
Abstract
Background
Natural hybridization is prevalent in ferns, and plays an important role in fern evolution and speciation. In the Indo West-Pacific region, the mangrove fern genus Acrostichum consists of two largely sympatric species, A. aureum and A. speciosum. Although there has been no report of interspecific hybridization before, we found some individuals morphologically intermediate between them in Guangdong and Hainan, China, for the first time, which were suspected to be hybrids. In this study, we aimed to test the hypothesis of natural hybridization between A. aureum and A. speciosum in Guangdong and Hainan using three low-copy nuclear genes. A chloroplast intergenic spacer was used to infer the hybridization direction once the hybrid status was confirmed. In addition, we examined spore shapes and germination for these taxa.
Results
Both A. aureum and A. speciosum showed a low level of polymorphism at all three nuclear genes; however, they were well separated at these loci. At both locations, each individual of the putative hybrid showed additivity in chromatograms at all sites where the two species showed fixed differences. Haplotype analysis at all three nuclear genes indicated that each individual of the putative hybrid possessed two haplotypes, matching with those of A. aureum and A. speciosum, respectively. Sequencing of the chloroplast trnV-trnM regions showed that A. aureum differed from A. speciosum by eleven nucleotide substitutions and three indels (insertions/deletions), and all sampled individuals of the putative hybrid had the identical sequences with A. speciosum. Compared with A. aureum and A. speciosum, the putative hybrid had much reduced spore germination rate.
Conclusions
Sequence data of the three nuclear genes provide compelling evidence for natural hybridization between A. aureum and A. speciosum, and all the hybrid individuals are likely F1s. The hybridization is unidirectional and A. speciosum is the maternal parent of the hybrid based on the assumption of maternal inheritance of chloroplast DNA. Human disturbance on mangrove habitats may facilitate the establishment of hybrids of Acrostichum.
Similar content being viewed by others


Background
Natural hybridization is a common phenomenon in plants and has a significant role in plant evolution [1, 2]. Depending on the interaction of intrinsic and extrinsic factors, interspecific hybridization can lead to formation of stable genetic lineages in the forms of polyploid or homoploid hybrid species (reviewed in [3]), reinforcement of premating isolation as a response to maladaptive hybrids [4], and gene transfer through genetic recombination or introgression [5–8]. Hybridization occurs more prevalent in ferns than flowering plants and a wide spectrum of fern species are found to be derived from interspecific hybridization [2, 9–13]. In the past several decades, many cases of natural hybridization in ferns have been identified (e.g. [10, 14–18]). In this study, we will focus on natural hybridization between two species in the mangrove fern genus Acrostichum.
Consisting of about 70 species from about 20 families [19], mangroves are a special group of plants that inhabit the intertidal zones of tropical and subtropical coasts. Acrostichum L., the only fern genus of mangroves, comprises three species, namely, A. aureum L., A. speciosum Willd. and A. danaeifolium Langsd. & Fish [20]. Among the three species, A. speciosum is restricted in the Indo West-Pacific (IWP) region and A. danaeifolium is confined to the Atlantic East-Pacific (AEP) region, while A. aureum is widely distributed in both regions [20, 21]. Species of Acrostichum are usually called mangrove ferns.
In China, A. aureum is a common species, ranging from Hainan through Guangdong and Guangxi to Fujian and Taiwan. In contrast, A. speciosum is a rare species, occurring in only two locations, Wenchang, Hainan and Zhanjiang, Guangdong (W. Wang, unpublished data). Both A. aureum and A. speciosum occur on the landward edge of mangrove swamps, however, they show different ecological preferences to light and salinity. A. aureum is often encountered as a river or creek plant where fresh water influence is strong [20, 22, 23]. It also behaves like a light demanding fern, and often can be found in deteriorated or disturbed mangrove areas [24, 25]. In contrast, A. speciosum is usually found at the mangrove understory, and just at the margins of high intertidal zones. These areas are frequently inundated by tides and usually are shady [20, 26]. In fields, the two species are easily distinguished based on frond shape and texture. The frond of A. speciosum narrows gradually to a pointed tip, while that of A. aureum has a broadly rounded end (Figure 1). Additionally, the frond of A. aureum is thickly coriaceous, while that of A. speciosum is papery. It seems that the textures of the two species conform to their ecological preferences to light.

Morphological illustrations for A. aureum (left), the putative hybrid (middle), and A. speciosum (right).
