2001, 45 (1): 1-12.
Rapin JR, Wiernsperger N: Possible links between intestinal permeability and food processing: A potential therapeutic niche for glutamine. Clinics (Sao Paulo). 2010, 65 (6): 635-643.
Scheppach W: Effects of short chain fatty acids on gut morphology and function. Gut. 1994, 35 (1 Suppl): S35-S38.
Eun CS, Han DS, Lee SH, Paik CH, Chung YW, Lee J, Hahm JS: Attenuation of colonic inflammation by PPARgamma in intestinal epithelial cells: effect on Toll-like receptor pathway. Dig Dis Sci. 2006, 51 (4): 693-697.
Canani RB, Costanzo MD, Leone L, Pedata M, Meli R, Calignano A: Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J Gastroenterol. 2011, 17 (12): 1519-1528.
Delzenne NM, Cani PD: Interaction between obesity and the gut microbiota: relevance in nutrition. Annu Rev Nutr. 2011, 31: 15-31.
Russo F, Linsalata M, Clemente C, Chiloiro M, Orlando A, Marconi E, Chimienti G, Riezzo G: Inulin-enriched pasta improves intestinal permeability and modifies the circulating levels of zonulin and glucagon-like peptide 2 in healthy young volunteers. Nutr Res. 2012, 32 (12): 940-946.
Cani PD, Possemiers S, Van de Wiele T, Guiot Y, Everard A, Rottier O, Geurts L, Naslain D, Neyrinck A, Lambert DM, Muccioli GG, Delzenne NM: Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut. 2009, 58 (8): 1091-1103.
Cani PD: Crosstalk between the gut microbiota and the endocannabinoid system: impact on the gut barrier function and the adipose tissue. Clin Microbiol Infect. 2012, 18 (Suppl 4): 50-53.
Schalkwijk CG, Miyata T: Early- and advanced non-enzymatic glycation in diabetic vascular complications: the search for therapeutics. Amino Acids. 2012, 42 (4): 1193-1204.
Anderson MM, Heinecke JW: Production of N(epsilon)-(carboxymethyl)lysine is impaired in mice deficient in NADPH oxidase: a role for phagocyte-derived oxidants in the formation of advanced glycation end products during inflammation. Diabetes. 2003, 52 (8): 2137-2143.
Huebschmann AG, Regensteiner JG, Vlassara H, Reusch JE: Diabetes and advanced glycoxidation end products. Diabetes Care. 2006, 29 (6): 1420-1432.
Hildebrandt MA, Hoffmann C, Sherrill-Mix SA, Keilbaugh SA, Hamady M, Chen YY, Knight R, Ahima RS, Bushman F, Wu GD: High-fat diet determines the composition of the murine gut microbiome independently of obesity. Gastroenterology. 2009, 137 (5): 1716-1724. e1711-1712
Vrieze A, Holleman F, Zoetendal EG, de Vos WM, Hoekstra JB, Nieuwdorp M: The environment within: how gut microbiota may influence metabolism and body composition. Diabetologia. 2010, 53 (4): 606-613.
Esteve E, Ricart W, Fernandez-Real JM: Gut microbiota interactions with obesity, insulin resistance and type 2 diabetes: did gut microbiote co-evolve with insulin resistance?. Curr Opin Clin Nutr Metab Care. 2011, 14 (5): 483-490.
Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, Neyrinck AM, Fava F, Tuohy KM, Chabo C, Waget A, Delmee E, Cousin B, Sulpice T, Chamontin B, Ferrieres J, Tanti JF, Gibson GR, Casteilla L, Delzenne NM, Alessi MC, Burcelin R: Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. 2007, 56 (7): 1761-1772.
Priebe MG, Wang H, Weening D, Schepers M, Preston T, Vonk RJ: Factors related to colonic fermentation of nondigestible carbohydrates of a previous evening meal increase tissue glucose uptake and moderate glucose-associated inflammation. Am J Clin Nutr. 2010, 91 (1): 90-97.
Creely SJ, McTernan PG, Kusminski CM, Fisher FM, Da Silva NF, Khanolkar M, Evans M, Harte AL, Kumar S: Lipopolysaccharide activates an innate immune system response in human adipose tissue in obesity and type 2 diabetes. Am J Physiol Endocrinol Metab. 2007, 292 (3): E740-E747.