During our field trips in Wenchang, Hainan and Zhanjiang, Guangdong, where the two species coexist, we found some individuals morphologically intermediate between A. aureum and A. speciosum. These individuals have leathery fronds similar to A. aureum, and gradually narrowed and pointed tips like A. speciosum. Most, if not all, individuals were found very close to the shrimp ponds, where the natural mangrove habitats have been seriously disturbed or destroyed. In China, we haven’t found any individuals of this kind in other areas where only A. aureum occurs. With intermediate morphology and overlapping geographic distribution, these individuals are suspected to be interspecific hybrids between A. aureum and A. speciosum.
Because plants often show a high degree of morphological plasticity in response to various environmental factors, and the intermediate morphology can arise from forces other than hybridization, morphological criteria alone for identifying natural hybrids are inadequate. Conventional approaches such as morphological analysis, chromosome number and pairing behavior, and allozyme assay have been widely used to study hybridization in ferns (e.g. [11, 15, 27, 28], these methods, however, either fail to provide convincing conclusions or are time and labor consuming, especially when the sample size is large. Recently, single or low-copy nuclear genes have proven successful in identifying hybrids in plants [14, 29–35]. In this study, we aimed to test the hypothesis of natural hybridization between A. aureum and A. speciosum using the sequences of three low-copy nuclear genes. Once the hybrid status was confirmed, a chloroplast DNA fragment was used to determine the maternal parent of these hybrid individuals, because chloroplast DNA is usually maternally transmitted in ferns [36]. In addition, we examined the spore shapes and germination rates for the three taxa to see if the putative hybrid has reduced fertility relative to its putative parents.
Results
At the three nuclear genes, almost all samples of A. aureum and A. speciosum, and most samples of the putative hybrid, could be directly sequenced and clear sequences were obtained. For each gene, both A. aureum and A. speciosum exhibited limited sequence variation (see below for details). However, these two species were highly diverged at all three nuclear genes and one chloroplast DNA fragment.
Sequences analysis of cam genes in A. aureum, A. speciosumand the putative hybrid
The partial sequences of the cam gene we analyzed were 453 bp in length in the three taxa of Acrostichum. There were nine fixed nucleotide substitutions between A. aureum and A. speciosum. For the putative hybrid, all individuals showed chromatogram additivity at these nine fixed sites between A. aureum and A. speciosum. Both species showed a low level of haplotype diversity at the cam gene, with two haplotypes being observed in each species (Figure 2A). All but the Danzhou population of A. aureum were monomorphic and shared the common haplotype aA1, and the other haplotype aA2 differed from haplotype aA1 by one mutation (Figure 3A). Both populations of A. speciosum possessed haplotypes sA1 and sA2, although the frequencies of the two haplotypes were different between them (Figure 3A). For the putative hybrid, seven individuals from Wenchang and nine individuals from Suixi showed a combination of haplotype aA1 of A. aureum and haplotype sA1 of A. speciosum (Table 1; Additional file 1: Table S1). Haplotypes of the remaining six individuals were a combination of haplotype aA1 of A. aureum and haplotype sA2 of A. speciosum (Table 1).

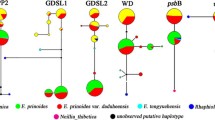
Haplotype networks of cam (A), gapCp1 (B) and gapCp2 (C) genes for A. speciosum and A. aureum . Color circles represent haplotypes of A. aureum, and black, white and grey squares represent haplotypes of A. speciosum. a and s in the haplotype names mean A. aureum and A. speciosum, respectively. Small black circles represent hypothetical or unsampled haplotypes. Mutational steps are shown by the length of the connecting lines.

Haplotype distribution of the three nuclear genes cam (A), gapCp1 (B) and gapCp2 (C) in each population of A. aureum and A. speciosum . Color codes correspond to those in Figure 2.
Sequences analysis of gapCp1 genes in A. aureum, A. speciosumand the putative hybrid
The partial gapCp1 genes were 601 bp in the three taxa after sequence alignment. A. aureum and A. speciosum differed in eleven fixed nucleotide substitutions and one fixed 5-bp indel (insertion/deletion). All individuals of the putative hybrid showed chromatogram additivity at these eleven fixed sites and one indel region. Six haplotypes were identified among all samples of A. aureum, and haplotype aB1 was dominant across all populations (Figure 2B). Only Huizhou population was monomorphic, and the number of haplotypes varied from two to four in other populations (Figure 3B). In contrast, A. speciosum exhibited a much lower level of haplotype diversity, with only two haplotypes being detected (Figure 2B). Wenchang population had two haplotypes (sB1 and sB2), while Suixi population had only one haplotype sB1 (Figure 3B). For the putative hybrid from Wenchang, six individuals showed a combination of haplotype aB1 of A. aureum and sB1 of A. speciosum, two individuals showed a combination of haplotype aB1 of A. aureum and haplotype sB2 of A. speciosum, one individual showed a combination of haplotype aB2 of A .aureum and haplotype sB1of A. speciosum, and one individual showed a combination of haplotype aB2 of A. aureum and haplotype sB2 of A. speciosum (Table 1; Additional file 1: Table S1). For the putative hybrid from Suixi, eight individuals showed a mix of haplotype aB1 of A. aureum and haplotype sB1 of A. speciosum, and haplotypes of the remaining four individuals were a combination of haplotype sB1 of A. speciosum and a haplotype unique to the putative hybrid, which differed by one mutation from the common haplotype aB1 of A. aureum (Table 1).
Sequence analysis of gapCp2 genes in A. aureum, A. speciosumand the putative hybrid
The aligned length of gapCp2 genes in the three taxa was 490 bp. There were seven fixed nucleotide substitutions between A. aureum and A. speciosum. For the putative hybrid, all individuals showed chromatogram additivity at these seven fixed sites. In the haplotype analysis, six and three haplotypes were detected for A. aureum and A. speciosum, respectively (Figure 2C). The number of haplotypes varied from one to three in the populations of A. aureum and haplotype aC1 was dominant across all populations (Figure 2C, Figure 3C). In A. speciosum, three haplotypes (sC1, sC2 and sC3) were shared by the two populations (Figure 3C). For the putative hybrid from both locations, haplotypes of all but one individual were a combination of haplotype aC1 of A. aureum and haplotype sC1 or sC2 of A. speciosum (Table 1; Additional file 1: Table S1). Haplotypes of the special individual were a combination of haplotype sC2 and a unique haplotype to the putative hybrid, which differed by one mutation from the common hyplotype aC1 of A. aureum (Table 1).
Sequence analysis of chloroplast trnV-trnM in A. aureum, A. speciosumand the putative hybrid
The chloroplast trnV-trnM sequences of A. aureum and A. speciosum were both 975 bp in length. No sequence variation was detected within each species. There were eleven nucleotide substitutions and three indels in this region between A. aureum and A. speciosum. All sampled individuals of the putative hybrid from Wenchang and Suixi had the same trnV-trnM sequences as A. speciosum.
Spore morphology and germination of A. aureum, A. speciosumand the putative hybrid
The spores of A. aureum, A. speciosum and the putative hybrid were all globose and similar in size (diameter: 40–55 μm). However, the putative hybrid showed a much lower spore germination rate (20.3%) compared with A. aureum (79.8%) and A. speciosum (65.4%).
Discussion
Genetic variation in A. aureum and A. speciosum
Both A. aureum and A. speciosum are widespread in the Indo West-Pacific region; in China, however, A. aureum is much more common than A. speciosum. Our results indicate that both species harbor a low level of polymorphism at the three nuclear genes and a chloroplast intergenic spacer. Many populations have only one or two haplotypes at all of the loci examined, suggesting severe diversity loss or recent colonization in these species. It is well recognized that repeated population bottlenecks due to fluctuation of sea levels in the Pleistocene glaciation caused diversity loss in mangrove species [37]. Low levels of polymorphism in the two mangrove ferns are consistent with this historical demography. However, A. aureum and A. speciosum are well separated based on network analysis of the three nuclear genes and sequence divergence at the chloroplast fragment, indicating that the two species have diverged for a long time.
Molecular identification of natural hybridization between A. aureum and A. speciosum
Although A. aureum and A. speciosum overlap largely in geographic distribution in the Indo West-Pacific region, there has been no report of natural hybridization between them before. In this study, we used three low-copy nuclear genes to test the hypothesis of natural hybridization between them. Multiple individuals of A. aureum and A. speciosum, as well as their putative hybrid, have been used for this purpose. Although one haplotype unique to the putative hybrid is observed at each of the gapCp1 and gapCp2 genes, it is very close to the common haplotype of A. aureum (one mutational step). Unsampled polymorphisms in A. aureum may cause this pattern. This finding is not surprising given comparatively high diversity at these two genes in A. aureum. All individuals of the putative hybrid show perfect chromatogram additivity at each of the three nuclear genes, providing convincing evidence for natural hybridization between A. aureum and A. speciosum. The much reduced spore germination rate in the putative hybrid provides another line of evidence for this hypothesis. Moreover, these hybrid individuals are all likely F1s, because all of them show biparental inheritance at all three randomly selected nuclear genes. At the gapCp1 gene, haplotype sB2 of A. speciosum is unique to Wenchang population, and hybrids with this haplotype are found in only that location, suggesting the hybridization should arise locally. Given chloroplast DNA is predominantly maternally inherited in ferns [36], the chloroplast data indicate that A. speciosum might be the maternal parent of these hybrid individuals and the hybridization was unidirectional. Unidirectional hybridization in Acrostichum may owe to differential abundance of parental species. At both sites we studied, A. aureum is much more abundant than A. speciosum. A. aureum may thus produce much more mobile sperms, which have more chances to fertilize the egg of A. speciosum. In addition, strong prezygotic and (or) postzygotic barriers in the other direction (e.g. incompatibility of foreign gamete) and selection pressures from the environments on hybrid progeny may be other possibilities [6, 16, 18, 38, 39].