Amar J: Energy intake is associated with endotoxemia induces adipose inflammation and insulin resistance in humans. Diabetes. 2010, 59: 9-
Cani PD, Neyrinck AM, Fava F, Knauf C, Burcelin RG, Tuohy KM, Gibson GR, Delzenne NM: Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia. 2007, 50 (11): 2374-2383.
Lin MY, Yen CL: Inhibition of lipid peroxidation by Lactobacillus acidophilus and Bifidobacterium longum. J Agric Food Chem. 1999, 47 (9): 3661-3664.
Yen CH, Kuo YW, Tseng YH, Lee MC, Chen HL: Beneficial effects of fructo-oligosaccharides supplementation on fecal bifidobacteria and index of peroxidation status in constipated nursing-home residents–a placebo-controlled, diet-controlled trial. Nutrition. 2011, 27 (3): 323-328.
Maslowski KM, Vieira AT, Ng A, Kranich J, Sierro F, Yu D, Schilter HC, Rolph MS, Mackay F, Artis D, Xavier RJ, Teixeira MM, Mackay CR: Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature. 2009, 461 (7268): 1282-1286.
Vinolo MA, Rodrigues HG, Nachbar RT, Curi R: Regulation of inflammation by short chain fatty acids. Nutrients. 2011, 3 (10): 858-876.
Lewis K, Lutgendorff F, Phan V, Soderholm JD, Sherman PM, McKay DM: Enhanced translocation of bacteria across metabolically stressed epithelia is reduced by butyrate. Inflamm Bowel Dis. 2010, 16 (7): 1138-1148.
Erridge C, Attina T, Spickett CM, Webb DJ: A high-fat meal induces low-grade endotoxemia: evidence of a novel mechanism of postprandial inflammation. Am J Clin Nutr. 2007, 86 (5): 1286-1292.
Krogh-Madsen R, Plomgaard P, Akerstrom T, Moller K, Schmitz O, Pedersen BK: Effect of short-term intralipid infusion on the immune response during low-dose endotoxemia in humans. Am J Physiol Endocrinol Metab. 2008, 294 (2): E371-E379.
Macia L, Thorburn AN, Binge LC, Marino E, Rogers KE, Maslowski KM, Vieira AT, Kranich J, Mackay CR: Microbial influences on epithelial integrity and immune function as a basis for inflammatory diseases. Immunol Rev. 2012, 245 (1): 164-176.
Maslowski KM, Mackay CR: Diet, gut microbiota and immune responses. Nat Immunol. 2011, 12 (1): 5-9.
Musso G, Gambino R, Cassader M: Interactions between gut microbiota and host metabolism predisposing to obesity and diabetes. Annu Rev Med. 2011, 62: 361-380.
Gugliucci A, Kotani K, Taing J, Matsuoka Y, Sano Y, Yoshimura M, Egawa K, Horikawa C, Kitagawa Y, Kiso Y, Kimura S, Sakane N: Short-term low calorie diet intervention reduces serum advanced glycation end products in healthy overweight or obese adults. Ann Nutr Metab. 2009, 54 (3): 197-201.
Cani PD, Joly E, Horsmans Y, Delzenne NM: Oligofructose promotes satiety in healthy human: a pilot study. Eur J Clin Nutr. 2006, 60 (5): 567-572.
Parnell JA, Reimer RA: Weight loss during oligofructose supplementation is associated with decreased ghrelin and increased peptide YY in overweight and obese adults. Am J Clin Nutr. 2009, 89 (6): 1751-1759.
Ruijschop RBA, te Giffela MC: Satiety effects of a dairy beverage fermented with propionic acid bacteria. Int Dairy J. 2008, 18: 5-
Backhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, Semenkovich CF, Gordon JI: The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci U S A. 2004, 101 (44): 15718-15723.
Aronsson L, Huang Y, Parini P, Korach-Andre M, Hakansson J, Gustafsson JA, Pettersson S, Arulampalam V, Rafter J: Decreased fat storage by Lactobacillus paracasei is associated with increased levels of angiopoietin-like 4 protein (ANGPTL4). PLoS One. 2010, 5 (9): doi:10.1371/journal.pone.0013087
Dewulf EM, Cani PD, Neyrinck AM, Possemiers S, Van Holle A, Muccioli GG, Deldicque L, Bindels LB, Pachikian BD, Sohet FM, Mignolet E, Francaux M, Larondelle Y, Delzenne NM: Inulin-type fructans with prebiotic properties counteract GPR43 overexpression and PPARgamma-related adipogenesis in the white adipose tissue of high-fat diet-fed mice. J Nutr Biochem. 2011, 22 (8): 712-722.