Factors contributing to natural hybridization between A. aureum and A. speciosum
In ferns, meiosis results in spores which give rise to gametophytes, and just in this situation sexual reproduction can occur [40]. The tiny and short-lived gametophytic generation plays an important role in the reproduction and dispersal ecology of ferns [41]. There is considerable overlap in geographic distribution between A. aureum and A. speciosum, especially in Southeast Asia and North Australia [19, 20]. Although these two species have distinctive habitat preferences, the spores of both species can be dispersed by sea currents [20] and their gametophytes may come into contact as a consequence of spore dispersal. Zygotes may thus establish via the fusion of gametes respectively originating from gametophytes of these two species, providing spatial chances for hybridization.
The consequences of hybridization between A. aureum and A. speciosum
Despite frequent occurrence of hybridization in Acrostichum, it seems that all hybrid individuals we sampled are simple F1s. The restriction of hybrids to F1s appears to be a general phenomenon in ferns [9, 42], and interspecific hybrids are sterile mostly because chromosomes do not behave normally in the process of spore production, resulting in abortive spores [14, 42]. In this study, although the hybrid is very similar to its parental species in spore shape and size, its spore germination rate is much reduced. Post-germination selection may further cause the absence of later-generation hybrids. The persistence of only sterile F1 hybrids is guaranteed by recurrent hybridization, which is to some extent in accord with that the hybrid is only found in places where its parental species overlap.
It is well known that hybridization followed by polyploidy is important in fern evolution and speciation [10, 43–45]. Sterile F1 hybrids can obtain the reproductive capacity through genome doubling [10, 45]. The chromosome number of these two Acrostichum species are both 2n=60 [46, 47]. So far, we have no data on chromosome number of these hybrid individuals, so further survey on this is deserved.
It is widely believed that endogenous and exogenous factors both play important roles in selection against hybrids [2, 5, 48]. We have not found any hybrids of Acrostichum in typical habitats of both parental species, implying selection against hybrids in the habitats of parental species. The hybrids, however, are frequently observed in the intermediate environments, for example, newly deforested sites for fish ponds, suggesting higher fitness of these hybrids in this kind of habitat. Hence, human disturbance on mangrove habitats may facilitate the establishment of hybrids of Acrostichum.
Growing at the interface between land and sea, mangroves are of great ecological significance [49, 50]. They protect and stabilize coastlines, yield commercial forest products, furnish nursery and spawning areas for commercially important coastal fishes and so on [49, 51]. Acrostichum, the only fern genus of mangroves, is one of the important mangrove components in both the IWP and AEP regions. A. aureum is a pioneer species of mangroves and it grows rapidly after mangrove forests have been destroyed and forms extensive dense thickets of about 3 to 4 m height, particularly in the more elevated inland areas less frequently being inundated by tides [52]. A. speciosum, an understory plant, shows special adaptation to shady habitats. Their hybrid is usually found on the areas disturbed by human such as aquacultural ponds. All of them could affect the establishment and growth of other mangrove species, especially in the areas destroyed by human and natural disasters [52, 53], so further study about the ecological significance of the hybrid is important for mangrove management and protection. In particular, it is valuable to conduct well-designed studies on fitness of the hybrid in a variety of habitats, including parental, intermediate, as well as novel habitats.
Conclusions
Acrostichum aureum and A. speciosum are the only two mangrove fern species in the Indo West-Pacific region and they show largely overlapping distribution although their habitats are distinct. In this study, we identified natural hybridization between them for the first time, based on sequence data of three nuclear genes. Interestingly, all the hybrid individuals are likely F1s, suggesting no gene flow between the two species. Therefore, the two species can maintain genetic integrity despite hybridization. Chloroplast DNA shows that the hybridization is unidirectional, with A. speciosum as the maternal parent, and this might be caused by much higher abundance of A. aureum at the study sites. The two species usually occur in differentiated habitats, which might be an effective barrier for interspecific hybridization. Human disturbance on mangrove habitats, however, may bring their spores into contact and thus facilitate the establishment of hybrids of Acrostichum. The impact of hybridization in this genus on mangrove ecosystems should be investigated in future.
Methods
Plant sampling
Plants of A. aureum and A. speciosum and their putative hybrid were sampled from two locations in China: Qinglan Bay, Wenchang, Hainan, and Suixi, Zhanjiang, Guangdong. Frond tissues of A. aureum and A. speciosum and the putative hybrid were collected in plastic bags with silica gels for DNA extraction. In addition, we also collected samples of A. aureum from Sanya, Qionghai, Danzhou and Haikou of Hainan and Huizhou, Guangzhou, and Zhanjiang of Guangdong, where only A. aureum occurs and no putative hybrid individuals were found. Voucher specimens were deposited in the Herbarium of Sun Yat-sen University (SYS). Table 2 listed the details of the samples. We also collected mature fertile fronds of the three taxa in Qinglan Bay, Wenchang, Hainan, to examine the morphology and germination of their spores.
Spore morphology and germination
We collected mature sporangia (partially opened) under a dissecting microscope and transferred them to a drop of water on the microscope slide, tapped gently on the top of the cover glass to scatter the spores. The shape of the spores was examined under a Leica DMI4000 B microscope. Because we found that mature spores released from the sporangia of the three taxa could directly germinate on the fertile fonds after the fertile fonds were placed in petri dishes containing distilled water for three days, we compared the spore germination rates for the three taxa by this means. For each taxon, more than 200 mature spores from multiple fertile fonds were used to calculate the germination rate. Spores were considered be germinated with the emergence of the rhizoid.
Loci studied
Three low-copy nuclear genes, namely, cam, gapCp1 and gapCp2, and one chloroplast intergenic spacer, trnV-trnM, were used in this study. We first used the universal cam primers for angiosperms [54] to test the feasibility in Acrostichum, and when we got the sequences from an individual of A. aureum, we designed specific cam primers camb-F (5′ AAGTTCTTTGGTTGTGATGCAA 3′) and camb-R (5′ GATGGTGAGTGTGTGCATTTG 3′) for Acrostichum. With regard to the gapCp gene, we first used universal gapCp primers in ferns [55] to test their feasibility in Acrostichum, and two bright bands with different length were amplified. We sequenced these two bands and found both of them were members of the gapCp gene family based on sequence homology. We then designated them as gapCp1 and gapCp2 respectively and designed two pairs of specific primers, gapCp1-F (5′ GCTGCATATACATAGGGACATCG 3′) and gapCp1-R (5′ CAAAGTGATCATTCAAAGCAATG 3′), gapCp2-F (5′ CGAGAAGAATCTTTAGCCAACCT 3′) and gapCp2-R (5′ GTCGTACCAAGCCACAAGTTTC 3′), to amplify them. The chloroplast trnV-trnM was amplified using the universal primers trnV and trnM [56].
DNA extraction, PCR and sequencing
Total DNA was extracted from dried frond tissues using the CTAB method according to Doyle and Doyle (1987) [57]. PCR was conducted using KOD-FX DNA polymerase (Toyobo, Osaka, Japan) with the following conditions: initial denaturation at 94°C for 4 min, followed by 30 cycles of 98°C for 10 s, corresponding annealing temperature (54°C for gapCp2 and trnV-trnM, 58°C for gapCp1, and 62°C for cam) for 30 s, and 68°C for 1 min. PCR products were purified using the Pearl Gel Extraction Kit (Pearl Biotech, Guangzhou, China). Purified PCR products were directly sequenced on an ABI 3730 DNA Analyzer with the BigDye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems, Foster City, California, USA). Intra-individual length polymorphism for the nuclear genes could cause failure of direct sequencing from the polymorphic sites. In addition, some individuals, mainly from the putative hybrid, had superimposed chromatograms at multiple sites of the nuclear genes, and the haplotypes could not be inferred. Under these circumstances, cloning sequencing was used to phase the haplotypes. Ligations were conducted using the pMD18-T&A cloning kit (Takara, Dalian, China). Eight positive colonies for each individual were selected for sequencing. The sequences were deposited in GenBank with accession numbers JX575141-JX575175 (Additional file 1: Table S2).