Vyas U, Ranganathan N: Probiotics, prebiotics, and synbiotics: gut and beyond. Gastroenterology Res Pract. 2012, 2012: 872716-
Mehta R, Shangari N, O'Brien PJ: Preventing cell death induced by carbonyl stress, oxidative stress or mitochondrial toxins with vitamin B anti-AGE agents. Mol Nutr Food Res. 2008, 52 (3): 379-385.
Stoyanova S, Geuns J, Hideg E, Van Den Ende W: The food additives inulin and stevioside counteract oxidative stress. Int J Food Sci Nutr. 2011, 62 (3): 207-214.
Wu WT, Chen HL: Effects of konjac glucomannan on putative risk factors for colon carcinogenesis in rats fed a high-fat diet. J Agric Food Chem. 2011, 59 (3): 989-994.
Todesco T, Rao AV, Bosello O, Jenkins DJ: Propionate lowers blood glucose and alters lipid metabolism in healthy subjects. Am J Clin Nutr. 1991, 54 (5): 860-865.
Gao Z, Yin J, Zhang J, Ward RE, Martin RJ, Lefevre M, Cefalu WT, Ye J: Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes. 2009, 58 (7): 1509-1517.
Cani PD, Hoste S, Guiot Y, Delzenne NM: Dietary non-digestible carbohydrates promote L-cell differentiation in the proximal colon of rats. Br J Nutr. 2007, 98 (1): 32-37.
Cani PD, Knauf C, Iglesias MA, Drucker DJ, Delzenne NM, Burcelin R: Improvement of glucose tolerance and hepatic insulin sensitivity by oligofructose requires a functional glucagon-like peptide 1 receptor. Diabetes. 2006, 55 (5): 1484-1490.
Uchiki T, Weikel KA, Jiao W, Shang F, Caceres A, Pawlak D, Handa JT, Brownlee M, Nagaraj R, Taylor A: Glycation-altered proteolysis as a pathobiologic mechanism that links dietary glycemic index, aging, and age-related disease (in nondiabetics). Aging Cell. 2012, 11 (1): 1-13.
Garcia AL, Otto B, Reich SC, Weickert MO, Steiniger J, Machowetz A, Rudovich NN, Mohlig M, Katz N, Speth M, Meuser F, Doerfer J, Zunft HJ, Pfeiffer AH, Koebnick C: Arabinoxylan consumption decreases postprandial serum glucose, serum insulin and plasma total ghrelin response in subjects with impaired glucose tolerance. Eur J Clin Nutr. 2007, 61 (3): 334-341.
Lu ZX, Walker KZ, Muir JG, O'Dea K: Arabinoxylan fibre improves metabolic control in people with Type II diabetes. Eur J Clin Nutr. 2004, 58 (4): 621-628.
Magliano DJ, Peeters A, Vos T, Sicree R, Shaw J, Sindall C, Haby M, Begg SJ, Zimmet PZ: Projecting the burden of diabetes in Australia–what is the size of the matter?. Aust N Z J Public Health. 2009, 33 (6): 540-543.
Pre-publication history
The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1472-6823/14/55/prepub
Acknowledgements
The authors thank Associate Professor Rory Wolfe (SPHPM, Monash University) for advice regarding biostatistical analyses. NJ Kellow is the recipient of a National Health and Medical Research Council (NHMRC) Postgraduate Public Health Scholarship APP1039709. MT Coughlan is supported by an Australian and New Zealand Society of Nephrology Early Career Fellowship.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
NJK and MTC designed the study, NJK drafted the manuscript with assistance from MTC, GSS and CMR. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Kellow, N.J., Coughlan, M.T., Savige, G.S. et al. Effect of dietary prebiotic supplementation on advanced glycation, insulin resistance and inflammatory biomarkers in adults with pre-diabetes: a study protocol for a double-blind placebo-controlled randomised crossover clinical trial. BMC Endocr Disord 14, 55 (2014). https://doi.org/10.1186/1472-6823-14-55
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1472-6823-14-55