Sequence analyses
All sequences of the three nuclear genes and one chloroplast intergenic spacer were aligned and compared in SeqMan™ (DNASTAR, Madison, Wisconsin, USA). Haplotype inference of these three nuclear genes was implemented with PHASE v2.1 [58, 59]. Haplotype network was constructed for each gene using Network 4.6.1.0 [60] with the median-joining algorithm [61].
References
Arnold ML: Natural Hybridization and Evolution. Oxford: Oxford University Press; 1997.
Rieseberg LH: The role of hybridization in evolution: old wine in new skins. Am J Bot. 1995, 82: 944-953. 10.2307/2445981.
Mallet J: Hybrid speciation. Nature. 2007, 446: 279-283. 10.1038/nature05706.
Dobzhansky T: Genetics and the origin of species. New York: Colombia University Press; 1937.
Rieseberg LH: Hybrid origins of plant species. Annu Rev Ecol Syst. 1997, 28: 359-389. 10.1146/annurev.ecolsys.28.1.359.
Rieseberg LH, Willis JH: Plant speciation. Science. 2007, 317: 910-914. 10.1126/science.1137729.
Soltis PS, Soltis DE: The role of hybridization in plant speciation. Annu Rev Plant Biol. 2009, 60: 561-588. 10.1146/annurev.arplant.043008.092039.
Whitney KD, Randell RA, Rieseberg LH: Adaptive introgression of abiotic tolerance traits in the sunflower Helianthus annuus. New Phytol. 2010, 187: 230-239. 10.1111/j.1469-8137.2010.03234.x.
Knobloch IW: Pteridophyte hybrids. Publ Mus Mich State Univ Biol. 1976, 5: 273-352.
Barrington DS, Haufler CH, Werth CR: Hybridization, reticulation, and species concepts in the ferns. Amer Fern J. 1989, 79 (2): 55-64. 10.2307/1547160.
Walker TG: Hybridization in some species of Pteris L. Evolution. 1958, 12: 82-92. 10.2307/2405906.
Walker TG: Cytology and evolution in the fern genus Pteris L. Evolution. 1962, 16: 27-43. 10.2307/2406264.
Haufler CH: Species and speciation. The biology and evolution of ferns and lycophytes. Edited by: Ranker TA, Haufler CH. Cambridge: Cambridge University Press; 2008: 303-331.
Mayer MS, Mesler MR: Morphometric evidence of hybrid swarms in mixed populations of Polystichum munitum and P. imbricans (Dryopteridaceae). Syst Bot. 1993, 18: 248-260. 10.2307/2419401.
Mullenniex A, Hardig TM, Mesler M: Molecular confirmation of hybrid swarms in the fern genus Polystichum (Dryopteridaceae). Syst Bot. 1998, 23: 421-426. 10.2307/2419374.
Peng CI, Chiang TY: Molecular confirmation of unidirectional hybridization in Begonia × taipeiensis Peng (Begoniaceae) from Taiwan. Ann Missouri Bot Gard. 2000, 87: 273-285. 10.2307/2666164.
Van den Heede CJ, Pajaron S, Pangua E, Viane RLL: A new species and a new hybrid of Asplenium (Aspleniaceae) from Cyprus and evidence of their origin. Belg J Bot. 2002, 135: 92-116.
Xiang L, Werth CR, Emery SN, Mccauley DE: Population-specific gender-biased hybridization between Dryopteris intermedia and D. carthusiana: evidence from chloroplast DNA. Am J Bot. 2000, 87: 1175-1180. 10.2307/2656654.
Duke NC: Mangrove floristics and biogeography. Tropical Mangrove. Edited by: Robertson AI, Alongi DM. 1992, Washington, D.C: Coastal and Estuarine Studies Series, American Geophysical Union, 63-100.
Tomlinson PB: The Botany of Mangroves. Cambridge: Cambridge University Press; 1986.
Adams DC, Tomlinson PB: Acrostichum in Florida. Amer Fern J. 1979, 69: 42-46. 10.2307/1546893.
Taylor FJ: Mangroves in freshwater. Blumea. 1986, 31: 271-272.
Sun WQ, Li XP, Ong BL: Preferential accumulation of D-pinitol in Acrostichum aureum gametophytes in response to salt stress. Physiol Plantarum. 2002, 105: 51-57.
Aksornkoae S, Maxwell GS, Havanond S, Panichsuko S: Plants in mangroves. Bangkok: Chalongrat Co., Ltd Press; 1992.
Medina E, Cuevas E, Popp M, Lugo AE: Soil salinity, sun exposure, and growth of Acrostichum aureum, the mangrove fern. Botanical Gazette. 1990, 151: 41-49. 10.1086/337803.
Turner IM, Xing FW, Corlett RT: An annotated check list of the vascular plants of the south China sea and its shores. Raffles Bull Zool. 1999, 7 (Suppl): 23-116.
Werth CR: Isozyme studies on the Dryopteris ‘spinulosa’ complex, I: The origin of the log fern Dryopteris celsa. Syst Bot. 1991, 16: 446-461. 10.2307/2419336.
Barrington DS: Hybridization and allopolyploidy in Central American Polystichum: Cytological and isozyme documentation. Ann Missouri Bot Gard. 1990, 1990 (77): 297-305.
Gaskin JF, Schaal BA: Hybrid Tamarix widespread in U.S. invasion and undetected in native Asian range. Proc Natl Acad Sci USA. 2002, 99: 11256-11259. 10.1073/pnas.132403299.
Pan J, Zhang D, Sang T: Molecular phylogenetic evidence for the origin of a diploid hybrid of Paeonia (Paeoniaceae). Am J Bot. 2007, 94: 400-408. 10.3732/ajb.94.3.400.
Qiu S, Zhou RC, Li YQ, Havanond S, Jaengjai C, Shi SH: Molecular evidence for natural hybridization between Sonneratia alba and S. griffithii. J Syst Evol. 2008, 46: 391-395.
Chang HM, Chiou WL, Wang JC: Molecular evidence for genetic heterogeneity and the hybrid origin of Acrorumohra subreflexipinna from Taiwan. Amer Fern J. 2009, 99: 61-77. 10.1640/0002-8444-99.2.61.
Guo M, Zhou R, Huang Y, Ouyang J, Shi S: Molecular confirmation of natural hybridization between Lumnitzera racemosa and Lumnitzera littorea. Aquat Bot. 2011, 95: 59-64. 10.1016/j.aquabot.2011.03.001.
Wu W, Zhou R, Huang Y, Boufford DE, Shi S: Molecular evidence for natural intergeneric hybridization between Liquidambar and Altingia. J Plant Res. 2010, 123: 231-239. 10.1007/s10265-009-0275-z.
Dai S, Wu W, Zhang R, Liu T, Chen Y, Shi S, Zhou R: Molecular evidence for hybrid origin of Melastoma intermedium. Biochem Syst Ecol. 2012, 41: 136-141.
Gastony GJ, Yatskievych G: Maternal inheritance of the chloroplast and mitochondrial genomes in cheilanthoid ferns. Am J Bot. 1992, 78: 716-722.
Triest L: Molecular ecology and biogeography of mangrove trees towards conceptual insights on gene flow and barriers, a review. Aquat Bot. 2008, 89: 138-154. 10.1016/j.aquabot.2007.12.013.
Wendel JF, Stewart JM, Rettig JH: Molecular evidence for homoploid reticulate evolution among Australian species of Gossypium. Evolution. 1991, 45: 694-711. 10.2307/2409921.
Tiffin P, Olson MS, Moyle LC: Asymmetric crossing barriers in angiosperms. Proc Roy Soc Lond B. 2001, 268: 861-867. 10.1098/rspb.2000.1578.
Lloyd RM: Reproductive biology and gametophyte morphology of New World populations of Acrostichum aureum. Amer Fern J. 1980, 70: 99-110. 10.2307/1546994.
de Groot GA, During HJ, Maas JW, Schneider H, Vogel JC, Erkens RHJ: Use of rbcL and trnL-F as a two-locus DNA barcode for identification of NW-European ferns: an ecological perspective. PLoS One. 2011, 6: e16371. 10.1371/journal.pone.0016371.
Wagner WH, Chen KL: Abortion of spores and sporangia as a tool in the detection of Dryopteris hybrids. Amer Fern J. 1965, 55: 9-29. 10.2307/1546429.
Ebihara A, Matsumoto S, Ito M: Hybridization Involving Independent Gametophytes in the Vandenboschia radicans complex (Hymenophyllaceae): A New Perspective on the Distribution of Fern Hybrids. Mol Ecol. 2009, 18: 4904-4911. 10.1111/j.1365-294X.2009.04406.x.
Watano Y, Sahashi N: Predominant inbreeding and its genetic consequences in a homosporous fern genus, Sceptridium (Ophioglossaceae). Syst Bot. 1992, 17: 486-502. 10.2307/2419487.
Arnold ML: Natural hybridization as an evolutionary process. Annu Rev Ecol Syst. 1992, 1992 (23): 237-261.
Marcon AB, Barros ICL, Guerra M: A karyotype comparison between two closely related species of Acrostichum. Amer Fern J. 2003, 93: 116-125. 10.1640/0002-8444(2003)093[0116:AKCBTC]2.0.CO;2.
Tindale MD, Roy SK: A cytotaxonomic survey of the Pteridophyta of Australia. Aust Syst Bot. 2002, 15: 839-937. 10.1071/SB00034.
Arnold ML, Hodges SA: Are natural hybrids fit or unfit relative to their parents?. Trends Ecol Evol. 1995, 10: 67-71. 10.1016/S0169-5347(00)88979-X.
Kathiresan K, Bingham BL: Biology of mangroves and mangrove ecosystems. Adv Mar Biol. 2001, 40: 81-251.
Duke NC, Meynecke JO, Dittmann S, Ellison AM, Anger K, Berger U, Cannicci S, Diele K, Ewel KC, Field CD, Koedam N, Lee SY, Marchand C, Nordhaus I, Dahdouh-Guebasl F: A world without mangroves. Science. 2007, 317: 41.
Valiela I, Bowen JL, York JK: Mangrove forests: one of the world’s threatened major tropical environments. Bioscience. 2001, 51: 807-815. 10.1641/0006-3568(2001)051[0807:MFOOTW]2.0.CO;2.
Chan HT: A note on the eradication of Acrostichum aureum ferns in the Matang mangroves, Perak. Peninsular Malaysia. J Trop Forest Sci. 1989, 2 (2): 171-173.
Sharp JM: Responses of the mangrove fern Acrostichum danaeifolium Langsd. & Fisch. (Pteridaceae, Pteridophyta) to disturbances resulting from increased soil salinity and Hurricane Georges at the Jobos Bay National Estuarine Research Reserve, Puerto Rico. Wetl Ecol Manag. 2010, 18: 57-68. 10.1007/s11273-009-9148-4.
Strand AE, Leebens-Mack J, Milligan BG: Nuclear DNA-based markers for plant evolutionary biology. Mol Biol. 1997, 6: 113-118.
Schuettpelz E, Grusz AL, Windham MD, Pryer KM: The utility of nuclear gapCp in resolving polyploid fern origins. Syst Bot. 2008, 33: 621-629. 10.1600/036364408786500127.
Cheng YP, Hwang SY, Lin TP: Potential refugia in Taiwan revealed by the phylogeographical study of Castanopsis carlesii Hayata (Fagaceae). Mol Ecol. 2005, 14: 2075-2085. 10.1111/j.1365-294X.2005.02567.x.
Doyle JJ, Doyle JL: A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull. 1987, 19: 11-15.
Stephens M, Smith N, Donnelly P: A new statistical method for haplotype reconstruction from population data. Am J Hum Genet. 2001, 68: 978-989. 10.1086/319501.
Stephens M, Scheet P: Accounting for Decay of Linkage Disequilibrium in Haplotype Inference and Missing-Data Imputation. Am J Hum Genet. 2005, 76: 449-462. 10.1086/428594.
Network 4.610. http://www.fluxus-engineering.com.
Bandelt HJ, Forster P, Rohl A: Median-joining networks for inferring intraspecific phylogenies. Mol Biol Evol. 1999, 16: 37-48. 10.1093/oxfordjournals.molbev.a026036.
Acknowledgements
We thank Yuechao Chen and Cairong Zhong for their help during the field work. This study is supported by grants from the National Natural Science Foundation of China (31170213, 91231106, 41130208, 31070290 and 41276107), the Fundamental Research Funds for the Central Universities (10lgpy20 and 09lgpy35), and Chang Hungta Science Foundation of Sun Yat-sen University.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare no competing interests.
Authors’ contributions
The experimental design was conceived by RCZ and RSZ and all experiments were performed by RSZ and TL. Data was analyzed by RSZ and RCZ with assistance from WW, YL, LC, LH, YH, and SS. This paper was written by RCZ, RSZ, WW and SS. All authors read and approved the final manuscript.
Electronic supplementary material
12870_2012_1291_MOESM1_ESM.doc
Additional file 1: Table S1: Genotype information at the three nuclear genes for all sampled individuals of the three taxa in Acrostichum. Table S2 Haplotypes at the three nuclear genes and their GenBank accession numbers for the three taxa in Acrostichum.(DOC 390 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Zhang, R., Liu, T., Wu, W. et al. Molecular evidence for natural hybridization in the mangrove fern genus Acrostichum. BMC Plant Biol 13, 74 (2013). https://doi.org/10.1186/1471-2229-13-74
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2229-13-74